
Color and Chemical Composition of Timber Woods (Daniellia oliveri, Isoberlinia doka, Khaya senegalensis, and Pterocarpus erinaceus) from Different Locations in Southern Mali

Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of the Wood Materials and Wood Species
2.2. Sample Preparation
2.3. Analytical Techniques
2.4. Data Analysis
3. Results and Discussion
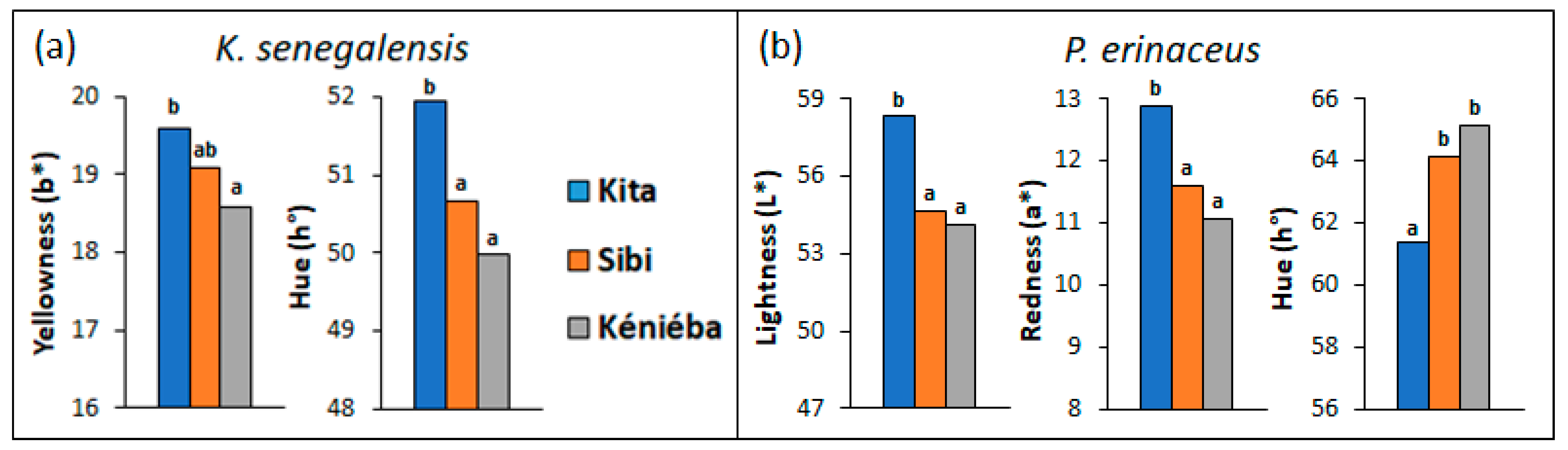
3.1. Variations in Wood Color Parameters
3.2. Variations in Wood FTIR Absorption Bands
3.3. Variations in Wood FTIR Ratios
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jozsa, L.A.; Middleton, G.R. A Discussion of Wood Quality Attributes and Their Practical Implications; Forintek Canada Corporation Special Publication No. SP-34; Forintek Canada Corporation: Vancouver, BC, Canada, 1944. [Google Scholar]
- Rowell, R.M.; Pettersen, R.; Han, J.S.; Rowell, J.S.; Tshabalala, M.A. Cell Wall Chemistry. In Handbook of Wood Chemistry and Wood Composites; Rowell, R.M., Ed.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Wegst, U.G. Wood for sound. Am. J. Bot. 2006, 93, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Traoré, B.; Brancheriau, L.; Perré, P.; Stevanovic, T.; Diouf, P. Acoustic quality of vène wood (Pterocarpus erinaceus Poir.) for xylophone instrument manufacture in Mali. Ann. For. Sci. 2010, 67, 815. [Google Scholar] [CrossRef] [Green Version]
- Fengel, D.; Wegener, G. Wood: Chemistry, Ultrastructure, Reactions; Walter de Gruyter: Berlin, Germany, 1984. [Google Scholar]
- Nabais, C.; Hansen, J.K.; David-Schwartz, R.; Klisz, M.; Lopez, R.; Rozenberg, P. The effect of climate on wood density: What provenance trials tell us? For. Ecol. Manag. 2018, 408, 148–156. [Google Scholar] [CrossRef]
- Hevia, A.; Campelo, F.; Chambel, R.; Vieira, J.; Alía, R.; Majada, J.; Sánchez-Salguero, R. Which matters more for wood traits in Pinus halepensis Mill., provenance or climate? Ann. For. Sci. 2020, 77, 55. [Google Scholar] [CrossRef]
- Moya, R.; Gaitan-Alvarez, J.; Berrocal, A.; Caceres, C.B.; Hernandez, R.E. Wood properties of nine acetylated tropical hardwoods from fast-growth plantations in Costa Rica. Wood Fiber Sci. 2022, 54, 134–148. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Academic Press: Cambridge, MA, USA, 1976. [Google Scholar]
- Côté, B.; Hendershot, W.H.; Fyles, J.W.; Roy, A.G.; Bradley, R.; Biron, P.M.; Courchesne, F. The phenology of fine root growth in a maple-dominated ecosystem: Relationships with some soil properties. Plant Soil 1998, 201, 59–69. [Google Scholar] [CrossRef]
- Fritts, H.C.; Swetnam, T.W. Dendroecology: A tool for evaluating variations in past and present forest environments. Adv. Ecol. Res. 1989, 19, 111–188. [Google Scholar]
- Schweingruber, F.H. Trees and Wood in Dendrochronology: Morphological, Anatomical, and Tree-Ring Analytical Characteristics of Trees Frequently used in Dendrochronology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Hughes, M.K. Dendrochronology in climatology—The state of the art. Dendrochronologia 2002, 20, 95–116. [Google Scholar] [CrossRef]
- Sass-Klaassen, U. Tree physiology: Tracking tree carbon gain. Nat. Plants 2015, 1, 15175. [Google Scholar] [CrossRef]
- Ibanez, T.; Chave, J.; Barrabé, L.; Elodie, B.; Boutreux, T.; Trueba, S.; Vandrot, H.; Birnbaum, P. Community variation in wood density along a bioclimatic gradient on a hyper-diverse tropical island. J. Veg. Sci. 2017, 28, 19–33. [Google Scholar] [CrossRef]
- Jansen, K.; Sohrt, J.; Kohnle, U.; Ensminger, I.; Gessler, A. Tree ring isotopic composition, radial increment and height growth reveal provenance-specific reactions of Douglas-fir towards environmental parameters. Trees 2013, 27, 37–52. [Google Scholar] [CrossRef]
- Kusiak, W.; Majka, J.; Ratajczak, I.; Górska, M.; Zborowska, M. Evaluation of environmental impact on selected properties of lime (Tilia cordata Mill.) wood. Forests 2020, 11, 746. [Google Scholar] [CrossRef]
- Sotelo Montes, C.; Hernández, R.E.; Beaulieu, J.; Weber, J.C. Genetic variation in wood color and its correlations with tree growth and wood density of Calycophyllum spruceanum at an early age in the Peruvian Amazon. New For. 2008, 35, 57–73. [Google Scholar] [CrossRef]
- Moya, R.; Calvo-Alvarado, J. Variation of wood color parameters of Tectona grandis and its relationship with physical environmental factors. Ann. For. Sci. 2012, 69, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Bessa, F.; Sousa, V.; Quilhó, T.; Pereira, H. An Integrated Similarity Analysis of Anatomical and Physical Wood Properties of Tropical Species from India, Mozambique, and East Timor. Forests 2022, 13, 1675. [Google Scholar] [CrossRef]
- Rana, R.; Müller, G.; Naumann, A.; Polle, A. FTIR spectroscopy in combination with principal component analysis or cluster analysis as a tool to distinguish beech (Fagus sylvatica L.) trees grown at different sites. Holzforschung 2008, 62, 530–538. [Google Scholar] [CrossRef]
- Santoni, I.; Callone, E.; Sandak, A.; Sandak, J.; Dirè, D. Solid state NMR and IR characterization of wood polymer structure in relation to tree provenance. Carbohydr. Polym. 2015, 17, 710–721. [Google Scholar] [CrossRef]
- Sharma, V.; Yadav, J.; Kumar, R.; Tesarova, D.; Ekielski, A.; Mishra, P.K. On the rapid and non-destructive approach for wood identification using ATR-FTIR spectroscopy and chemometric methods. Vib. Spectrosc. 2020, 110, 103097. [Google Scholar] [CrossRef]
- Toscano, G.; Maceratesi, V.; Leoni, E.; Stipa, P.; Laudadio, E.; Sabbatini, S. FTIR spectroscopy for determination of the raw materials used in wood pellet production. Fuel 2022, 313, 123017. [Google Scholar] [CrossRef]
- Schollaen, K.; Baschek, H.; Heinrich, I.; Slotta, F.; Pauly, M.; Helle, G. A guideline for sample preparation in modern tree-ring stable isotope research. Dendrochronologia 2017, 44, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Delmás, M.; Rich, S.; Traoré, M.; Hajj, F.; Poszwa, A.; Akhmetzyanov, L.; García-González, I.; Groenendijk, P. Tree-ring chronologies, stable strontium isotopes and biochemical compounds: Towards reference datasets to provenance Iberian shipwreck timbers. J. Archaeol. Sci. Rep. 2020, 34, 102640. [Google Scholar] [CrossRef]
- Konaré, D. Équations allométriques pour l’évaluation de la biomasse foliaire de trois espèces ligneuses fourragères. Cas de Afzelia africana, Ficus gnaphalocarpa et Pterocarpus erinaceus, des parcours naturels du cercle de Kéniéba au Sud-Ouest du Mali. Rev. Afr. Des Sci. Soc. Et De La Sante Publique 2019, 1, 29–52. [Google Scholar]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Simons, A. Agroforestree Database: A Tree Reference and Selection Guide Version 4.0.; World Agroforestry Centre: Nairobi, Kenya, 2009. [Google Scholar]
- Segla, K.N.; Adjonou, K.; Rabiou, H.; André Bationo, B.; Mahamane, A.; Guibal, D.; Kokou, K.; Chaix, G.; Kokutse, A.D.; Langbour, P. Relations between the ecological conditions and the properties of Pterocarpus erinaceus Poir. wood from the Guinean-Sudanian and Sahelian zones of West Africa. Holzforschung 2020, 74, 999–1009. [Google Scholar] [CrossRef]
- Sanogo, K.; Gebrekirstos, A.; Bayala, J.; van Noordwijk, M. Climate-growth relationships of Daniellia oliveri (Rolfe) Hutch. & Dalziel in the Sudanian zone of Mali, West Africa. Trees For. People 2022, 10, 100333. [Google Scholar]
- Hänke, H.; Börjeson, L.; Hylander, K.; Enfors-Kautsky, E. Drought tolerant species dominate as rainfall and tree cover returns in the West African Sahel. Land Use Policy 2016, 59, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Hérault, B.; N’guessan, A.K.; Ahoba, A.; Bénédet, F.; Coulibaly, B.; Doua-Bi, Y.; Koffi, T.; Koffi-Konan, J.C.; Konaté, I.; Tiéoulé, F.; et al. The long-term performance of 35 tree species of sudanian West Africa in pure and mixed plantings. For. Ecol. Manag. 2020, 468, 118171. [Google Scholar] [CrossRef]
- Brandt, M.; Romankiewicz, C.; Spiekermann, R.; Samimi, C. Environmental change in time series–An interdisciplinary study in the Sahel of Mali and Senegal. J. Arid Environ. 2014, 105, 52–63. [Google Scholar] [CrossRef]
- Faye, M.D.; Weber, J.C.; Abasse, T.A.; Boureima, M.; Larwanou, M.; Bationo, A.B.; Diallo, B.O.; Sigué, H.; Dakouo, J.M.; Samaké, O.; et al. Farmers’ preferences for tree functions and species in the West African Sahel. For. Trees Livelihoods 2011, 20, 113–136. [Google Scholar] [CrossRef]
- World Conservation Monitoring Centre. Khaya senegalensis. In The IUCN Red List of Threatened Species; e.T32171A9684583; World Conservation Monitoring Centre: Cambridge, UK, 1998. [Google Scholar]
- Contu, S. Isoberlinia doka. In The IUCN Red List of Threatened Species; e.T19892774A20090324; World Conservation Monitoring Centre: Cambridge, UK, 2012. [Google Scholar]
- Barstow, M. Pterocarpus erinaceus. In The IUCN Red List of Threatened Species; e.T62027797A62027800; World Conservation Monitoring Centre: Cambridge, UK, 2018. [Google Scholar]
- Botanic Gardens Conservation International (BGCI); IUCN SSC Global Tree Specialist Group. Daniellia oliveri. In The IUCN Red List of Threatened Species; e.T144315321A149026856; World Conservation Monitoring Centre: Cambridge, UK, 2019. [Google Scholar]
- Ky-Dembele, C.; Bayala, J.; Kalinganire, A.; Traoré, F.T.; Koné, B.; Olivier, A. Vegetative propagation of twelve fodder tree species indigenous to the Sahel, West Africa. South. For. A J. For. Sci. 2016, 78, 185–192. [Google Scholar] [CrossRef]
- Martínez Cortizas, A.; López-Costas, O.; Orme, L.; Mighall, T.; Kylander, M.E.; Bindler, R.; Gallego Sala, Á. Holocene atmospheric dust deposition in NW Spain. Holocene 2020, 30, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Traoré, M.; Kaal, J.; Martínez Cortizas, A. Variation of wood color and chemical composition in the stem cross-section of oak (Quercus spp.) trees, with special attention to the sapwood-heartwood transition zone. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2023, 285, 121893. [Google Scholar] [CrossRef] [PubMed]
- Álvarez Fernández, N.; Martínez Cortizas, A. Andurinha: Make Spectroscopic Data Processing Easier; R Package Version 0.0.2.; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Auxenfans, T.; Crônier, D.; Chabbert, B.; Paës, G. Understanding the structural and chemical changes of plant biomass following steam explosion pretreatment. Biotechnol. Biofuels 2017, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, M.C.; Popescu, C.M.; Lisa, G.; Sakata, Y. Evaluation of morphological and chemical aspects of different wood species by spectroscopy and thermal methods. J. Mol. Struct. 2011, 988, 65–72. [Google Scholar] [CrossRef]
- Klumpers, J.; Janin, G.; Becker, M.; Lévy, G. The influences of age, extractive content and soil water on wood color in oak: The possible genetic determination of wood color. In Annales des Sciences Forestières; EDP Sciences: Les Ulis, France, 1993; Volume 50, pp. 403s–409s. [Google Scholar]
- Gierlinger, N.; Jacques, D.; Grabner, M.; Wimmer, R.; Schwanninger, M.; Rozenberg, P.; Pâques, L.E. Colour of larch heartwood and relationships to extractives and brown-rot decay resistance. Trees 2004, 18, 102–108. [Google Scholar] [CrossRef]
- Moya, R.; Berrocal, A. Wood colour variation in sapwood and heartwood of young trees of Tectona grandis and its relationship with plantation characteristics, site, and decay resistance. Ann. For. Sci. 2010, 67, 109. [Google Scholar] [CrossRef] [Green Version]
- Moya, R.; Fallas, R.S.; Bonilla, P.J.; Tenorio, C. Relationship between wood color parameters measured by the CIELab system and extractive and phenol content in Acacia mangium and Vochysia guatemalensis from fast-growth plantations. Molecules 2012, 17, 3639–3652. [Google Scholar] [CrossRef] [Green Version]
- Nelson, N.D.; Maeglin, R.R.; Wahlgren, H.E. Relationship of black walnut wood color to soil properties and site. Wood Fiber Sci. 1969, 1, 29–37. [Google Scholar]
- Sotelo Montes, C.; Weber, J.C.; Silva, D.A.; Andrade, C.; Muñiz, G.I.; Garcia, R.A.; Kalinganire, A. Effects of region, soil, land use, and terrain type on fuelwood properties of five tree/shrub species in the Sahelian and Sudanian ecozones of Mali. Ann. For. Sci. 2012, 69, 747–756. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, F.; Kang, X. Chemical properties of wood are under stronger genetic control than growth traits in Populus tomentosa Carr. Ann. For. Sci. 2015, 72, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Segla, K.N.; Kokutse, A.D.; Adjonou, K.; Langbour, P.; Chaix, G.; Guibal, D.; Kokou, K. Caractéristiques biophysiques du bois de Pterocarpus erinaceus (Poir.) en zones guinéenne et soudanienne au Togo. Bois Forêts Trop. 2015, 324, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Creber, G.T.; Chaloner, W.G. Influence of environmental factors on the wood structure of living and fossil trees. Bot. Rev. 1984, 50, 357–448. [Google Scholar] [CrossRef]
- Nelson, M.L.; O’Connor, R.T. Relation of certain infrared bands to cellulose crystallinity and crystal lattice type. Part II. A new infrared ratio for estimation of crystallinity in celluloses I and II. J. Appl. Polym. Sci. 1964, 8, 1325–1341. [Google Scholar] [CrossRef]
- Colom, X.; Carrillo, F. Comparative study of wood samples of the northern area of Catalonia by FTIR. J. Wood Chem. Technol. 2005, 25, 1–11. [Google Scholar] [CrossRef]
- Popescu, C.M.; Singurel, G.; Popescu, M.C.; Vasile, C.; Argyropoulos, D.S.; Willför, S. Vibrational spectroscopy and X-ray diffraction methods to establish the differences between hardwood and softwood. Carbohydr. Polym. 2009, 77, 851–857. [Google Scholar] [CrossRef]
- Pandey, K.K. A note on the influence of extractives on the photo-discoloration and photo-degradation of wood. Polym. Degrad. Stab. 2005, 87, 375–379. [Google Scholar] [CrossRef]
- Whetten, R.W.; MacKay, J.J.; Sederoff, R.R. Recent advances in understanding lignin biosynthesis. Annu. Rev. Plant Biol. 1998, 49, 585–609. [Google Scholar] [CrossRef]
- Novaes, E.; Kirst, M.; Chiang, V.; Winter-Sederoff, H.; Sederoff, R. Lignin and Biomass: A Negative Correlation for Wood Formation and Lignin Content in Trees. Plant Physiol. 2010, 154, 555–561. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wood Species | Kéniéba | Kita | Sibi | Total |
|---|---|---|---|---|
| Daniellia oliveri | 6 | 5 | 6 | 17 |
| Isoberlinia doka | 5 | 5 | - | 10 |
| Khaya senegalensis | 8 | 8 | 8 | 24 |
| Pterocarpus erinaceus | 6 | 6 | 5 | 17 |
| Total | 68 | |||
| Wood Species | Color Parameters | Mean | SD | df | F | Sig. |
|---|---|---|---|---|---|---|
| Daniellia oliveri | Lightness (L*) | 60.14 | 2.10 | 2 | 0.78 | 0.48 |
| Redness (a*) | 8.24 | 0.29 | 2 | 3.01 | 0.08 | |
| Yellowness (b*) | 25.33 | 1.64 | 2 | 0.58 | 0.57 | |
| Chroma (C*) | 26.65 | 1.57 | 2 | 0.54 | 0.59 | |
| Hue (h°) | 71.92 | 1.23 | 2 | 1.39 | 0.28 | |
| Isoberlinia doka | Lightness (L*) | 47.82 | 4.12 | 1 | 0.12 | 0.74 |
| Redness (a*) | 14.88 | 0.58 | 1 | 0.47 | 0.51 | |
| Yellowness (b*) | 18.08 | 0.93 | 1 | 0.33 | 0.58 | |
| Chroma (C*) | 23.42 | 0.90 | 1 | 0.03 | 0.87 | |
| Hue (h°) | 50.52 | 1.54 | 1 | 1.12 | 0.32 | |
| Khaya senegalensis | Lightness (L*) | 50.86 | 3.18 | 2 | 2.68 | 0.09 |
| Redness (a*) | 15.53 | 0.52 | 2 | 0.70 | 0.51 | |
| Yellowness (b*) | 19.09 | 0.66 | 2 | 7.63 | 0.00 (1) | |
| Chroma (C*) | 24.61 | 0.66 | 2 | 2.16 | 0.14 | |
| Hue (h°) | 50.87 | 1.23 | 2 | 9.11 | 0.00 (1) | |
| Pterocarpus erinaceus | Lightness (L*) | 55.76 | 3.20 | 2 | 4.25 | 0.04 (2) |
| Redness (a*) | 11.87 | 1.09 | 2 | 8.29 | 0.00 (1) | |
| Yellowness (b*) | 23.83 | 0.85 | 2 | 0.37 | 0.70 | |
| Chroma (C*) | 26.65 | 0.62 | 2 | 0.91 | 0.43 | |
| Hue (h°) | 63.52 | 2.64 | 2 | 4.91 | 0.02 (2) |
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
|---|---|---|---|---|---|---|
| Bands with positive loadings | 1623 (0.94) | 822 (0.91) | 2922 (0.92) | 1019 (0.96) | 1422 (0.79) | 1107 (0.77) |
| 1606 (0.87) | 1245 (0.91) | 2935 (0.92) | 986 (0.91) | 1543 (0.76) | 1124 (0.61) | |
| 1592 (0.86) | 1228 (0.89) | 2853 (0.89) | 1031 (0.91) | 1560 (0.67) | ||
| 1318 (0.74) | 1264 (0.82) | 1055 (0.71) | 1375 (0.55) | |||
| 895 (0.78) | 1159 (0.52) | |||||
| 1510 (0.64) | ||||||
| 1461 (0.64) | ||||||
| Bands with negative loadings | 1735 (−0.86) | 1655 (−0.70) | ||||
| 1720 (−0.86) | 3323 (−0.62) | |||||
| 1703 (−0.83) | ||||||
| Wood Species | PCA Component | df | F | Sig. |
|---|---|---|---|---|
| Daniellia oliveri | PC1 | 2 | 0.21 | 0.81 |
| PC2 | 2 | 0.41 | 0.67 | |
| PC3 | 2 | 0.42 | 0.66 | |
| PC4 | 2 | 6.99 | 0.01 (1) | |
| PC5 | 2 | 0.44 | 0.65 | |
| PC6 | 2 | 3.03 | 0.08 | |
| Isoberlinia doka | PC1 | 1 | 1.54 | 0.25 |
| PC2 | 1 | 0.01 | 0.92 | |
| PC3 | 1 | 0.15 | 0.71 | |
| PC4 | 1 | 0.00 | 0.99 | |
| PC5 | 1 | 2.93 | 0.13 | |
| PC6 | 1 | 0.35 | 0.57 | |
| Khaya senegalensis | PC1 | 2 | 1.39 | 0.27 |
| PC2 | 2 | 1.30 | 0.29 | |
| PC3 | 2 | 5.50 | 0.01 (2) | |
| PC4 | 2 | 4.51 | 0.02 (2) | |
| PC5 | 2 | 0.49 | 0.62 | |
| PC6 | 2 | 2.25 | 0.13 | |
| Pterocarpus erinaceus | PC1 | 2 | 3.50 | 0.06 |
| PC2 | 2 | 3.40 | 0.06 | |
| PC3 | 2 | 2.87 | 0.09 | |
| PC4 | 2 | 1.15 | 0.35 | |
| PC5 | 2 | 12.08 | 0.00 (1) | |
| PC6 | 2 | 4.56 | 0.03 (2) |
| Wood Species | FTIR Ratios | df | F | Sig. |
|---|---|---|---|---|
| Daniellia oliveri | 1425/895 (LOI) | 2 | 0.72 | 0.50 |
| 1265/1230 (GS1) | 2 | 1.18 | 0.34 | |
| 1510/1595 (GS2) | 2 | 4.53 | 0.03 (2) | |
| 1510/1460 (LC1) | 2 | 5.59 | 0.02 (2) | |
| 1510/1375 (LC2) | 2 | 4.34 | 0.03 (2) | |
| 1510/895 (LC3) | 2 | 4.26 | 0.04 (2) | |
| Isoberlinia doka | 1425/895 (LOI) | 1 | 2.38 | 0.16 |
| 1265/1230 (GS1) | 1 | 0.01 | 0.94 | |
| 1510/1595 (GS2) | 1 | 0.22 | 0.65 | |
| 1510/1460 (LC1) | 1 | 0.00 | 0.96 | |
| 1510/1375 (LC2) | 1 | 0.32 | 0.59 | |
| 1510/895 (LC3) | 1 | 0.32 | 0.59 | |
| Khaya senegalensis | 1425/895 (LOI) | 2 | 0.40 | 0.67 |
| 1265/1230 (GS1) | 2 | 3.42 | 0.05 | |
| 1510/1595 (GS2) | 2 | 2.43 | 0.11 | |
| 1510/1460 (LC1) | 2 | 0.90 | 0.42 | |
| 1510/1375 (LC2) | 2 | 1.80 | 0.19 | |
| 1510/895 (LC3) | 2 | 1.40 | 0.27 | |
| Pterocarpus erinaceus | 1425/895 (LOI) | 2 | 18.24 | 0.00 (1) |
| 1265/1230 (GS1) | 2 | 18.23 | 0.00 (1) | |
| 1510/1595 (GS2) | 2 | 6.01 | 0.01 (2) | |
| 1510/1460 (LC1) | 2 | 5.29 | 0.02 (2) | |
| 1510/1375 (LC2) | 2 | 3.65 | 0.05 | |
| 1510/895 (LC3) | 2 | 8.19 | 0.00 (1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Traoré, M.; Martínez Cortizas, A. Color and Chemical Composition of Timber Woods (Daniellia oliveri, Isoberlinia doka, Khaya senegalensis, and Pterocarpus erinaceus) from Different Locations in Southern Mali. Forests 2023, 14, 767. https://0-doi-org.brum.beds.ac.uk/10.3390/f14040767
Traoré M, Martínez Cortizas A. Color and Chemical Composition of Timber Woods (Daniellia oliveri, Isoberlinia doka, Khaya senegalensis, and Pterocarpus erinaceus) from Different Locations in Southern Mali. Forests. 2023; 14(4):767. https://0-doi-org.brum.beds.ac.uk/10.3390/f14040767
Chicago/Turabian StyleTraoré, Mohamed, and Antonio Martínez Cortizas. 2023. "Color and Chemical Composition of Timber Woods (Daniellia oliveri, Isoberlinia doka, Khaya senegalensis, and Pterocarpus erinaceus) from Different Locations in Southern Mali" Forests 14, no. 4: 767. https://0-doi-org.brum.beds.ac.uk/10.3390/f14040767