Estimating the Annual Above-Ground Biomass Production of Various Species on Sites in Sweden on the Basis of Individual Climate and Productivity Values

Abstract
:1. Introduction
2. Objectives
3. Materials and Methods
3.1. The Modified Miami Model



- (1)
- MAT ≥ 0 °C: this model gives an estimate of ABM, starting at a MAT of 0 °C. It is not valid for values below 0 °C.
- (2)
- MATnat > 0 °C: the MATnat must be positive because of the mathematical formulation.
3.2. Validation of the Modified Miami Model

{kind=link}
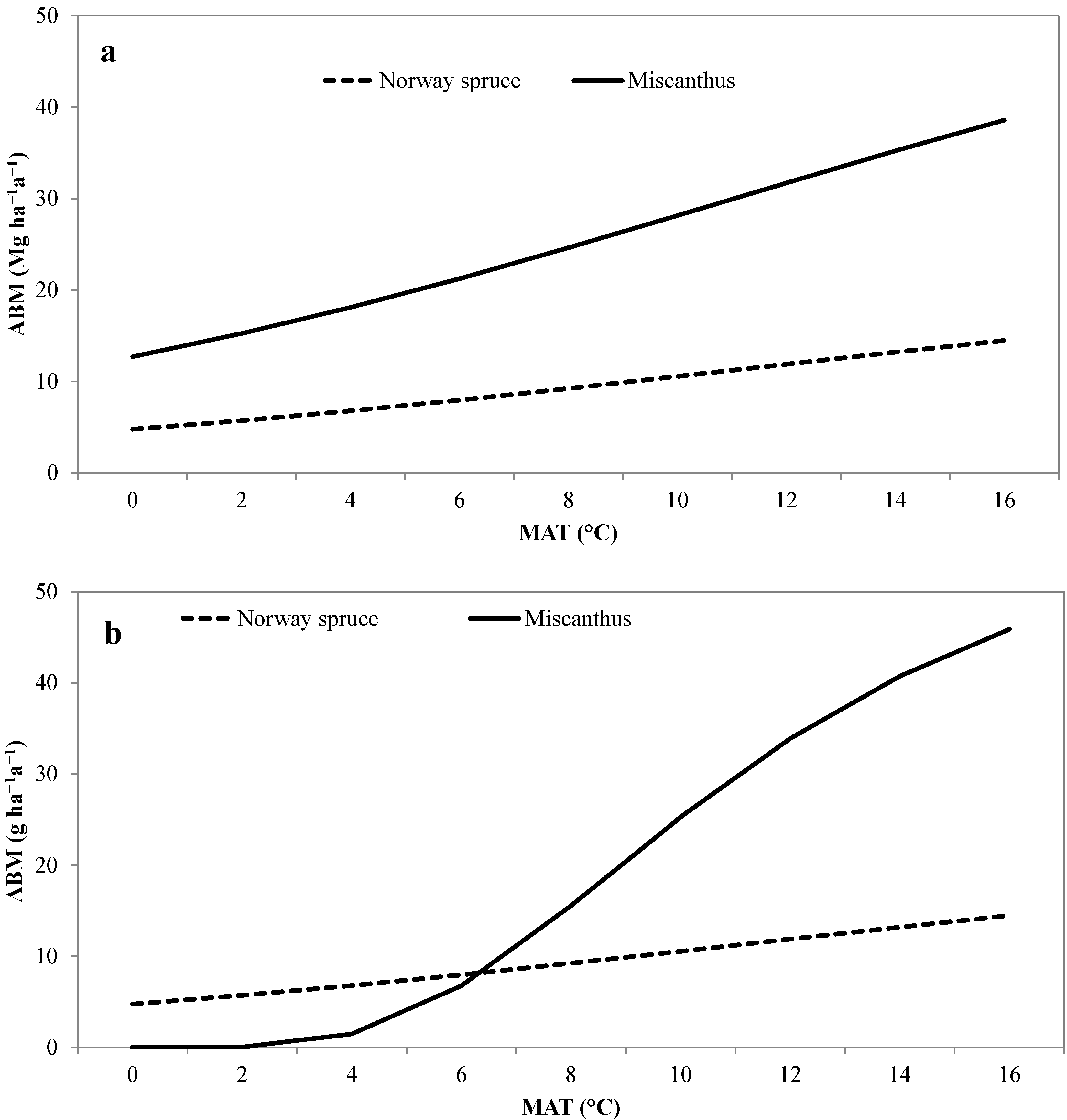{kind=link}
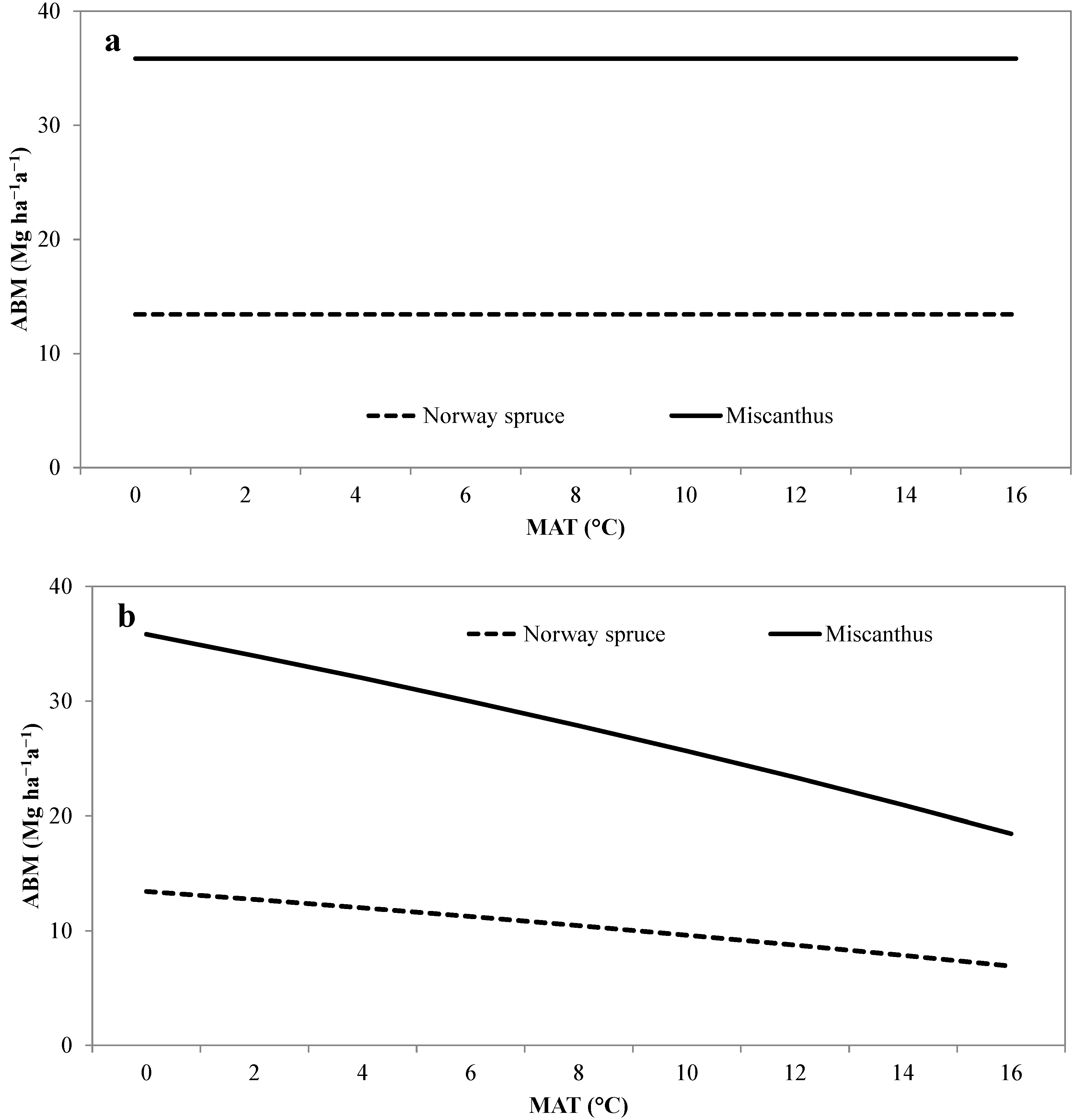{kind=link}
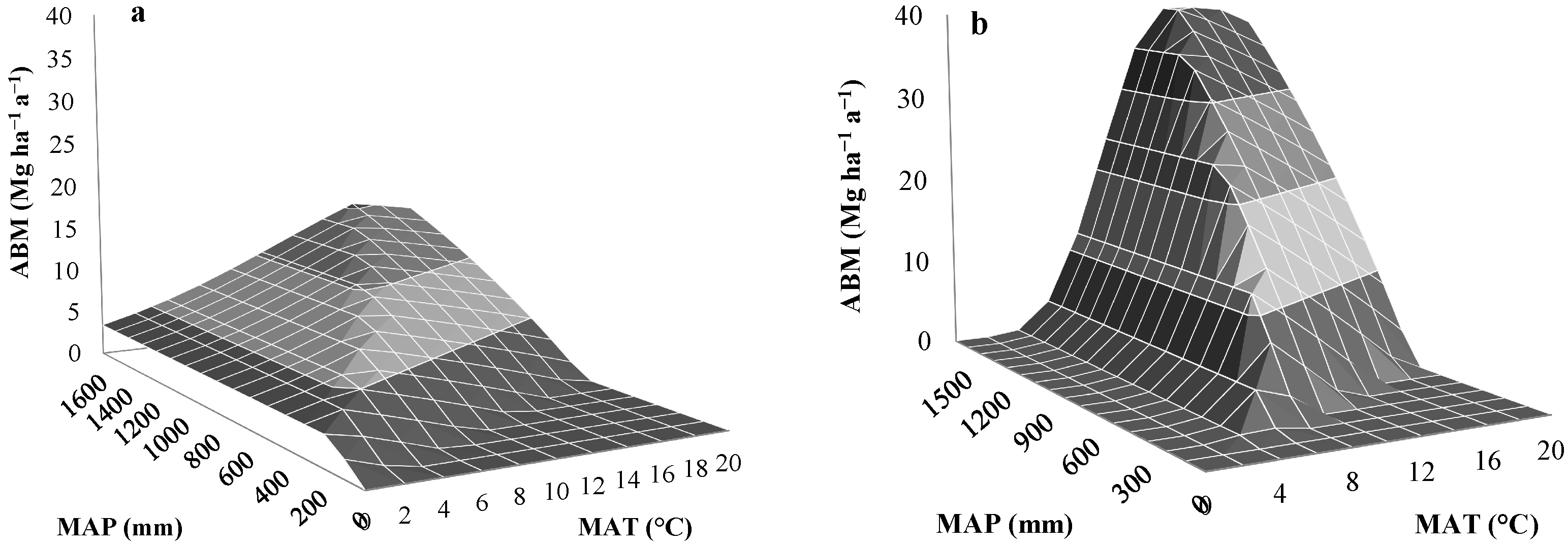{kind=link}
{kind=link}
| Literature data | Parameters | Output | ||||||
|---|---|---|---|---|---|---|---|---|
| Species | MAT | MAP | BMmax | BMmax | MATnat | ABM | r2 | References |
| °C | mm | Mg ha−1a−1 | Mg ha−1a−1 | °C | Mg ha−1a−1 | - | ||
| Norway spruce | 0.0 | 718 * | 4.5 | 15 | 1 | 4.8 | 0.90 | [20,35,36,37] |
| 2.0 | 718 * | 6.2 | 5.7 | |||||
| 4.0 | 718 * | 7.9 | 6.8 | |||||
| 6.0 | 718 * | 9.6 | 8.0 | |||||
| 8.0 | 718 * | 11.3 | 8.0 | |||||
| 10.0 | 718 * | 13.0 | 7.0 | |||||
| Reed canary grass | 4.6 | 632 | 6.4 | 14 | 2 | 6.0 | 0.58 | [31,38,39,40,41,42] |
| 5.9 | 750 | 6.2 | 7.0 | |||||
| 6.9 | 657 | 6.0 | 7.1 | |||||
| 7.6 | 514 | 5.5 | 4.7 | |||||
| 8.2 | 477 | 6.2 | 3.8 | |||||
| 8.4 | 547 | 6.2 | 4.8 | |||||
| Miscanthus | 7.6 | 783 | 11.8 | 40 | 18 | 13.7 | 0.41 | [23,24,34,43,44] |
| 7.8 | 578 | 15.1 | 14.6 | |||||
| 8.3 | 630 | 14.7 | 17.0 | |||||
| 8.9 | 634 | 15.4 | 16.7 | |||||
| 9.6 | 689 | 18.5 | 18.1 | |||||
| 9.6 | 769 | 12.6 | 21.2 | |||||
| 9.6 | 766 | 17.5 | 21.1 | |||||
| 9.8 | 660 | 21.0 | 16.6 | |||||
| 10.4 | 594 | 19.6 | 12.9 | |||||
| 10.4 | 594 | 12.5 | 12.9 | |||||
| 10.5 | 676 | 19.6 | 16.3 | |||||
| 11.5 | 813 | 34.3 | 20.6 | |||||
| * Estimated mean values. | ||||||||
- (1)
- The literature data were compared to modeled data using each single MATnat and BMmax value in the literature directly as a species-specific parameter. The coefficient of determination of this modification was r2 = 0.903 with p < 0.0001.
- (2)
- The literature data were compared to the modeled data using as parameters the average MATnat and BMmax values from the literature data. The coefficient of determination of this modification was r2 = 0.759 with p < 0.0001.

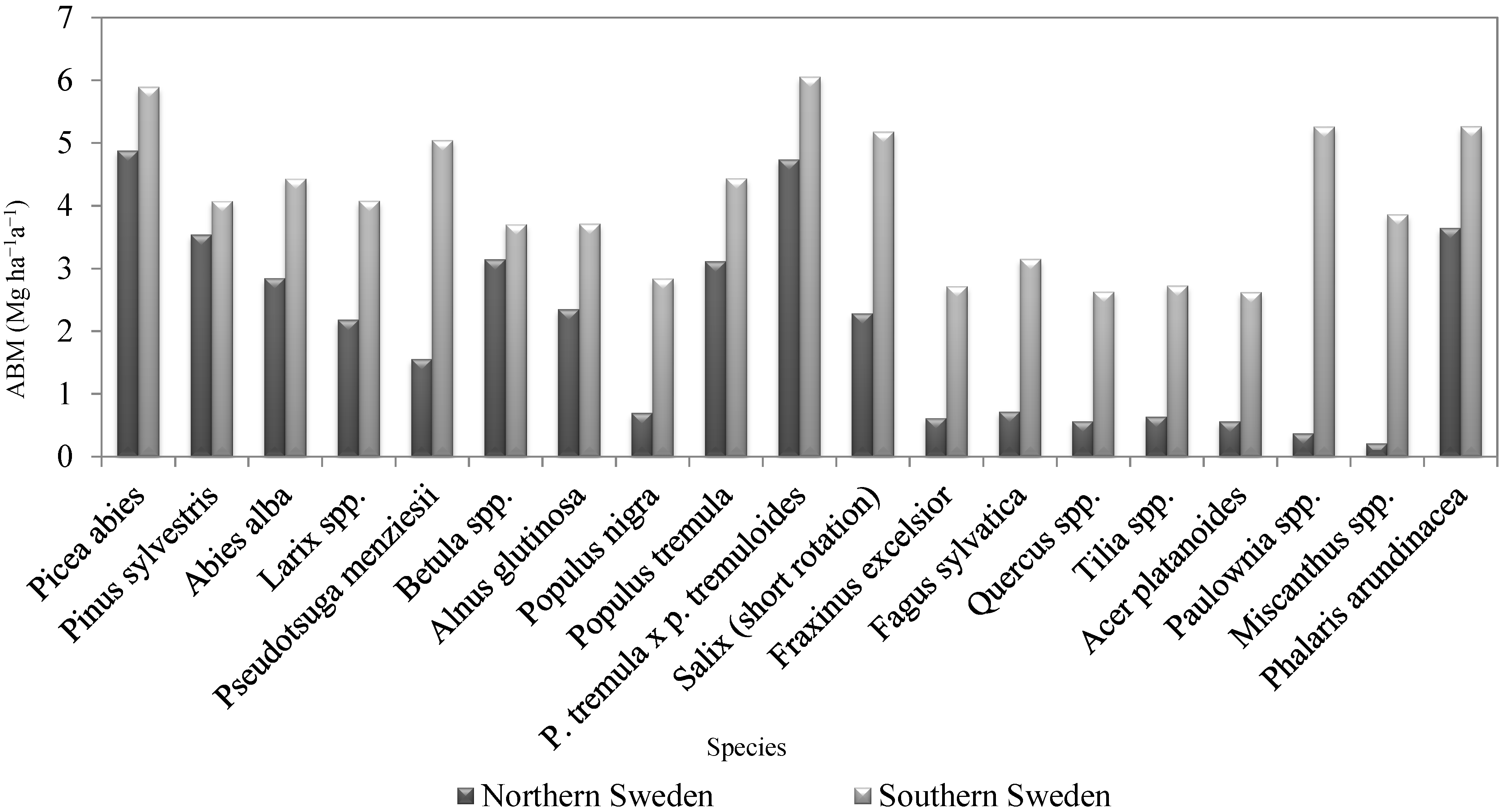
3.3. Choice of Species-Specific Parameters for the Modified MIAMI Model to Estimate the Above-Ground Biomass Production in Dry Matter for Different Selected Species
| Species | BMmax | MATnat | References |
|---|---|---|---|
| Mg ha−1a−1 | °C | ||
| Picea abies Karst | 10.8 | 0.5 | [48,49,50,51,52,53] |
| Pinus sylvestris L. | 7.3 | 0.1 | [50,51,52,53,54,55] |
| Abies alba Mill. | 9.1 | 2.6 | [50,51,52,53,54,55,56,57] |
| Larix spp. | 9.1 | 3.9 | [50,51,52,58,59,60,61,62] |
| Pseudotsuga menziesii Mirb. Franco | 14.2 | 7.6 | [52,63,64,65,66] |
| Betula spp. | 6.7 | 0.3 | [50,51,53,55,67] |
| Alnus glutinosa (L.) Gaertn. | 7.7 | 2.7 | [51,53,68] |
| Populus nigra L. | 8.8 | 9.1 | [69,70,71,72,73,74,75] |
| Populus tremula L. | 8.8 | 1.9 | [50,53,67,71,76,77,78,79] |
| P. tremula L. x p. tremuloides Michx. | 11.4 | 1.0 | [71,76,80,81] |
| Salix spp. (short rotation) | 12.5 | 5.3 | [50,71,74,75,82,83] |
| Fraxinus excelsior L. | 8.7 | 9.6 | [51,55,75,84,85,86,87] |
| Fagus sylvatica L. | 10.0 | 9.5 | [51,52,55,73,84,85,86,87] |
| Quercus spp. | 7.4 | 9.9 | [51,52,73,75,84,85,86] |
| Tilia spp. | 8.3 | 9.4 | [84,88,89] |
| Acer platanoides L. | 7.0 | 9.9 | [51,75,84,85,87] |
| Paulownia spp. | 28.3 | 16.8 | [26,32,90,91] |
| Miscanthus spp. | 23.5 | 18.4 | [33,34,44,71,92,93] |
| Phalaris arundinacea L. | 10.5 | 2.0 | [31,41,42,94,95,96] |
4. Results and Discussion

5. Conclusions
Conflicts of Interest
References
- Lévêque, C. Ecology: From Ecosystem to Biosphere; BIOS Scientific Publishers Limited: Milton Park, UK, 2003; p. 472. [Google Scholar]
- Nebel, B.J.; Wright, R.T. Environmental Science: The Way the World Works; Prentice Hall: Upper Saddle River, NJ, USA, 1993; p. 630. [Google Scholar]
- Hoy, M.A. Classical biological control. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Gainesville, FL, USA, 2008; Volume 2, pp. 906–923. [Google Scholar]
- Orcutt, D.M.; Nilsen, E.T. The Physiology of Plants under Stress: Soil and Biotic Factors; Wiley: Hoboken, NJ, USA, 2000; Volume 2, p. 696. [Google Scholar]
- Ellenberg, H. Vegetation Mitteleuropas mit den Alpen: In ökologischer Sicht; Ulmer: Stuttgart, Germany, 1986; p. 989. [Google Scholar]
- Burschel, P.; Huss, J. Grundriss des Waldbaus: Ein Leitfaden für Studium und Praxis; Eugen Ulmer Verlag: Stuttgart, Germany, 1987; p. 487. [Google Scholar]
- Akhtar, M.; Jaiswal, A.; Taj, G.; Jaiswal, J.P.; Qureshi, M.I.; Singh, N.K. DREB1/CBF transcription factors: Their structure, function and role in abiotic stress tolerance in plants. J. Genet. 2012, 91, 385–395. [Google Scholar] [CrossRef]
- Benito-Garzón, M.; Ruiz-Benito, P.; Zavala, M.A. Interspecific differences in tree growth and mortality responses to environmental drivers determine potential species distributional limits in Iberian forests. Glob. Ecol. Biogeogr. 2013, 22, 1141–1151. [Google Scholar] [CrossRef]
- Boucher-Lalonde, V.; Morin, A.; Currie, D.J. How are tree species distributed in climatic space? A simple and general pattern. Glob. Ecol. Biogeogr. 2012, 21, 1157–1166. [Google Scholar] [CrossRef]
- Pellissier, L.; Anne Bråthen, K.; Pottier, J.; Randin, C.F.; Vittoz, P.; Dubuis, A.; Yoccoz, N.G.; Alm, T.; Zimmermann, N.E.; Guisan, A.; et al. Species distribution models reveal apparent competitive and facilitative effects of a dominant species on the distribution of tundra plants. Ecography 2010, 33, 1004–1014. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Strayer, D.L. Usefulness of bioclimatic models for studying climate change and invasive species. In The Year in Ecology and Conservation Biology; Ostfeld, R.S., Schlesinger, W.H., Eds.; Blackwell Scientific Publishing: Boston, MA, USA, 2008; p. 1134. [Google Scholar]
- Sykes, M.T. Modelling the potential distribution and community dynamics of lodgepole pine (Pinus contorta Dougl. ex. Loud.) in Scandinavia. For. Ecol. Manag. 2001, 141, 69–84. [Google Scholar] [CrossRef]
- Sykes, M.T.; Prentice, I.C.; Cramer, W. A bioclimatic model for the potential distributions of north European tree species under present and future climates. J. Biogeogr. 1996, 23, 203–233. [Google Scholar]
- Lieth, H. Modeling the primary productivity of the world. In Primary Productivity of the Biosphere; Lieth, H., Whittaker, R.H., Eds.; Springer-Verlag: New York, NY, USA, 1975; pp. 237–263. [Google Scholar]
- Schuur, E.A.G. Productivity and global climate revisited: The sensitivity of tropical forest growth to precipitation. Ecology 2003, 84, 1165–1170. [Google Scholar] [CrossRef]
- Grosso, S.D.; Parton, W.; Stohlgren, T.; Zheng, D.; Bachelet, D.; Prince, S.; Hibbard, K.; Olson, R. Global potential net primary production predicted from vegetation class, precipitation, and temperature. Ecology 2008, 89, 2117–2126. [Google Scholar] [CrossRef]
- Shoo, L.P.; Ramirez, V.V. Global potential net primary production predicted from vegetation class, precipitation, and temperature: Comment. Ecology 2010, 91, 921–923. [Google Scholar] [CrossRef]
- Zha, T.S.; Barr, A.G.; Bernier, P.Y.; Lavigne, M.B.; Trofymow, J.A.; Amiro, B.D.; Arain, M.A.; Bhatti, J.S.; Black, T.A.; Margolis, H.A.; et al. Gross and aboveground net primary production at Canadian forest carbon flux sites. Agric. For. Meteorol. 2013, 174–175, 54–64. [Google Scholar] [CrossRef]
- Waring, R.H.; Landsberg, J.J.; Williams, M. Net primary production of forests: A constant fraction of gross primary production? Tree Physiol. 1998, 18, 129–134. [Google Scholar] [CrossRef]
- Bergh, J.; Linder, S.; Bergström, J. Potential production of Norway spruce in Sweden. For. Ecol. Manag. 2005, 204, 1–10. [Google Scholar] [CrossRef]
- Troedsson, T.; Nykvist, N. Marklära och Markvård; Almqvist & Wiksell: Stockholm, Sweden, 1973; p. 403. [Google Scholar]
- Troedsson, T. Mark, Marksligtage och Markhushållning; LTs förlag: Helsingborg, Sweden, 1975; p. 167. [Google Scholar]
- McKervey, Z.; Woods, V.B.; Easson, D.L. Miscanthus as an Energy Crop and Its Potential for Northern Ireland: A Review of Current Knowledge; Agri-Food and Biosciences Institute, Global Research Unit: Hillsborough, UK, 2008; p. 72. [Google Scholar]
- Greef, J.M.; Deuter, M.; Jung, C.; Schondelmaier, J. Genetic diversity of European Miscanthus species revealed by AFLP fingerprinting. Genet. Resour. Crop Evol. 1997, 44, 185–195. [Google Scholar] [CrossRef]
- Komatsu, H.; Maita, E.; Otsuki, K. A model to estimate annual forest evapotranspiration in Japan from mean annual temperature. J. Hydrol. 2008, 348, 330–340. [Google Scholar] [CrossRef]
- Zhu, Z.-H.; Chao, C.-J.; Lu, X.-Y.; Xiong, Y.G. Paulownia in China: Cultivation and Utilization; Chinese Academy of Forestry/Asian Network for Biological Sciences/International Development Research Center: Beijing, China, 1986; p. 65. [Google Scholar]
- Schröder, B. Paulownien für die Plantagenwirtschaft—Characteristikum der Paulownia Sorten. Available online: http://www.paulownia-baumschule.de/forstpflanzen/ (accessed on 12 April 2013).
- Helfer, F.; Lemckert, C.; Zhang, H. Impacts of climate change on temperature and evaporation from a large reservoir in Australia. J. Hydrol. 2012, 475, 365–378. [Google Scholar] [CrossRef]
- Spiecker, H. The growth of Norway spruce (Picea abies [L.] Karst.) in Europe within and beyond its natural range. In Forest Ecosystem Restoration, Proceedings of the International Conference, Vienna, Austria, 10–12 April 2000; Hasenauer, H., Ed.; Institute of Forest Growth Research: Wien, Austria, 2000; pp. 247–256. [Google Scholar]
- Lavergne, S.; Molofsky, J. Reed canary grass (Phalaris arundinacea) as a biological model in the study of plant invasions. Crit. Rev. Plant Sci. 2004, 23, 415–429. [Google Scholar] [CrossRef]
- Strašil, Z. Evaluation of reed canary grass (Phalaris arundinacea L.) grown for energy use. Res. Agric. Eng. 2012, 58, 119–130. [Google Scholar]
- Lawrence, J.S. TGG Paulownia Biomass Production; Toad Gully Growers: Melbourne, Australia, 2011; p. 2. [Google Scholar]
- Nishiwaki, A.; Mizuguti, A.; Kuwabara, S.; Toma, Y.; Ishigaki, G.; Miyashita, T.; Yamada, T.; Matuura, H.; Yamaguchi, S.; Rayburn, A.L.; et al. Discovery of natural miscanthus (Poaceae) triploid plants in sympatric populations of Miscanthus sacchariflorus and Miscanthus sinensis in southern Japan. Am. J. Bot. 2011, 98, 154–159. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Lewandowski, I.; Andersson, B.; Basch, G.; Christian, D.G.; Kjeldsen, J.B.; Jørgensen, U.; Mortensen, J.V.; Riche, A.B.; Schwarz, K.-U.; et al. Performance of 15 miscanthus genotypes at five sites in Europe. Agron. J. 2001, 93, 1013–1019. [Google Scholar] [CrossRef]
- Schulze, E.D. Carbon and nitrogen cycling in European forest ecosystems. In Ecological Studies; Schulze, E.D., Ed.; Springer-Verlag: Heidelberg, Germany, 2000; Volume 142, pp. 3–14. [Google Scholar]
- Virkesförrådet Fördelat på Trädslag Inom Diameterklasser. Exkl Torra och Vindfällda Träd. Alla Ägoslag. 2006–2010; SLU (Institutionen för skoglig resurshushållning), Sveriges officiella statistik: Umeå, Sweden, 2011; p. 128.
- SMHI Årsmedelnederbörden sedan 1860. Available online: http://www.smhi.se/klimatdata/meteorologi/nederbord/1.2887 (accessed on 22 March 2013).
- Xiong, S.; Lötjönen, T.; Knuuttila, K. Energiproduktion. från Rörflen: Handbok för el och Värmeproduktion (Energy Conversion with Reed Canary Grass: A Guide for Electricity and Heat Production); Swedish University of Agricultures, Unit of Biomass Technology and Chemistry: Umeå, Sweden, 2008; p. 36. [Google Scholar]
- Carlsson, E. Rörflen som Alternativt Strukturfoder och Strömedel—Är Rörflen ett Ekonomiskt Alternativ som Strukturfoder och Strömedel? Sveriges Lantbruksuniversitet, Fakulteten för Lantskapsplanering, Trädgårds-och Jordbruksvetenskap: Alnarp, Sweden, 2012; p. 25. [Google Scholar]
- Pahkala, K.; Aalto, M.; Isolahti, M.; Poikola, J.; Jauhiainen, L. Large-scale energy grass farming for power plants—A case study from Ostrobothnia, Finland. Biomass Bioenergy 2008, 32, 1009–1015. [Google Scholar] [CrossRef]
- Pahkala, K.; Pihala, M. Different plant parts as raw material for fuel and pulp production. Ind. Crops Prod. 2000, 11, 119–128. [Google Scholar] [CrossRef]
- Jansone, B.; Rancane, S.; Berzins, P.; Stesele, V. Reed canary grass (Phalaris Arundinacea L.) in natural biocenosis of Latvia, research experiments and production fields. In Proceedings of the International Scientific Conference: Renewable Energy and Energy Efficiency, Jelgava, Latvia, 28–30 May 2012; Latvia University of Agriculture: Jelgava, Latvia, 2012; pp. 61–65. [Google Scholar]
- Lewandowski, I.; Clifton-Brown, J.C.; Andersson, B.; Basch, G.; Christian, D.G.; Jørgensen, U.; Jones, M.B.; Riche, A.B.; Schwarz, K.U.; Tayebi, K.; et al. Environment and harvest time affects the combustion qualities of miscanthus genotypes. Agron. J. 2003, 95, 1274–1280. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Stampfl, P.F.; Jones, M.B. Miscanthus biomass production for energy in Europe and its potential contribution to decreasing fossil fuel carbon emissions. Glob. Chang. Biol. 2004, 10, 509–518. [Google Scholar] [CrossRef]
- Schulz, B. Average annual minimum temperatures for Europe. In Flora der Gehölze: Bestimmung, Eigenschaften, Verwendung; Roloff, A., Bärtels, A., Eds.; Ulmer Eugen Verlag: Stuttgart, Germany, 2008; p. 1. [Google Scholar]
- Skogsdata 2013; SLU: Umeå, Sweden, 2013; p. 157.
- Troedsson, T.; Wiberg, M. Sveriges Jordmåner (Soil Map of Sweden); Sveriges Lantbruksuniversitetet, Institutionen för Skoglig Marklära: Uppsala, Sweden, 1986; p. 2. [Google Scholar]
- Bergh, J.; Linder, S.; Lundmark, T.; Elfving, B. The effect of water and nutrient availability on the productivity of Norway spruce in northern and southern Sweden. For. Ecol. Manag. 1999, 119, 51–62. [Google Scholar] [CrossRef]
- Johansson, T. Biomass production of Norway spruce (Picea abies (L.) Karst.) growing on abandoned farmland. Silva Fenn. 1999, 33, 261–280. [Google Scholar] [CrossRef]
- Lässig, R.; Močalov, S.A. Frequency and characteristics of severe storms in the Urals and their influence on the development, structure and management of the boreal forests. For. Ecol. Manag. 2000, 135, 179–194. [Google Scholar] [CrossRef]
- Kramer, H.; Gussone, H.A.; Schober, R. Waldwachstumslehre: Ökologische und Anthropogene Einflüsse auf das Wachstum des Waldes, Seine Massen—Und Wertleistung und die Bestandessicherheit; mit 165 Tabellen; Paul Parey: Hamburg, Germany, 1988; p. 374. [Google Scholar]
- Kaltschmitt, M.; Hartmann, H.; Hofbauer, H. Energie aus Biomasse: Grundlagen, Techniken und Verfahren; Springer: Heidelberg, Germany, 2009; p. 1032. [Google Scholar]
- Schulze, E.D.; Vygodskaya, N.N.; Tchebakova, N.M.; Czimczik, C.I.; Kozlov, D.N.; Lloyd, J.; Mollicone, D.; Parfenova, E.; Sidorov, K.N.; Varlagin, A.V.; et al. The Eurosiberian Transect: An introduction to the experimental region. Tellus B 2002, 54, 421–428. [Google Scholar] [CrossRef]
- Xiao, C.-W.; Yuste, J.C.; Janssens, I.A.; Roskams, P.; Nachtergale, L.; Carrara, A.; Sanchez, B.Y.; Ceulemans, R. Above- and belowground biomass and net primary production in a 73-year-old Scots pine forest. Tree Physiol. 2003, 23, 505–516. [Google Scholar] [CrossRef]
- Bray, J.R. Root production and the estimation of net productivity. Can. J. Bot. 1963, 41, 65–72. [Google Scholar] [CrossRef]
- Becker, M. The role of climate on present and past vitality of silver fir forests in the Vosges mountains of northeastern France. Can. J. For. Res. 1989, 19, 1110–1117. [Google Scholar] [CrossRef]
- Macias, M.; Andreu, L.; Bosch, O.; Camarero, J.J.; Gutiérrez, E. Increasing aridity is enhancing Silver fir (Abies Alba Mill.) water stress in its south-western distribution limit. Clim. Chang. 2006, 79, 289–313. [Google Scholar] [CrossRef]
- Zavitkovski, J.; Strong, T.F. Biomass Production of 12-Year-Old Intensively Cultured Larix Eurolepis; North Central Forest Experiment Station, Forest Service, USDA: St. Paul, MN, USA, 1984; p. 3. [Google Scholar]
- Peeters, A.; Vanbellinghen, C.; Frame, J. Wild and Sown Grasses: Profiles of a Temperate Species Selection: Ecology, Biodiversity and Use; Food and Agriculture Organization of the United Nations/Wiley-Blackwell: Rome, Italy, 2004; p. 328. [Google Scholar]
- Fior, C.; Anfodillo, T.; Masotto, G.P.e.A. Biomass Partitioning and Structural Traits in Larix decidua Mill. and Pinus Cembra L. at Different Elevation; Treeline Ecology Research Unit, Dipartimento Territorio e Sistemi Agro Forestali, Università di Padova: Legnaro, Italy, 2005; p. 11. [Google Scholar]
- Bollschweiler, M.; Stoffel, M.; Schneuwly, D.M.; Bourqui, K. Traumatic resin ducts in Larix decidua stems impacted by debris flows. Tree Physiol. 2008, 28, 255–263. [Google Scholar] [CrossRef]
- Lingua, E.; Cherubini, P.; Motta, R.; Nola, P. Spatial structure along an altitudinal gradient in the Italian central Alps suggests competition and facilitation among coniferous species. J. Veg. Sci. 2008, 19, 425–436. [Google Scholar] [CrossRef]
- Grier, C.C.; Logan, R.S. Old-growth Pseudotsuga menziesii communities of a Western Oregon watershed: Biomass distribution and production budgets. Ecol. Monogr. 1977, 47, 373–400. [Google Scholar] [CrossRef]
- Kohnle, U. Douglasienanbau in Südwest-Deutschland: Waldbauliche Erfolgsfaktoren. In BFW-Praxisinformationen; Mauser, H., Ed.; Bundesforschungs- und Ausbildungszentrum für Wald, Naturgefahren und Landschaft (BFW): Wien, Austria, 2007; Volume 16, pp. 12–13. [Google Scholar]
- Hermann, R.K.; Lavender, D.P. Pseudotsuga menziesii (Mirb.) Franco. Douglas-Fir. Available online: http://www.na.fs.fed.us/pubs/silvics_manual/Volume_1/pseudotsuga/menziesii.htm (accessed on 17 May 2013).
- Zhang, Q.-B.; Hebda, R.J. Variation in radial growth patterns of Pseudotsuga menziesii on the central coast of British Columbia, Canada. Can. J. For. Res. 2004, 34, 1946–1954. [Google Scholar] [CrossRef]
- Syrjanen, K.; Kalliola, R.; Puolasmaa, A.; Mattsson, J. Landscape structure and forest dynamics in subcontinental russian european taiga. Ann. Zool. Fenn. 1994, 31, 19–34. [Google Scholar]
- Johansson, T. Biomass equations for determining fractions of common and grey alders growing on abandoned farmland and some practical implications. Biomass Bioenergy 2000, 18, 147–159. [Google Scholar] [CrossRef]
- Laureysens, I.; Deraedt, W.; Indeherberge, T.; Ceulemans, R. Population dynamics in a 6-year old coppice culture of poplar. I. Clonal differences in stool mortality, shoot dynamics and shoot diameter distribution in relation to biomass production. Biomass Bioenergy 2003, 24, 81–95. [Google Scholar] [CrossRef]
- Trnka, M.; Fialová, J.; Koutecký, V.; Fajman, M.; Žalud, Z.; Hejduk, S. Biomass production and survival rates of selected poplar clones grown under a short-rotation system on arable land. Plant Soil Environ. 2008, 54, 78–88. [Google Scholar]
- Diamantidis, N.D.; Koukios, E.G. Agricultural crops and residues as feedstocks for non-food products in Western Europe. Ind. Crops Prod. 2000, 11, 97–106. [Google Scholar] [CrossRef]
- Benetka, V.; Bartáková, I.; Mottl, J. Productivity of Populus nigra L. ssp. nigra under short-rotation culture in marginal areas. Biomass Bioenergy 2002, 23, 327–336. [Google Scholar]
- Helle, G.; Schleser, G.H. Beyond CO2-fixation by Rubisco—An interpretation of 13C/12C variations in tree rings from novel intra-seasonal studies on broad-leaf trees. Plant Cell Environ. 2004, 27, 367–380. [Google Scholar] [CrossRef]
- Barsoum, N. Relative contributions of sexual and asexual regeneration strategies in Populus nigra and Salix alba during the first years of establishment on a braided gravel bed river. Evol. Ecol. 2001, 15, 255–279. [Google Scholar] [CrossRef]
- Deiller, A.-F.; Walter, J.-M.N.; Trémolières, M. Effects of flood interruption on species richness, diversity and floristic composition of woody regeneration in the Upper Rhine alluvial hardwood forest. Regul. Rivers: Res. Manag. 2001, 17, 393–405. [Google Scholar] [CrossRef]
- Liesebach, M.; von Wuehlisch, G.; Muhs, H.J. Aspen for short-rotation coppice plantations on agricultural sites in Germany: Effects of spacing and rotation time on growth and biomass production of aspen progenies. For. Ecol. Manag. 1999, 121, 25–39. [Google Scholar] [CrossRef]
- Johansson, T. Biomass equations for determining fractions of European aspen growing on abandoned farmland and some practical implications. Biomass Bioenergy 1999, 17, 471–480. [Google Scholar] [CrossRef]
- Latva-Karjanmaa, T.; Suvanto, L.; Leinonen, K.; Rita, H. Emergence and survival of Populus tremula seedlings under varying moisture conditions. Can. J. For. Res. 2003, 33, 2081–2088. [Google Scholar] [CrossRef]
- Ellis, C.J.; Coppins, B.J. Reproductive strategy and the compositional dynamics of crustose lichen communities on aspen (Populus tremula L.) in Scotland. Lichenologist 2007, 39, 377–391. [Google Scholar] [CrossRef]
- Ilstedt, B.; Gullberg, U. Genetic variation in a 26-year old hybrid aspen trial in southern Sweden. Scand. J. For. Res. 1993, 8, 185–192. [Google Scholar] [CrossRef]
- Christersson, L. Wood production potential in poplar plantations in Sweden. Biomass Bioenergy 2010, 34, 1289–1299. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Evans, E.J.; Bilsborrow, P.E.; Wright, C.; Hewison, W.O.; Pilbeam, D.J. Yield of willow cultivars at different planting densities in a commercial short rotation coppice in the north of England. Biomass Bioenergy 2007, 31, 469–474. [Google Scholar] [CrossRef]
- Buček, A.; Maděra, P.; Úradníček, L. Czech Approach to Implementation of Ecological Network. J. Landsc. Ecol. 2012, 5, 14–28. [Google Scholar]
- Neirynck, J.; Mirtcheva, S.; Sioen, G.; Lust, N. Impact of Tilia platyphyllos Scop., Fraxinus excelsior L., Acer pseudoplatanus L., Quercus robur L. and Fagus sylvatica L. on earthworm biomass and physico-chemical properties of a loamy topsoil. For. Ecol. Manag. 2000, 133, 275–286. [Google Scholar] [CrossRef]
- Vitasse, Y.; Porté, A.; Kremer, A.; Michalet, R.; Delzon, S. Responses of canopy duration to temperature changes in four temperate tree species: Relative contributions of spring and autumn leaf phenology. Oecologia 2009, 161, 187–198. [Google Scholar] [CrossRef]
- Ritter, E.; Dalsgaard, L.; Einhorn, K.S. Light, temperature and soil moisture regimes following gap formation in a semi-natural beech-dominated forest in Denmark. For. Ecol. Manag. 2005, 206, 15–33. [Google Scholar] [CrossRef]
- Petritan, A.M.; von Lüpke, B.; Petritan, I.C. Effects of shade on growth and mortality of maple (Acer pseudoplatanus), ash (Fraxinus excelsior) and beech (Fagus sylvatica) saplings. Forestry 2007, 80, 397–412. [Google Scholar] [CrossRef]
- Radoglou, K.; Dobrowolska, D.; Spyroglou, G.; Nicolescu, V.N. A review on the ecology and silviculture of limes (Tilia cordata Mill., Tilia platyphyllos Scop. and Tilia tomentosa Moench.) in Europe. Bodenkultur-Wien and München 2009, 60, 9–20. [Google Scholar]
- Kirlum, F. Vegetationskundliche Untersuchungen im “Colbitzer Lindenwald”; FH-Eberswalde: Eberswalde, Germany, 1995; p. 35. [Google Scholar]
- López, F.; Pérez, A.; Zamudio, M.A.M.; de Alva, H.E.; García, J.C. Paulownia as raw material for solid biofuel and cellulose pulp. Biomass Bioenergy 2012, 45, 77–86. [Google Scholar] [CrossRef]
- Cokesa, V.; Koprivica, M.; Markovic, N.; Stajic, S. Production experiments of Paulownia in Vojvodina. In Proceedings of an International Scientific Conference Marking 75 Years of the Forest Research Institute of the Bulgarian Academy of Sciences, Sofia, Bulgaria, 1–5 October 2003; Rossnev, B., Kitanova, S., Alexandrov, A., Raev, I., Tsakov, H., Dimitrov, V., Grozeva, M., Petrova, R., Popov, G., Grigorov, G., Eds.; Forest Research Institute: Sofia, Bulgaria, 2004; Volume 1, pp. 248–255. [Google Scholar]
- Hodgson, E.M.; Nowakowski, D.J.; Shield, I.; Riche, A.; Bridgwater, A.V.; Clifton-Brown, J.C.; Donnison, I.S. Variation in miscanthus chemical composition and implications for conversion by pyrolysis and thermo-chemical bio-refining for fuels and chemicals. Bioresour. Technol. 2011, 102, 3411–3418. [Google Scholar] [CrossRef]
- Heaton, E.A.; Dohleman, F.G.; Long, S.P. Meeting US biofuel goals with less land: The potential of Miscanthus. Glob. Chang. Biol. 2008, 14, 2000–2014. [Google Scholar] [CrossRef]
- Greenhalf, C.E.; Nowakowski, D.J.; Bridgwater, A.V.; Titiloye, J.; Yates, N.; Riche, A.; Shield, I. Thermochemical characterisation of straws and high yielding perennial grasses. Ind. Crops Prod. 2012, 36, 449–459. [Google Scholar] [CrossRef]
- Finell, M. The Use of Reed Canary Grass (Phalaris arundinacea) as a Short Fibre Raw Material for the Pulp and Paper Industry. Ph.D. Thesis, Swedish University of Agricultural Sciences, Umeå, Sweden, 2003; p. 53. [Google Scholar]
- Casler, M.; Cherney, J.; Brummer, E.C. Biomass yield of naturalized populations and cultivars of reed canary grass. Bioenergy Res. 2009, 2, 165–173. [Google Scholar] [CrossRef]
- Paulownia-welt.de Paulownia. Available online: http://www.paulownia-welt.de/paulownia/index.html (accessed on 12 April 2013).
- Institute_LLC Paulownia growing zones. Available online: http://www.worldpaulownia.com/html/zones.html (accessed on 12 April 2013).
- Bijak, S. Tree-ring chronology of Silver fir and its dependence on climate of the Kaszubskie lakeland (northern Poland). Geochromonetria 2010, 35, 91–94. [Google Scholar]
- Peguero-Pina, J.J.; Camarero, J.J.; Abadía, A.; Martín, E.; González-Cascón, R.; Morales, F.; Gil-Pelegrín, E. Physiological performance of silver-fir (Abies alba Mill.) populations under contrasting climates near the south-western distribution limit of the species. Flora Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 226–236. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trischler, J.; Sandberg, D.; Thörnqvist, T. Estimating the Annual Above-Ground Biomass Production of Various Species on Sites in Sweden on the Basis of Individual Climate and Productivity Values. Forests 2014, 5, 2521-2541. https://0-doi-org.brum.beds.ac.uk/10.3390/f5102521
Trischler J, Sandberg D, Thörnqvist T. Estimating the Annual Above-Ground Biomass Production of Various Species on Sites in Sweden on the Basis of Individual Climate and Productivity Values. Forests. 2014; 5(10):2521-2541. https://0-doi-org.brum.beds.ac.uk/10.3390/f5102521
Chicago/Turabian StyleTrischler, Johann, Dick Sandberg, and Thomas Thörnqvist. 2014. "Estimating the Annual Above-Ground Biomass Production of Various Species on Sites in Sweden on the Basis of Individual Climate and Productivity Values" Forests 5, no. 10: 2521-2541. https://0-doi-org.brum.beds.ac.uk/10.3390/f5102521