
Changes in Soil Microbial Communities under Mixed Organic and Inorganic Nitrogen Addition in Temperate Forests
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design and Sampling
2.3. Soil Property Determinations
2.4. DNA Isolation and Illumina HiSeq Sequencing
2.5. Statistical Analysis
3. Results
3.1. Effects of N Addition on Soil Properties
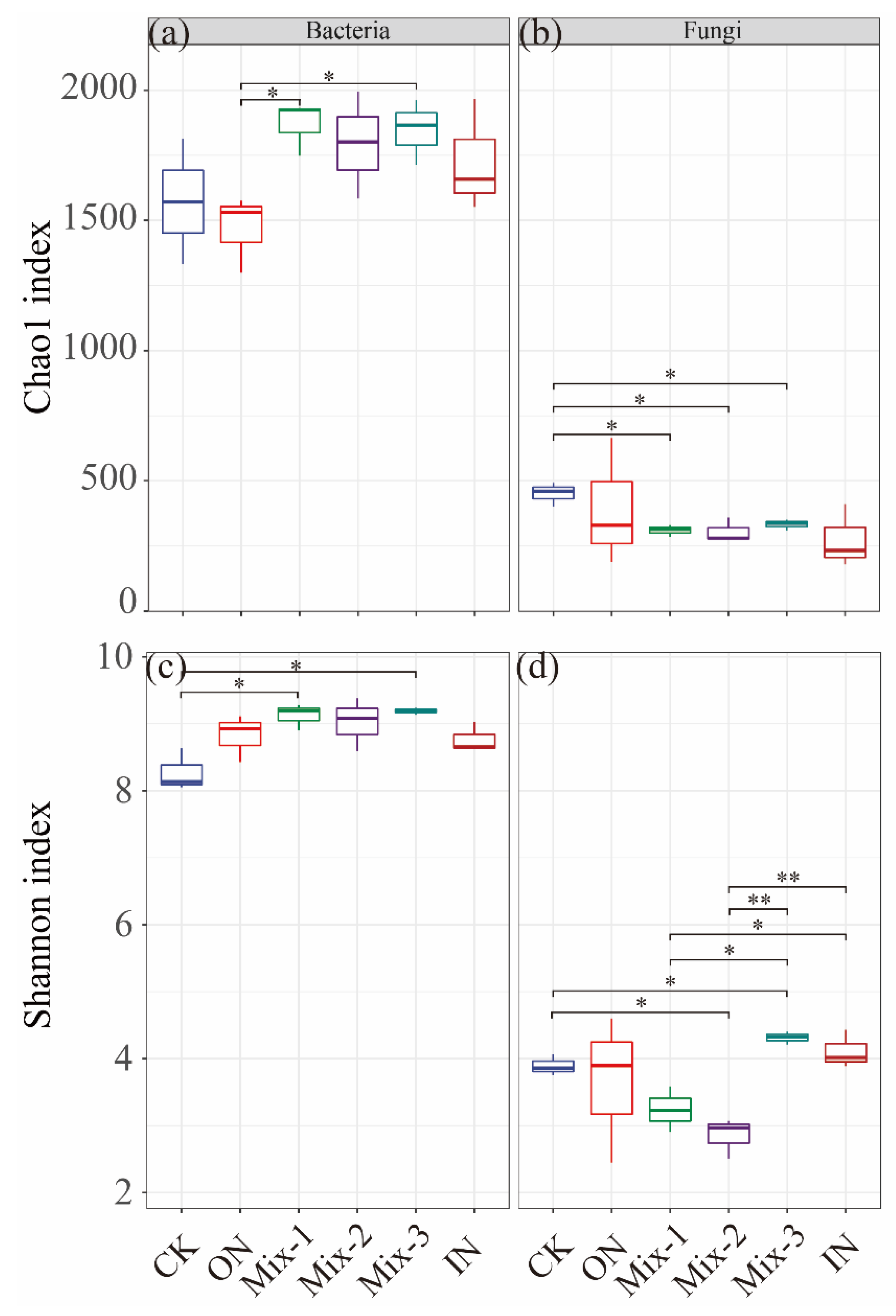
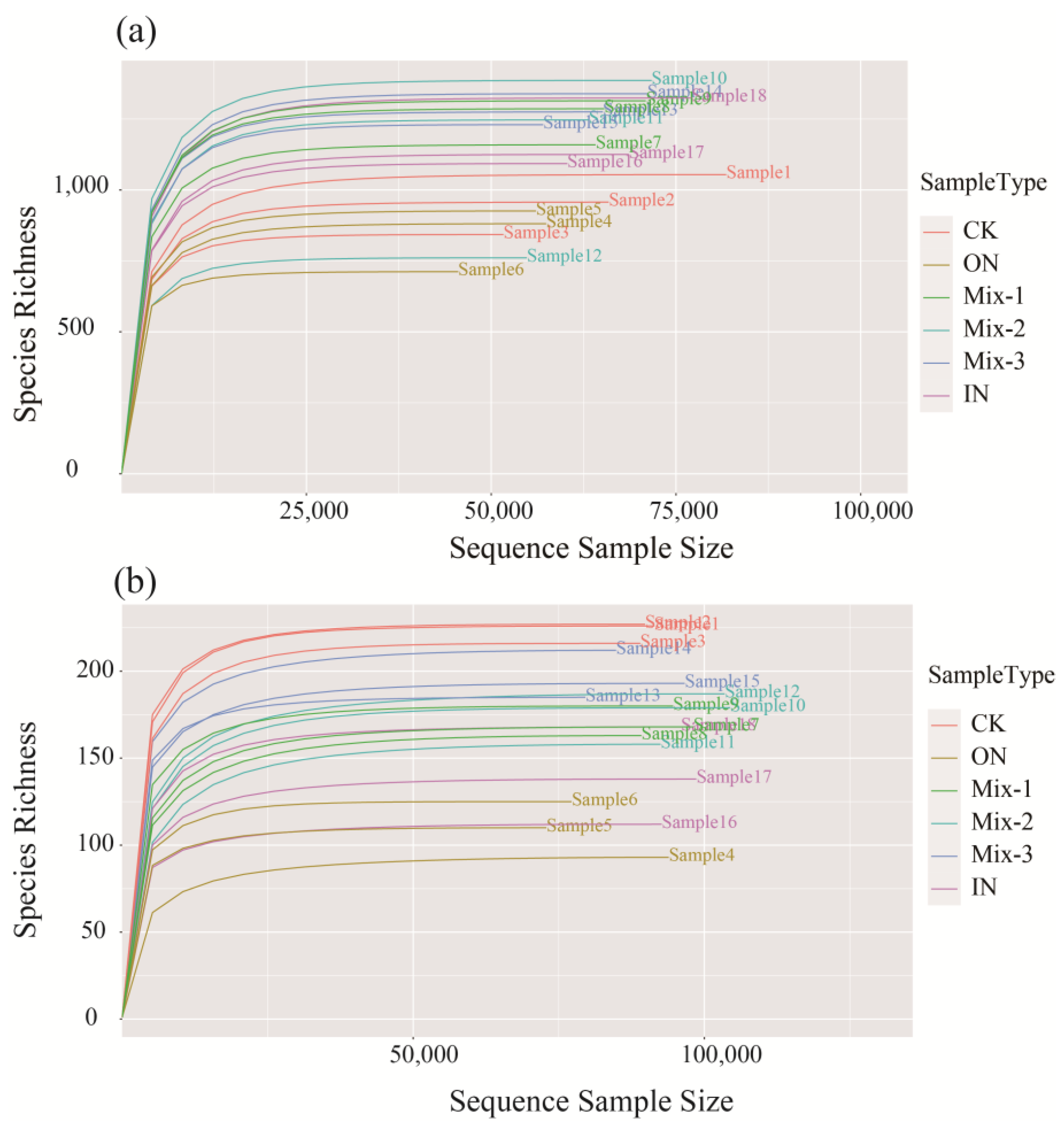
3.2. Soil Bacterial and Fungal Diversity
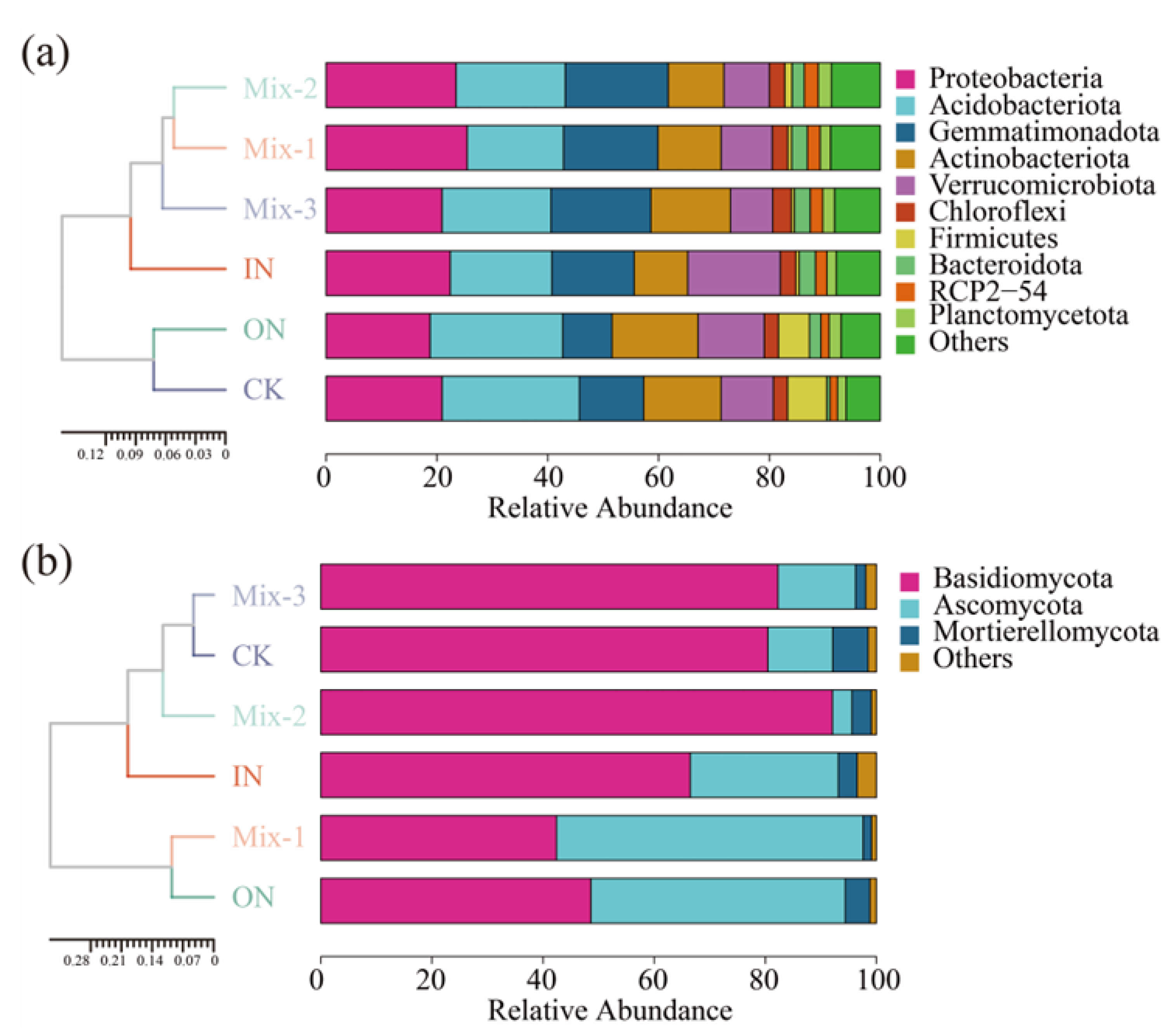
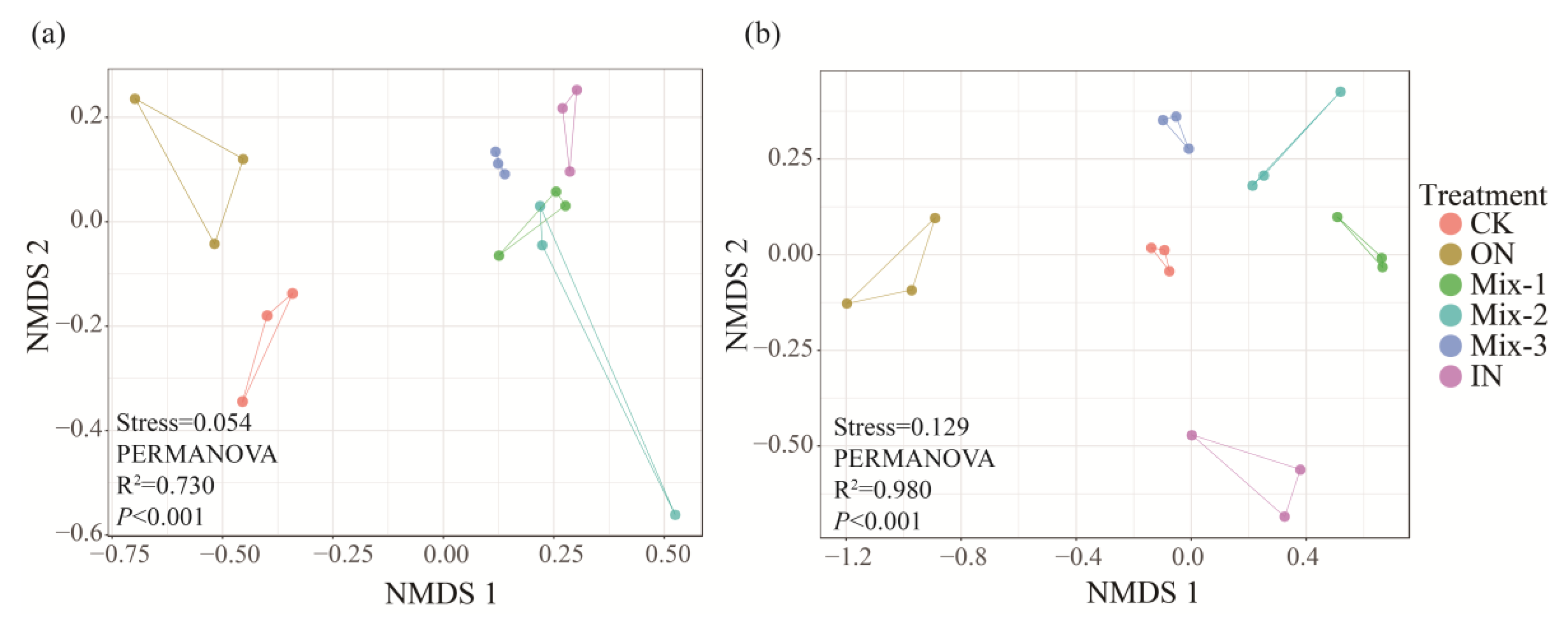
3.3. Soil Microbial Species Composition and Community Structure
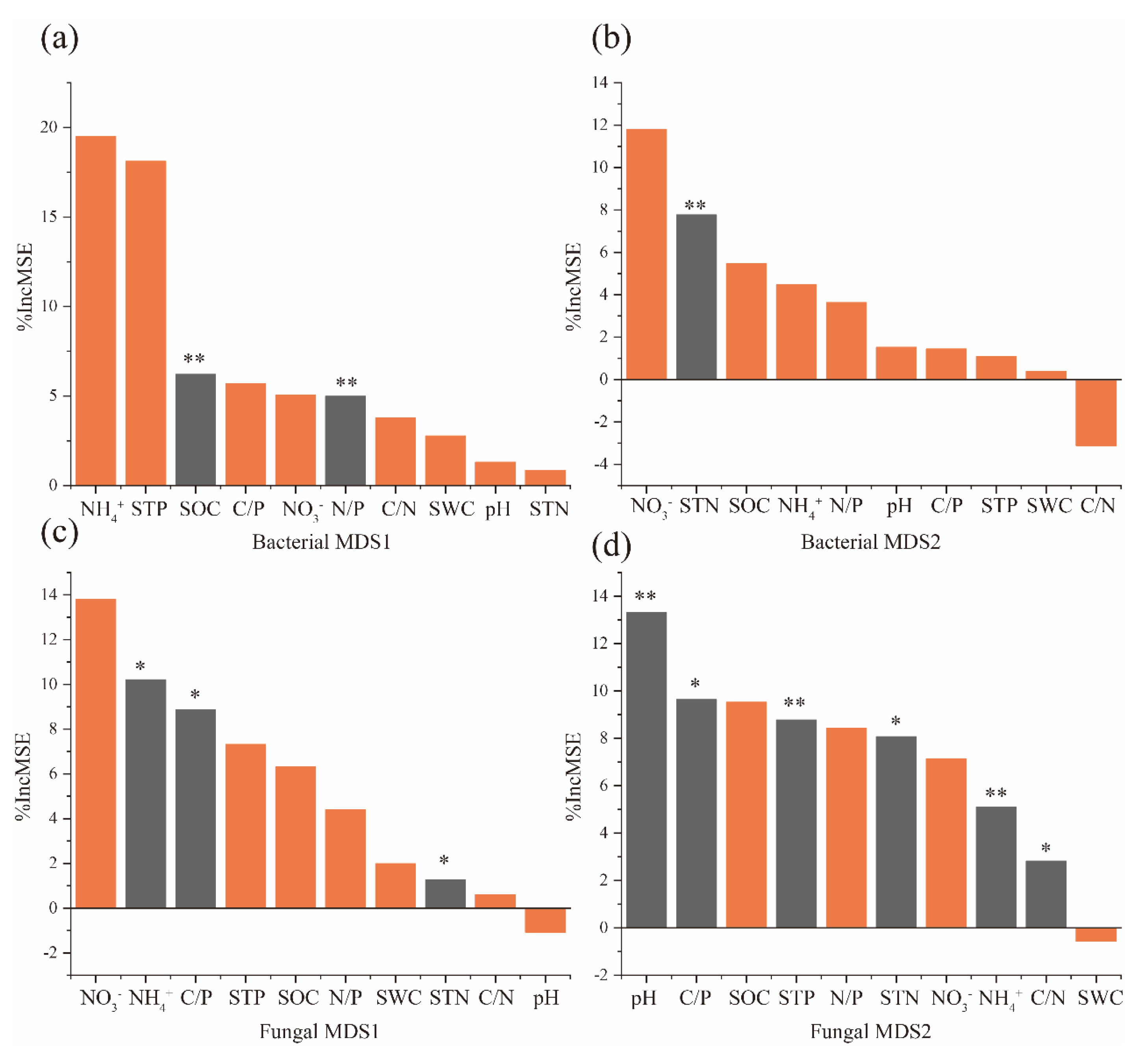
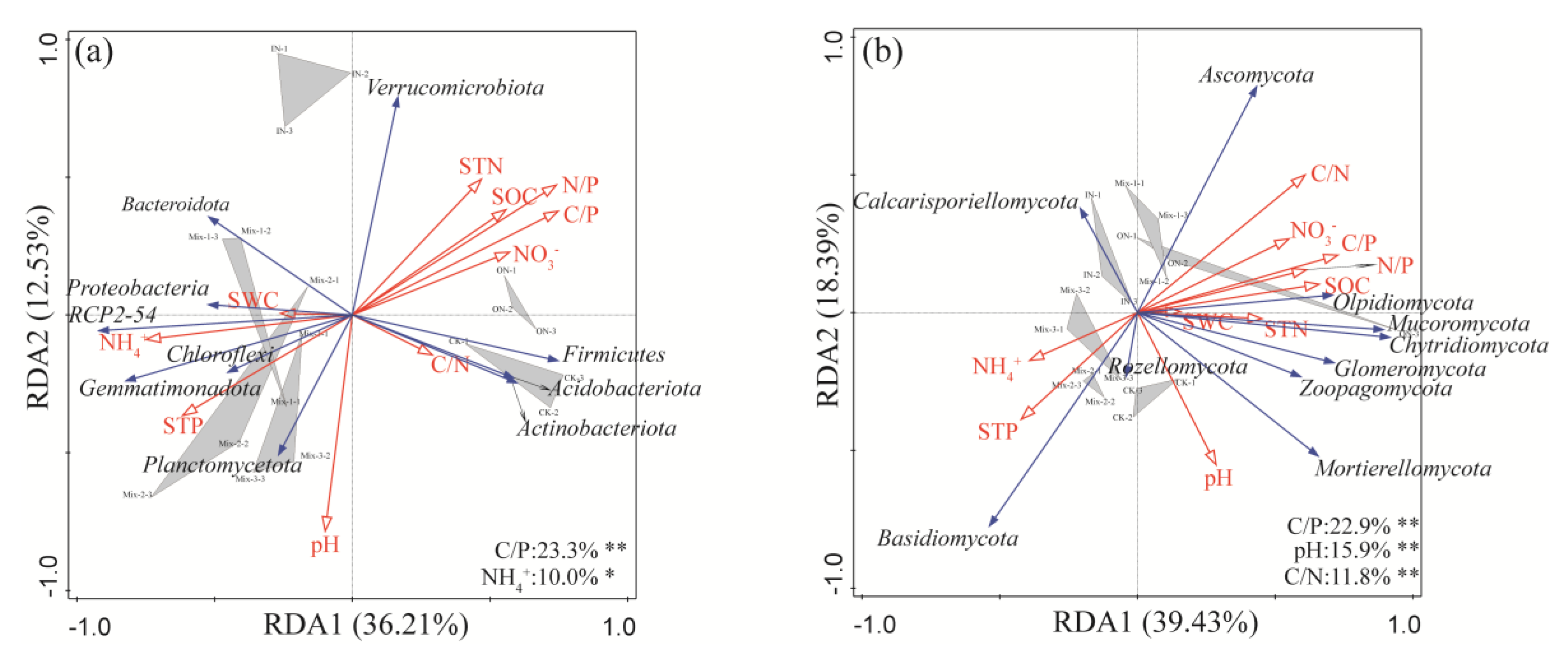
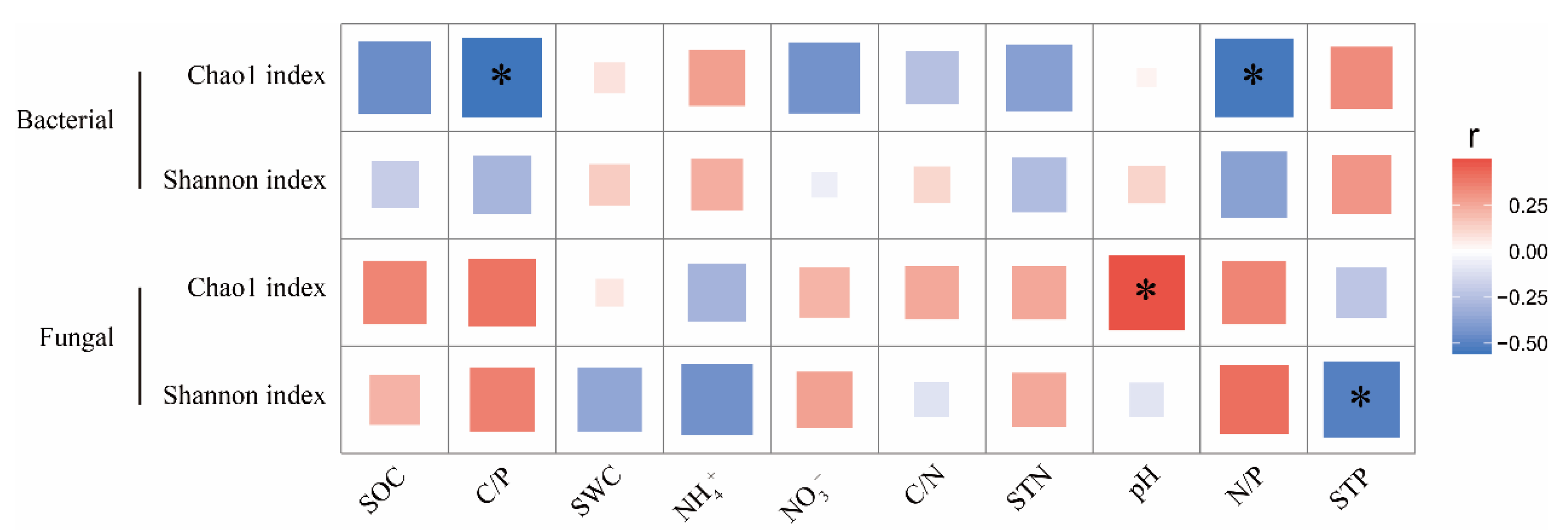
3.4. Relationship between Soil Factors and Microbial Communities
4. Discussion
4.1. Effects of Different Forms of N Addition on Soil Parameters
4.2. Effects of Different Forms of N Addition on the Alpha Diversity of Bacteria and Fungi
4.3. Effects of Different Forms of N Addition on Microbial Community Structure and Composition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Groups | CK | ON | Mix-1 | Mix-2 | Mix-3 | IN | One-Way ANOVA | ||
|---|---|---|---|---|---|---|---|---|---|
| Bacterial | Proteobacteria | 0.209 ab | 0.188 b | 0.255 a | 0.234 ab | 0.209 ab | 0.224 ab | F = 2.094 | p = 0.137 |
| Acidobacteriota | 0.248 a | 0.239 a | 0.174 b | 0.197 ab | 0.197 ab | 0.184 ab | F = 2.020 | p = 0.148 | |
| Gemmatimonadota | 0.116 bc | 0.089 c | 0.17 a | 0.186 a | 0.18 a | 0.149 ab | F = 6.527 | p = 0.004 | |
| Actinobacteriota | 0.14 abc | 0.155 a | 0.114 abc | 0.101 bc | 0.144 ab | 0.097 c | F = 2.986 | p = 0.056 | |
| Verrucomicrobiota | 0.094 b | 0.119 b | 0.092 b | 0.082 b | 0.076 b | 0.167 a | F = 4.98 | p = 0.011 | |
| Fungal | Basidiomycota | 0.805 b | 0.486 d | 0.424 d | 0.921 a | 0.823 b | 0.665 c | F = 46.004 | p = <0.001 |
| Ascomycota | 0.117 de | 0.458 b | 0.552 a | 0.035 e | 0.14 d | 0.267 c | F = 54.469 | p = <0.001 | |


References
- Kanakidou, M.; Myriokefalitakis, S.; Daskalakis, N.; Fanourgakis, G.; Nenes, A.; Baker, A.R.; Tsigaridis, K.; Mihalopoulos, N. Past, Present and Future Atmospheric Nitrogen Deposition. J. Atmos. Sci. 2016, 73, 2039–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, R.E.; Craine, J.M.; Lany, N.K.; Jonard, M.; Ollinger, S.V.; Groffman, P.M.; Fulweiler, R.W.; Angerer, J.; Read, Q.D.; Reich, P.B.; et al. Evidence, causes, and consequences of declining nitrogen availability in terrestrial ecosystems. Science 2022, 376, eabh3767. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Jiang, Y.; Fang, J.Y.; de Vries, W. Inorganic nitrogen deposition in China’s forests: Status and characteristics. Atmos. Env. 2014, 98, 474–482. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Duan, L.; Mo, J.; Du, E.; Shen, J.; Lu, X.; Zhang, Y.; Zhou, X.; He, C.; Zhang, F. Nitrogen deposition and its ecological impact in China: An overview. Env. Pollut. 2011, 159, 2251–2264. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.J.; Dise, N.B.; Field, C.D.; Dore, A.J.; Caporn, S.J.M.; Stevens, C.J. Nitrogen deposition and plant biodiversity: Past, present, and future. Front. Ecol. Env. 2017, 15, 431–436. [Google Scholar] [CrossRef]
- Gilliam, F.S. Responses of Forest Ecosystems to Nitrogen Deposition. Forests 2021, 12, 1190. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.H.; Wang, C.K.; Zheng, M.H.; Jiang, L.F.; Luo, Y.Q. Patterns and mechanisms of responses by soil microbial communities to nitrogen addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L. Microbial community response to nitrogen deposition in northern forest ecosystems. Soil Biol. Biochem. 2004, 36, 1443–1451. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, H.Y.H.; Ruan, H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tan, X.; Fu, S.; Shen, W. Canopy and Understory Nitrogen Addition Alters Organic Soil Bacterial Communities but Not Fungal Communities in a Temperate Forest. Front. Microbiol. 2022, 13, 888121. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.A.M.; Anthony, M.A.; Pec, G.J.; Trocha, L.K.; Trzebny, A.; Geyer, K.M.; van Diepen, L.T.A.; Frey, S.D. Fungal community structure and function shifts with atmospheric nitrogen deposition. Glob. Chang. Biol. 2021, 27, 1349–1364. [Google Scholar] [CrossRef] [PubMed]
- Zechmeister-Boltenstern, S.; Michel, K.; Pfeffer, M. Soil microbial community structure in European forests in relation to forest type and atmospheric nitrogen deposition. Plant Soil 2010, 343, 37–50. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Mo, J. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, K.; Sun, T.; Tian, J.H.; Chen, Z.H.; Chen, L.J. Soil microbial biomass, phosphatase and their relationships with phosphorus turnover under mixed inorganic and organic nitrogen addition in a Larix gmelinii plantation. For. Ecol. Manag. 2018, 422, 313–322. [Google Scholar] [CrossRef]
- Yan, Y.; Sun, X.; Sun, F.; Zhao, Y.; Sun, W.; Guo, J.; Zhang, T. Sensitivity of soil fungal and bacterial community compositions to nitrogen and phosphorus additions in a temperate meadow. Plant Soil 2021, 471, 477–490. [Google Scholar] [CrossRef]
- Allison, S.D.; Czimczik, C.I.; Treseder, K.K. Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest. Glob. Chang. Biol. 2008, 14, 1156–1168. [Google Scholar] [CrossRef] [Green Version]
- Lilleskov, E.A.; Kuyper, T.W.; Bidartondo, M.I.; Hobbie, E.A. Atmospheric nitrogen deposition impacts on the structure and function of forest mycorrhizal communities: A review. Env. Pollut. 2019, 246, 148–162. [Google Scholar] [CrossRef]
- Li, J.; Sang, C.P.; Yang, J.Y.; Qu, L.R.; Xia, Z.W.; Sun, H.; Jiang, P.; Wang, X.G.; He, H.B.; Wang, C. Stoichiometric imbalance and microbial community regulate microbial elements use efficiencies under nitrogen addition. Soil Biol. Biochem. 2021, 156, 108207. [Google Scholar] [CrossRef]
- Tian, D.S.; Niu, S.L. A global analysis of soil acidification caused by nitrogen addition. Env. Res. Lett. 2015, 10, 024019. [Google Scholar] [CrossRef]
- Zhang, X.M.; Liu, W.; Zhang, G.M.; Jiang, L.; Han, X.G. Mechanisms of soil acidification reducing bacterial diversity. Soil Biol. Biochem. 2015, 81, 275–281. [Google Scholar] [CrossRef]
- Chen, D.M.; Lan, Z.C.; Bai, X.; Grace, J.B.; Bai, Y.F. Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe. J. Ecol. 2013, 101, 1322–1334. [Google Scholar] [CrossRef]
- Cline, L.C.; Huggins, J.A.; Hobbie, S.E.; Kennedy, P.G. Organic nitrogen addition suppresses fungal richness and alters community composition in temperate forest soils. Soil Biol. Biochem. 2018, 125, 222–230. [Google Scholar] [CrossRef]
- Lim, H.; Jamtgard, S.; Oren, R.; Gruffman, L.; Kunz, S.; Nasholm, T. Organic nitrogen enhances nitrogen nutrition and early growth of Pinus sylvestris seedlings. Tree Physiol. 2022, 42, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wei, K.; Sun, T.; Jiang, N.; Chen, Z.; Feng, J.; Cai, K.; Chen, L. Different forms of nitrogen deposition show variable effects on soil organic nitrogen turnover in a temperate forest. Appl. Soil Ecol. 2022, 169, 104212. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.Y.; Zou, H.T.; Kou, L.; Yang, Y.; Wen, X.F.; Li, S.G.; Wang, H.M.; Sun, X.M. Contrasting effects of ammonium and nitrate additions on the biomass of soil microbial communities and enzyme activities in subtropical China. Biogeosciences 2017, 14, 4815–4827. [Google Scholar] [CrossRef] [Green Version]
- Qu, W.D.; Han, G.X.; Eller, F.; Xie, B.H.; Wang, J.; Wu, H.T.; Li, J.Y.; Zhao, M.L. Nitrogen input in different chemical forms and levels stimulates soil organic carbon decomposition in a coastal wetland. Catena 2020, 194, 104672. [Google Scholar] [CrossRef]
- Koulympoudi, L.; Papafilippou, A.; Tzanoudaki, M.; Chatzissavvidis, C.; Salamalikis, V. Effect of nitrogen form on trifoliate orange (Poncirus trifoliata (L.) Raf.) and sour orange (Citrus aurantium L.) plants grown under saline conditions. J. Plant Nutr. 2021, 44, 2546–2558. [Google Scholar] [CrossRef]
- Cornell, S.E. Atmospheric nitrogen deposition: Revisiting the question of the importance of the organic component. Env. Pollut. 2011, 159, 2214–2222. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, L.; Liu, X.J.; Li, W.Q.; Lü, S.H.; Zheng, L.X.; Bai, Z.C.; Cai, G.Y.; Zhang, F.S. Atmospheric organic nitrogen deposition in China. Atmos. Env. 2012, 46, 195–204. [Google Scholar] [CrossRef]
- Moran-Zuloaga, D.; Dippold, M.; Glaser, B.; Kuzyakov, Y. Organic nitrogen uptake by plants: Reevaluation by position-specific labeling of amino acids. Biogeochemistry 2015, 125, 359–374. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Joergensen, R.G.; Ludwig, B. Pathways of nitrogen utilization by soil microorganisms—A review. Soil Biol. Biochem. 2010, 42, 2058–2067. [Google Scholar] [CrossRef]
- Guo, P.; Wang, C.Y.; Jia, Y.; Wang, Q.A.; Han, G.M.; Tian, X.J. Responses of soil microbial biomass and enzymatic activities to fertilizations of mixed inorganic and organic nitrogen at a subtropical forest in East China. Plant Soil 2011, 338, 355–366. [Google Scholar] [CrossRef]
- Dong, L.L.; Berg, B.; Sun, T.; Wang, Z.W.; Han, X.G. Response of fine root decomposition to different forms of N deposition in a temperate grassland. Soil Biol. Biochem. 2020, 147, 107845. [Google Scholar] [CrossRef]
- Du, Y.; Guo, P.; Liu, J.; Wang, C.; Yang, N.; Jiao, Z. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Glob. Chang. Biol. 2014, 20, 3222–3228. [Google Scholar] [CrossRef]
- Zhu, H.; Gong, L.; Luo, Y.; Tang, J.; Ding, Z.; Li, X. Effects of Litter and Root Manipulations on Soil Bacterial and Fungal Community Structure and Function in a Schrenk’s Spruce (Picea schrenkiana) Forest. Front. Plant Sci. 2022, 13, 849483. [Google Scholar] [CrossRef]
- Zhao, J.J.; Gong, L. Response of Fine Root Carbohydrate Content to Soil Nitrogen Addition and Its Relationship with Soil Factors in a Schrenk (Picea schrenkiana) Forest. J. Plant Growth Regul. 2021, 40, 1210–1221. [Google Scholar] [CrossRef]
- Group, C. China Soil System Classification (Amendment Scheme); Agricultural Science and Technology Press of China: Beijing, China, 1995. [Google Scholar]
- Bao, S. Soil Agrochemical Analysis; China Agricultural Press: Beijing, China, 2000; p. 30. (In Chinese) [Google Scholar]
- Zhu, H.; Gong, L.; Ding, Z.; Li, Y. Effects of litter and root manipulations on soil carbon and nitrogen in a Schrenk’s spruce (Picea schrenkiana) forest. PLoS ONE 2021, 16, e0247725. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 20 September 2022).
- Yang, X.D.; Anwar, E.; Zhou, J.; He, D.; Gao, Y.C.; Lv, G.H.; Cao, Y.E. Higher association and integration among functional traits in small tree than shrub in resisting drought stress in an arid desert. Env. Exp. Bot. 2022, 201, 104993. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, Y.; Tang, Z.; Feng, Y.; Zhu, X.; Xu, W.; Bai, Y.; Zhou, G.; Xie, Z.; Fang, J. Patterns of nitrogen and phosphorus pools in terrestrial ecosystems in China. Earth Syst. Sci. Data 2021, 13, 5337–5351. [Google Scholar] [CrossRef]
- Wang, C.Y.; Zhou, J.W.; Liu, J.; Jiang, K.; Du, D.L. Responses of soil N-fixing bacteria communities to Amaranthus retroflexus invasion under different forms of N deposition. Agr. Ecosyst. Env. 2017, 247, 329–336. [Google Scholar] [CrossRef]
- Barak, P.; Jobe, B.O.; Krueger, A.R.; Peterson, L.A.; Laird, D.A. Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant Soil 1997, 197, 61–69. [Google Scholar] [CrossRef]
- Martikainen, P.J.; Aarnio, T.; Taavitsainen, V.-M.; Päivinen, L.; Salonen, K. Mineralization of carbon and nitrogen in soil samples taken from three fertilized pine stands: Long-term effects. Plant Soil 1989, 114, 99–106. [Google Scholar] [CrossRef]
- Bai, T.S.; Wang, P.; Ye, C.L.; Hu, S.J. Form of nitrogen input dominates N effects on root growth and soil aggregation: A meta-analysis. Soil Biol. Biochem. 2021, 157, 108251. [Google Scholar] [CrossRef]
- Song, L.; Kuang, F.; Skiba, U.; Zhu, B.; Liu, X.; Levy, P.; Dore, A.; Fowler, D. Bulk deposition of organic and inorganic nitrogen in southwest China from 2008 to 2013. Env. Pollut. 2017, 227, 157–166. [Google Scholar] [CrossRef]
- Dong, L.L.; Sun, T.; Berg, B.; Zhang, L.L.; Zhang, Q.Q.; Wang, Z.W. Effects of different forms of N deposition on leaf litter decomposition and extracellular enzyme activities in a temperate grassland. Soil Biol. Biochem. 2019, 134, 78–80. [Google Scholar] [CrossRef]
- Du, E.Z.; Terrer, C.; Pellegrini, A.F.A.; Ahlstrom, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.H.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221. [Google Scholar] [CrossRef]
- Liu, G.C.; Yan, G.Y.; Huang, B.B.; Sun, X.Y.; Xing, Y.J.; Wang, Q.G. Long-term nitrogen addition alters nutrient foraging strategies of Populus davidiana and Betula platyphylla in a temperate natural secondary forest. Eur. J. For. Res. 2022, 141, 307–320. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Liu, W.; Long, X.E.; Jia, Y.; Wang, X.; Chen, Y. Different Responses of Soil Bacterial Communities to Nitrogen Addition in Moss Crust. Front. Microbiol. 2021, 12, 665975. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Wang, M.; Zhang, W.; Ni, Z.; Hashidoko, Y.; Shen, W. Ammonium nitrogen content is a dominant predictor of bacterial community composition in an acidic forest soil with exogenous nitrogen enrichment. Sci. Total Env. 2018, 624, 407–415. [Google Scholar] [CrossRef]
- Wang, J.; Shi, X.; Zheng, C.; Suter, H.; Huang, Z. Different responses of soil bacterial and fungal communities to nitrogen deposition in a subtropical forest. Sci. Total Env. 2021, 755, 142449. [Google Scholar] [CrossRef]
- Wang, Z.H.; Wang, Z.R.; Li, T.P.; Wang, C.; Dang, N.; Wang, R.Z.; Jiang, Y.; Wang, H.Y.; Li, H. N and P fertilization enhanced carbon decomposition function by shifting microbes towards an r-selected community in meadow grassland soils. Ecol. Indic. 2021, 132, 108306. [Google Scholar] [CrossRef]
- Turlapati, S.A.; Minocha, R.; Bhiravarasa, P.S.; Tisa, L.S.; Thomas, W.K.; Minocha, S.C. Chronic N-amended soils exhibit an altered bacterial community structure in Harvard Forest, MA, USA. FEMS Microbiol. Ecol. 2013, 83, 478–493. [Google Scholar] [CrossRef] [Green Version]
- Schwede, D.B.; Simpson, D.; Tan, J.; Fu, J.S.; Dentener, F.; Du, E.; deVries, W. Spatial variation of modelled total, dry and wet nitrogen deposition to forests at global scale. Env. Pollut. 2018, 243, 1287–1301. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, H.; Gao, H.; An, S.S. Response and driving factors of soil microbial diversity related to global nitrogen addition. Land Degrad. Dev. 2020, 31, 190–204. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Khachane, A.N.; Campbell, C.D.; Thomas, N.; Freitag, T.E.; Abu Al-Soud, W.; Sorensen, S.; Bardgett, R.D.; Singh, B.K. It is elemental: Soil nutrient stoichiometry drives bacterial diversity. Env. Microbiol. 2017, 19, 1176–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.F.; Fu, B.J.; Zheng, X.X.; Liu, G.H. Plant biomass, soil water content and soil N:P ratio regulating soil microbial functional diversity in a temperate steppe: A regional scale study. Soil Biol. Biochem. 2010, 42, 445–450. [Google Scholar] [CrossRef]
- Lammel, D.R.; Barth, G.; Ovaskainen, O.; Cruz, L.M.; Zanatta, J.A.; Ryo, M.; de Souza, E.M.; Pedrosa, F.O. Direct and indirect effects of a pH gradient bring insights into the mechanisms driving prokaryotic community structures. Microbiome 2018, 6, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousk, J.; Brookes, P.C.; Baath, E. Investigating the mechanisms for the opposing pH relationships of fungal and bacterial growth in soil. Soil Biol. Biochem. 2010, 42, 926–934. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberan, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Su, W.; Chen, H.; Barberan, A.; Zhao, H.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe. Glob. Chang. Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef]
- Krumins, J.A.; Dighton, J.; Gray, D.; Franklin, R.B.; Morin, P.J.; Roberts, M.S. Soil microbial community response to nitrogen enrichment in two scrub oak forests. For. Ecol. Manag. 2009, 258, 1383–1390. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Zhu, R.; Wang, X.; Xu, X.; Ai, C.; He, P.; Liang, G.; Zhou, W.; Zhu, P. Effect of high soil C/N ratio and nitrogen limitation caused by the long-term combined organic-inorganic fertilization on the soil microbial community structure and its dominated SOC decomposition. J. Environ. Manag. 2022, 303, 114155. [Google Scholar] [CrossRef]
- Dimitriu, P.A.; Grayston, S.J. Relationship between soil properties and patterns of bacterial beta-diversity across reclaimed and natural boreal forest soils. Microb. Ecol. 2010, 59, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.C.; Xiong, J.B.; Zhang, H.Y.; Feng, Y.Z.; Lin, X.G.; Li, X.Y.; Liang, W.J.; Chu, H.Y. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.W.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470; discussion 3503–3414. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Lu, X.K.; Mori, T.; Mao, Q.G.; Zhou, K.J.; Zhou, G.Y.; Nie, Y.X.; Mo, J.M. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, X.; Zhou, B.K.; Zhao, B.S.; Ma, M.C.; Guan, D.W.; Li, J.; Chen, S.F.; Cao, F.M.; Shen, D.L.; et al. Thirty four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast China. Soil Biol. Biochem. 2016, 95, 135–143. [Google Scholar] [CrossRef]





| Treatment | CK | ON | Mix-1 | Mix-2 | Mix-3 | IN | One-Way ANOVA | |
|---|---|---|---|---|---|---|---|---|
| F | p | |||||||
| SOC (g kg−1) | 126.80 (2.2)c | 194.90 (23.96)a | 101.98 (3.48)cd | 117.16 (3.37)cd | 91.04 (4.68)d | 158.59 (16.81)b | 19.82 | <0.001 |
| STN (g kg−1) | 9.20 (0.24)b | 11.86 (1.04)a | 6.64 (0.66)cd | 8.51 (0.41)bc | 6.49 (0.42)d | 11.86 (1.62)a | 15.01 | <0.001 |
| STP (g kg−1) | 0.98 (0.03)d | 0.96 (0.02)d | 1.18 (0.1)bc | 2.26 (0.07)a | 1.32 (0.09)b | 1.14 (0.08)c | 92.16 | <0.001 |
| C/N | 13.79 (0.31)b | 16.39 (0.67)a | 15.48 (1.19)a | 13.78 (0.35)b | 14.04 (0.48)b | 13.44 (0.47)b | 6.45 | 0.004 |
| C/P | 129.62 (5.49)b | 204.03 (29.31)a | 86.91 (8.49)c | 51.76 (0.84)d | 69.23 (6.91)cd | 138.54 (5.46)b | 35.91 | <0.001 |
| N/P | 9.41 (0.54)b | 12.40 (1.31)a | 5.69 (0.94)c | 3.76 (0.15)d | 4.93 (0.45)cd | 10.34 (0.75)b | 38.40 | <0.001 |
| pH | 7.81 (0.04)a | 7.29 (0.06)a | 7.35 (0.16)a | 7.51 (0.21)a | 7.60 (0.06)a | 6.95 (0.15)b | 4.05 | 0.022 |
| NO3− (mg kg−1) | 8.55 (0.85)c | 55.39 (9.43)a | 2.70 (0.96)c | 1.53 (0.77)c | 11.09 (2.21)bc | 20.82 (4.93)b | 40.65 | <0.001 |
| NH4+ (mg kg−1) | 9.88 (0.85)d | 10.50 (2.95)d | 29.18 (4.42)bc | 67.44 (15.23)a | 20.55 (3.22)cd | 35.40 (1.64)b | 20.13 | <0.001 |
| SWC (%) | 43.55 (1.9)bc | 53.75 (7.21)ab | 55.58 (12.17)ab | 63.32 (8.39)a | 35.42 (1.32)c | 49.12 (8.84)abc | 3.24 | 0.440 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Z.; Gong, L.; Zhu, H.; Tang, J.; Li, X.; Zhang, H. Changes in Soil Microbial Communities under Mixed Organic and Inorganic Nitrogen Addition in Temperate Forests. Forests 2023, 14, 21. https://0-doi-org.brum.beds.ac.uk/10.3390/f14010021
Ding Z, Gong L, Zhu H, Tang J, Li X, Zhang H. Changes in Soil Microbial Communities under Mixed Organic and Inorganic Nitrogen Addition in Temperate Forests. Forests. 2023; 14(1):21. https://0-doi-org.brum.beds.ac.uk/10.3390/f14010021
Chicago/Turabian StyleDing, Zhaolong, Lu Gong, Haiqiang Zhu, Junhu Tang, Xiaochen Li, and Han Zhang. 2023. "Changes in Soil Microbial Communities under Mixed Organic and Inorganic Nitrogen Addition in Temperate Forests" Forests 14, no. 1: 21. https://0-doi-org.brum.beds.ac.uk/10.3390/f14010021