Effects of Site Preparation Methods on the Establishment and Natural-Regeneration Traits of Scots Pines (Pinus sylvestris L.) in Northeastern Poland

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
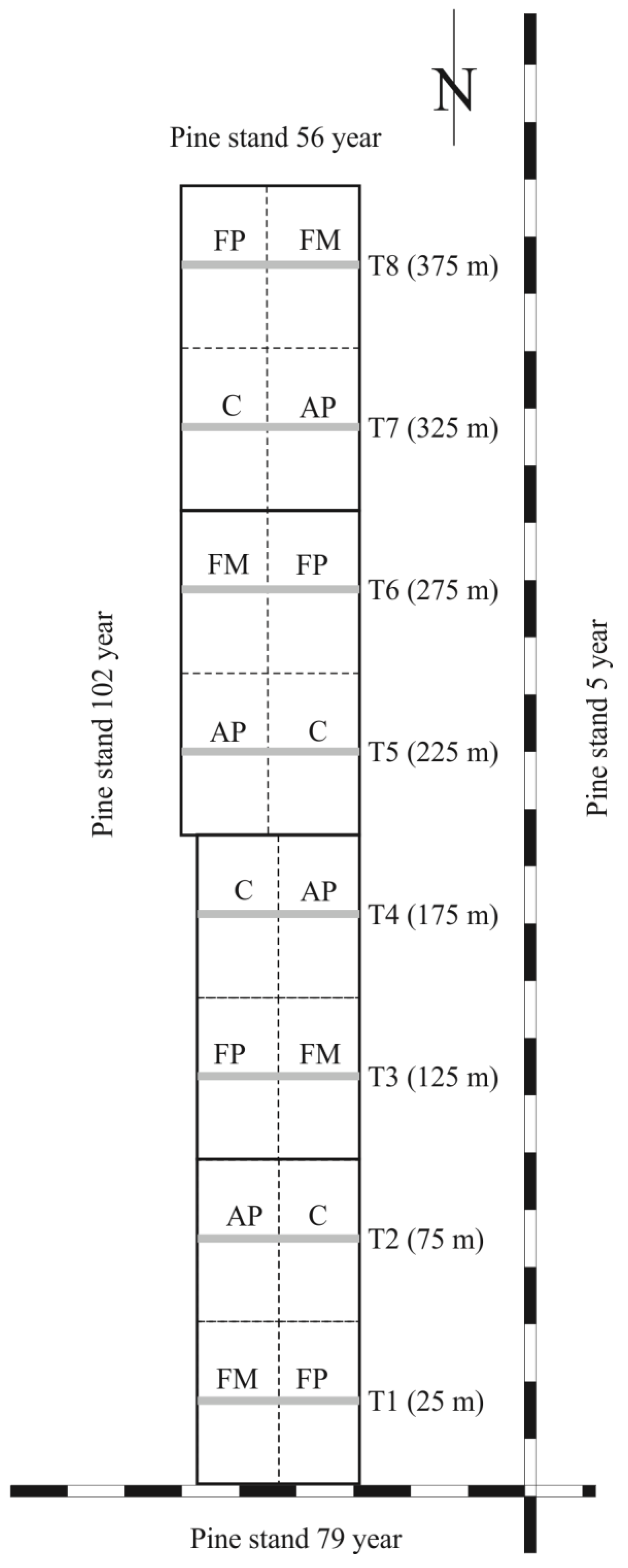
2.1. Study Area
2.2. Weather Conditions
2.3. Treatments
2.4. Measurements
2.5. Soil Analysis
2.6. Statistical Analysis
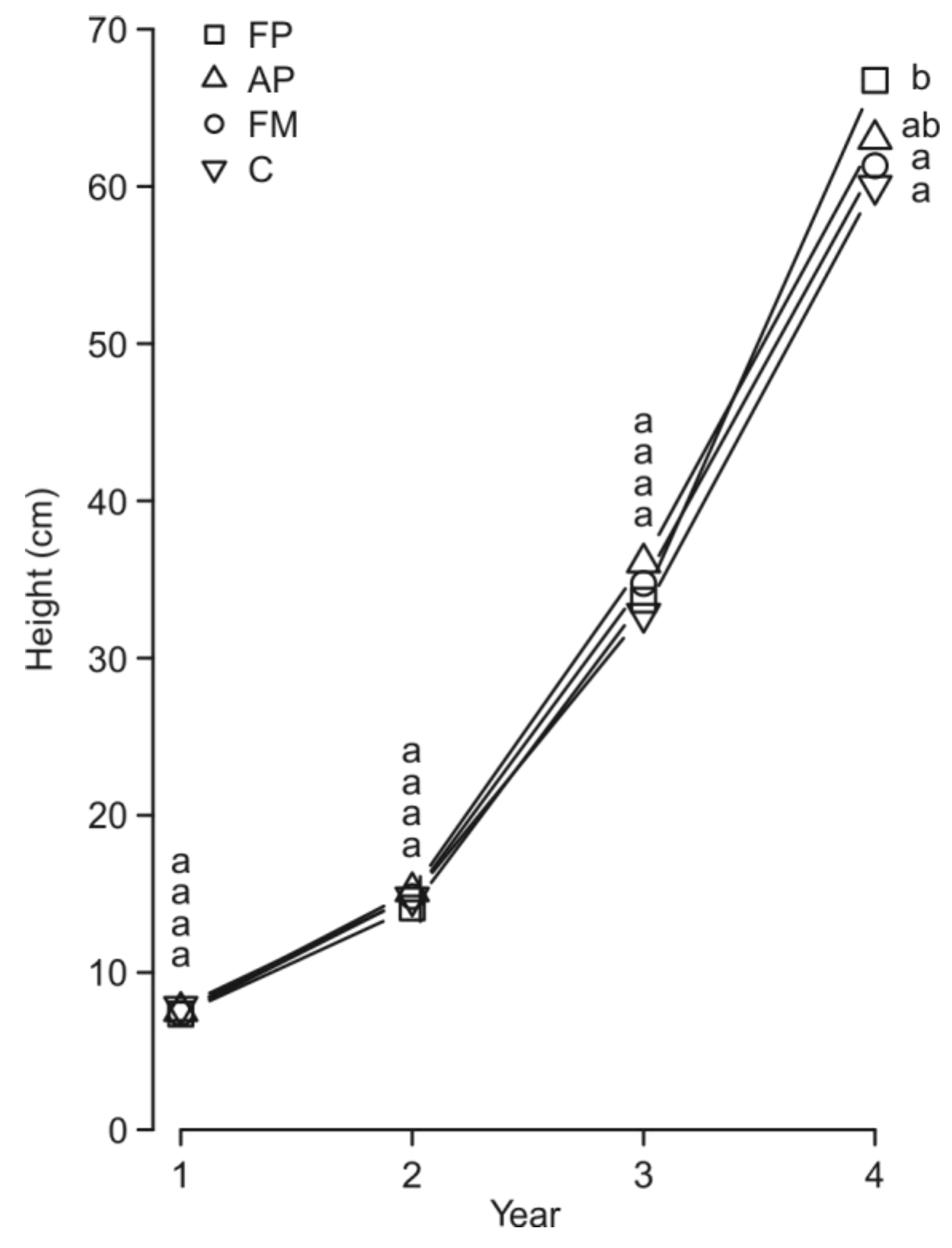
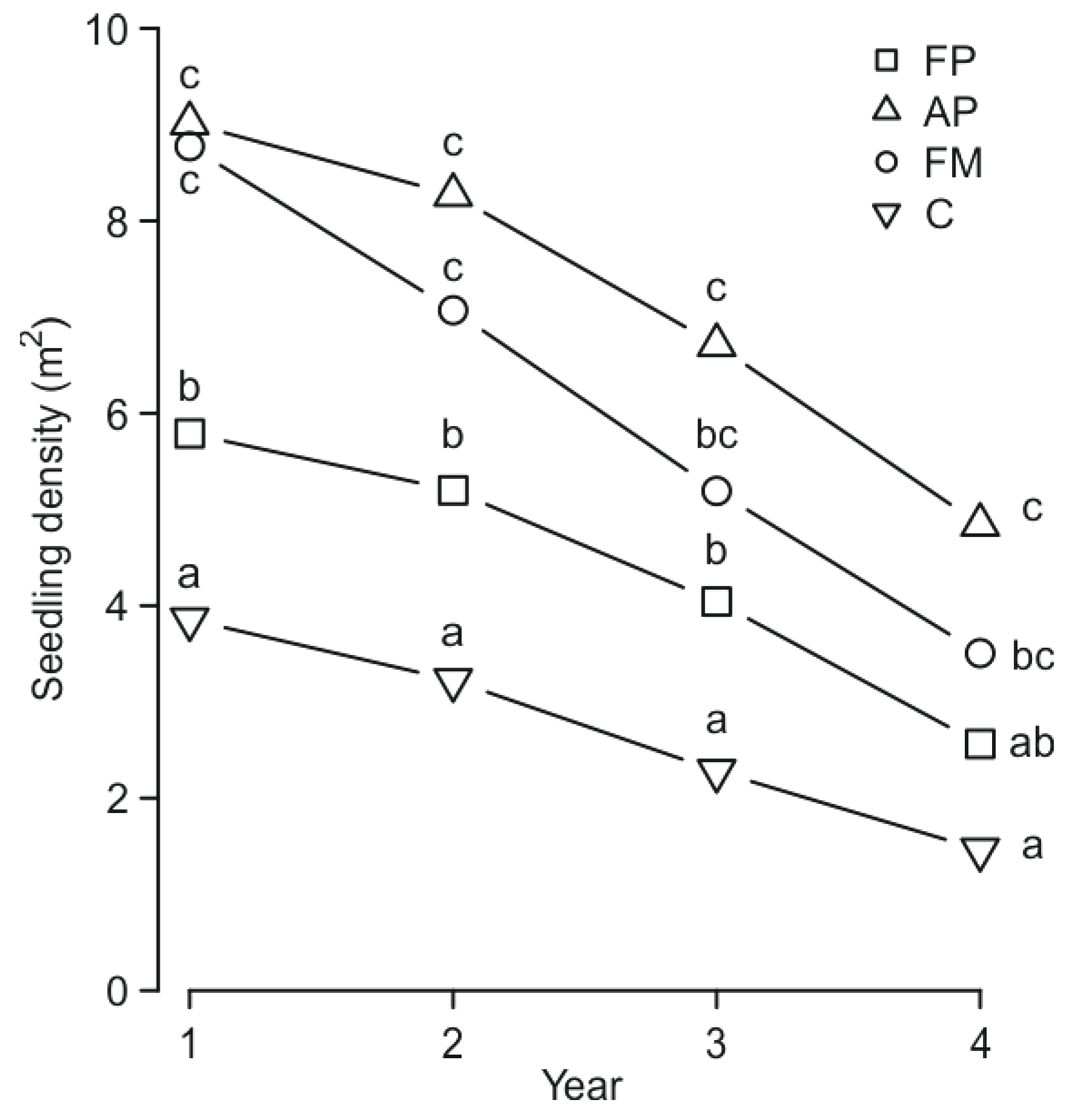
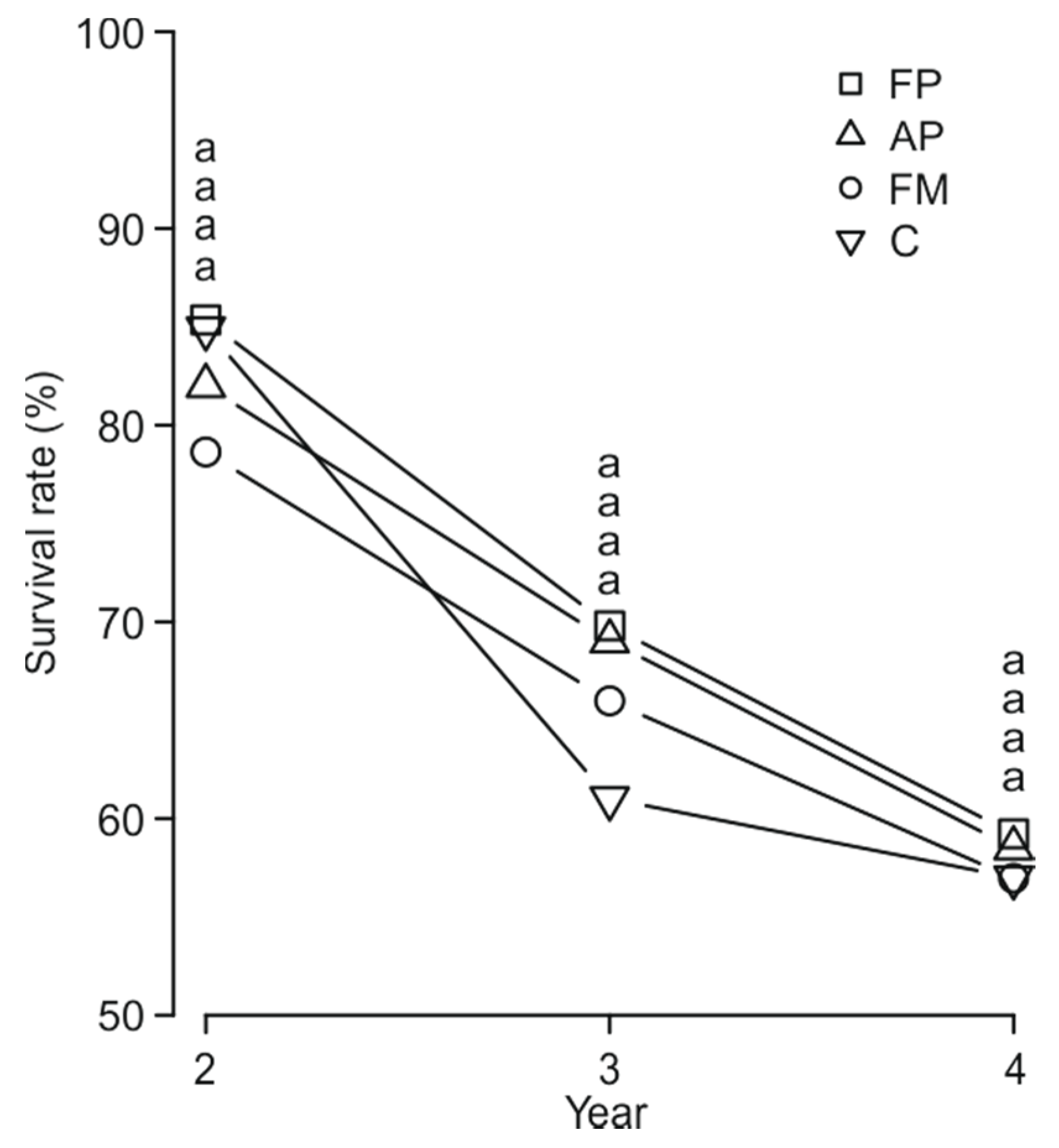
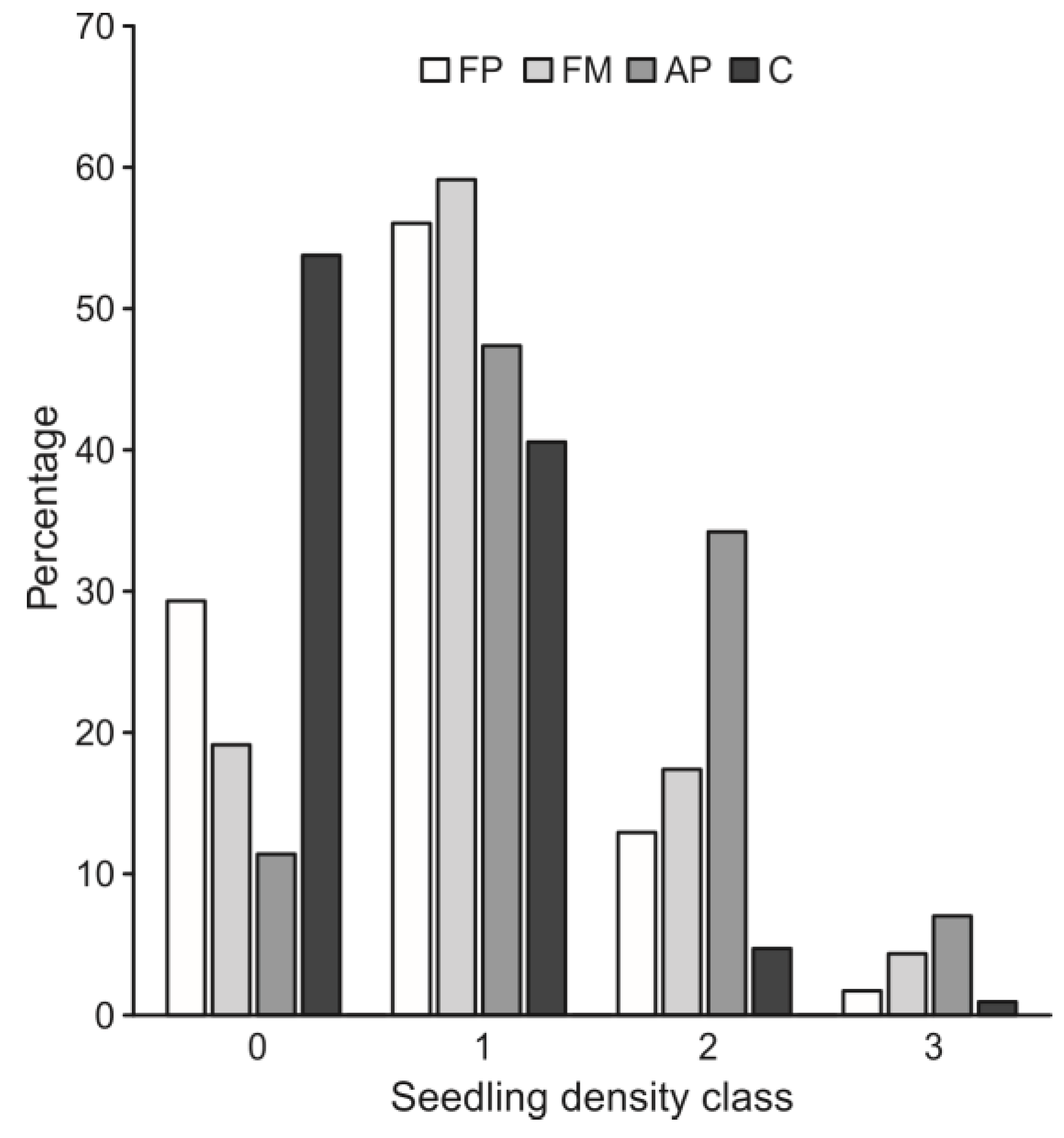
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brus, D.J.; Hengeveld, G.M.; Walvoort, D.J.J.; Goedhart, P.W.; Heidema, A.H.; Nabuurs, G.J.; Gunia, K. Statistical mapping of tree species over Europe. Eur. J. For. Res. 2011, 131, 145–157. [Google Scholar] [CrossRef]
- Forestry 2017 Statistical Information and Elaborations. Available online: http://stat.gov.pl/obszary-tematyczne/rolnictwo-lesnictwo/lesnictwo/lesnictwo-2017,1,13.html (accessed on 18 August 2018).
- Principles of Silviculture in Poland. Available online: http://www.lasy.gov.pl/pl/pro/publikacje/copy_of_gospodarkalesna/hodowla/zasady-hodowli-lasu-dokument-w-opracowaniuview (accessed on 18 August 2018).
- Tomczyk, S. Odnowienie Naturalne: Sosna; Biblioteczka leśniczego zeszyt 29; Wydawnictwo Świat: Warszawa, Poland, 1993; Volume 29, p. 23. [Google Scholar]
- Andrzejczyk, T.; Żybura, H. Sosna Zwyczajna—Odnawianie Naturalne i Alternatywne Metody Hodowli; PWRiL: Warsaw, Poland, 2012; p. 252. [Google Scholar]
- Kerr, G. Natural regeneration of Corsican pine (Pinus nigra subsp. laricio) in Great Britain. Forestry 2000, 73, 479–488. [Google Scholar]
- Oleskog, G.; Sahlén, K. Effect of seedbed substrate on moisture conditions and germination of Scots pine (Pinus sylvestris) seeds in a mixed conifer stand. New For. 2000, 20, 119–133. [Google Scholar] [CrossRef]
- Puhlick, J.J.; Laughlin, D.C.; Moor, M.M. Factors influencing ponderosa pine regeneration in the southwestern USA. For. Ecol. Manag. 2012, 264, 10–19. [Google Scholar] [CrossRef]
- Hille, M.; den Ouden, J. Improved recruitment and early growth of Scots pine (Pinus sylvestris L.) seedlings after fire and soil scarification. Eur. J. For. Res. 2004, 123, 213–218. [Google Scholar] [CrossRef]
- Caccia, F.D.; Ballaré, C.L. Effects of tree cover, understory vegetation, and litter on regeneration of Douglas-fir (Pseudotsuga manziessii) in southwestern Argentina. Can. J. For. Res. 1998, 28, 683–692. [Google Scholar] [CrossRef]
- Ibáñez, I.; Schupp, E.W. Effects of litter, soil surface conditions, and microhabitat on Cerocarpus ledifolius Nutt. Seedling emergence and establishment. J. Arid Environ. 2002, 52, 209–221. [Google Scholar] [CrossRef]
- Oleskog, G.; Sahlén, K. Effect of seedbed substrate on moisture conditions and germination of Pinus sylvestris (L.) seeds in clear-cut. Scand. J. For. Res. 2000, 15, 225–236. [Google Scholar] [CrossRef]
- Löf, M.; Dey, D.C.; Navarro, R.M.; Jacobs, D.F. Mechanical site preparation for forest restoration. New For. 2012, 43, 825–848. [Google Scholar] [CrossRef]
- Nilsson, U.; Örlander, G. Vegetation management on grass—Dominated clearcuts planted with Norway spruce in southern Sweden. Can. J. For. Res. 1999, 29, 1015–1026. [Google Scholar] [CrossRef]
- Steijlen, I.; Nilsson, M.-C.; Zackrisson, O. Seed regeneration of Scots pine in boreal forest stand dominated by lichen and feather moss. Can. J. For. Res. 1995, 25, 713–723. [Google Scholar] [CrossRef]
- Jäderlund, A.; Norberg, G.; Zackrisson, O.; Dahlberg, A.; Teketay, D.; Dolling, A.; Nilsson, M.C. Control of bilberry vegetation by steam treatment—Effects on seeded Scots pine and associated mycorrhizal fungi. For. Ecol. Manag. 1998, 108, 275–285. [Google Scholar] [CrossRef]
- Petersson, M.; Örlander, G. Effectiveness of combinations of shelterwood, scarification, and feeding barriers to reduce pine weevil damage. Can. J. For. Res. 2003, 33, 64–73. [Google Scholar] [CrossRef]
- Birkedal, M.; Löf, M.; Olsson, G.E.; Bergsten, U. Effects of granivorous rodents on direct seeding of oak and beech in relation to site preparation and sowing date. For. Ecol. Manag. 2010, 259, 2382–2389. [Google Scholar] [CrossRef]
- Lundmark-Thelin, A.; Johansson, M.B. Influence of mechanical site preparation on decomposition and nutrient dynamics of Norway spruce (Picea abies (L.) Karst.) needle litter and slash needles. For. Ecol. Manag. 1997, 97, 101–110. [Google Scholar] [CrossRef]
- Piirainen, S.; Finér, L.; Mannerkoski, H.; Starr, M. Carbon, nitrogen and phosphorus leaching after mechanical site preparation at a boreal forest clear-cut area. For. Ecol. Manag. 2007, 243, 10–18. [Google Scholar] [CrossRef]
- Piirainen, S.; Finér, L.; Mannerkoski, H.; Starr, M. Leaching of cations and sulphate after mechanical site preparation at a boreal forest clear-cut area. Geoderma 2009, 149, 386–392. [Google Scholar] [CrossRef]
- Alcázar, J.; Rothwell, L.R.; Woodard, M.P. Soil disturbance and the potential for erosion after mechanical site preparation. North. J. Appl. For. 2002, 19, 5–13. [Google Scholar]
- Archibold, O.W.; Acton, C.; Ripley, E.A. Effect of site preparation on soil properties and vegetation cover, and the growth and survival of white spruce (Picea glauca) seedlings, in Saskatchewan. For. Ecol. Manag. 2000, 131, 127–141. [Google Scholar] [CrossRef]
- Block, M.D.; van Rees, K.C.J. Mechanical site preparation impacts on soil properties and vegetation communities in the Northwest Territories. Can. J. For. Res. 2002, 32, 1381–1392. [Google Scholar]
- MacKenzie, M.D.; Schmidt, M.G.; Bedford, L. Soil microclimate and nitrogen availability 10 years after mechanical site preparation in northern British Columbia. Can. J. For. Res. 2005, 35, 1854–1866. [Google Scholar] [CrossRef]
- Heiskanen, J.; Mäkitalo, K.; Hyvönen, J. Long-term influence of site preparation on water-retention characteristics of forest soil in Finnish Lapland. For. Ecol. Manag. 2007, 241, 127–133. [Google Scholar] [CrossRef]
- Munson, A.D.; Timmer, V.R. Soil nitrogen dynamics and nutrition of pine following silvicultural treatments in boreal and Great lakes—St. Lawrence plantations. For. Ecol. Manag. 1995, 76, 169–179. [Google Scholar] [CrossRef]
- Mäkitalo, K. Effect of site preparation and reforestation method on survival and height growth of Scots pine. Scand. J. For. Res. 1999, 14, 512–525. [Google Scholar] [CrossRef]
- Wallertz, K.; Malmqvist, C. The effect of mechanical site preparation methods on the establishment of Norway spruce (Picea abies (L.) Karst.) and Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) in southern Sweden. Forestry 2013, 86, 71–78. [Google Scholar] [CrossRef]
- Sutton, R.F. Mounding site preparation: A review of European and North American experience. New For. 1993, 7, 151–192. [Google Scholar] [CrossRef]
- Bedford, L.; Sutton, R.F. Site preparation for establishing lodgepole pine in the sub-boreal spruce zone of interior British Columbia: The Bednesti trial, 10-year results. For. Ecol. Manag. 2000, 126, 227–238. [Google Scholar] [CrossRef]
- Neugebauer, Z. Poradnik dla Operatorów Maszyn Leśnych Agregowanych na Ciągnikach; Dyrekcja Generalna Lasów Państwowych: Warsaw-Bedoń, Poland, 2008; p. 249. [Google Scholar]
- Prévost, M. Effects of scarification on seedbed coverage and natural regeneration after a group seed-tree cutting in a black spruce (Picea mariana) stand. For. Ecol. Manag. 1997, 94, 219–231. [Google Scholar] [CrossRef]
- Mattsson, S.; Bergsten, U. Pinus contorta growth in northern Sweden as affected by soil scarification. New For. 2003, 26, 217–231. [Google Scholar] [CrossRef]
- Nordborg, F.; Nilsson, U. Growth, damage and net nitrogen uptake in Picea abies (L.) Karst. seedlings, effects of site preparation and fertilization. Ann. For. Sci. 2003, 60, 657–666. [Google Scholar] [CrossRef]
- Bilodeau-Gauthier, S.; Paré, D.; Messier, C.; Bélanger, N. Juvenile growth of hybrid poplars on acid boreal soil determined by environmental effects of soil preparation, vegetation control, and fertilization. For. Ecol. Manag. 2011, 261, 620–629. [Google Scholar] [CrossRef] [Green Version]
- Boateng, J.O.; Heineman, J.L.; McClarnon, J.; Bedford, L. Twenty-year responses of white spruce to mechanical site preparation and early chemical release in the boreal region of northeastern British Columbia. Can. J. For. Res. 2006, 36, 2386–2399. [Google Scholar] [CrossRef]
- Aleksandrowicz-Trzcińska, M.; Drozdowski, S.; Brzeziecki, B.; Rutkowska, P.; Jabłońska, B. Effect of different methods of site preparation on natural regeneration of Pinus sylvestris in Eastern Poland. Dendrobiology 2014, 71, 73–81. [Google Scholar] [CrossRef]
- Bureau for Forest Management and Geodesy in Olsztyn. Management Plan for Spychowo Forest District for Years 2013–2022: Stand description data, An internal document for Spychowo Forest District elaborated by the Bureau for Forest Management and Geodesy in Olsztyn. 2013. [Google Scholar]
- De Martonne, E. Une nouvelle fanction climatologique: L’indice d’aridité. Météorologie 1926, 2, 449–458. [Google Scholar]
- Hounam, C.E.; Burgos, J.J.; Kalik, M.S.; Palmer, W.C.; Rodda, J. Drought and Agriculture—Report of the Commission for Agricultural Meteorology Working Group on Assessment of Drought; Technical Note No. 138; World Meteorological Organization: Geneva, Switzerland, 1975; p. 127. [Google Scholar]
- Andrzejczyk, T.; Drozdowski, S. Rozwój naturalnego odnowienia sosny zwyczajnej na powierzchni przygotowanej pługiem dwuodkładnicowym. Sylwan 2003, 5, 28–35. [Google Scholar]
- Karlsson, M.; Nilsson, U. The effects of scarification and shelterwood treatments on naturally regenerated seedlings in southern Sweden. For. Ecol. Manag. 2005, 205, 183–197. [Google Scholar] [CrossRef]
- Karlsson, C.; Örlander, G. Soil scarification shortly before a rich seed fall improves seedling establishment in seed tree stands of Pinus sylvestris. Scand. J. For. Res. 2000, 15, 256–266. [Google Scholar] [CrossRef]
- Béland, M.; Agestam, E.; Ekö, P.M.; Gemmel, P.; Nilsson, U. Scarification and seedfall affects natural regeneration of Scots pine under two shelterwood densities and clear-cut in southern Sweden. Scand. J. For. Res. 2000, 15, 247–255. [Google Scholar] [CrossRef]
- Agestam, E.; Ekö, P.M.; Nilsson, U.; Welander, N.T. The effects of shelterwood density and site preparation on natural regeneration of Fagus sylvatica in southern Sweden. For. Ecol. Manag. 2003, 176, 61–73. [Google Scholar] [CrossRef]
- De Chantal, M.; Leinonen, K.; Ilvesniemi, H.; Westman, C.J. Effects of site preparation on soil properties and on morphology of Pinus silvestris and Picea abies seedlings sown at different dates. New For. 2004, 27, 159–173. [Google Scholar] [CrossRef]
- Sewerniak, P.; Gonet, S.S.; Quaium, M. Wpływ przygotowania gleby frezem leśnym na wzrost sadzonek sosny zwyczajnej w warunkach ubogich siedlisk Puszczy Bydgoskiej. Sylwan 2012, 156, 871–880. [Google Scholar]
- Bergsten, U. Temperature tolerance of invigorated seeds of Pinus sylvestris L., and Picea abies (L.) Karst. using TTGP-test. For. Suppl. 1989, 62, 107–115. [Google Scholar]
- Karlsson, M.; Nilsson, U.; Örlander, G. Natural regeneration in clear-cuts: Effects of scarification, slash removal and clear-cut age. Scand. J. For. Res. 2002, 17, 131–138. [Google Scholar] [CrossRef]
- Örlander, G.; Egnell, G.; Albrektson, A. Long-term effects of site preparation on growth in Scots pine. For. Ecol. Manag. 1996, 86, 27–37. [Google Scholar] [CrossRef]
- Andrzejczyk, T.; Augustyniak, G. Wpływ przygotowania gleby na wzrost sosny zwyczajnej w pierwszych latach uprawy. Sylwan 2007, 8, 3–8. [Google Scholar]
- Sewerniak, P.; Stelter, P. Wpływ sposobu przygotowania gleby na dynamikę jej temperatury na wydmach Kotliny Toruńskiej. Sylwan 2016, 160, 923–932. [Google Scholar]
- Collet, C.; Moguedec, G. Individual seedling mortality as a function in naturally regenerated beech seedlings. Forestry 2007, 80, 359–370. [Google Scholar] [CrossRef]
- Akashi, N. Dispersion pattern and mortality of seeds and seedlings of Fagus crenata Blume in a cool temperate forest in western Japan. Ecol. Res. 1997, 12, 159–165. [Google Scholar] [CrossRef]
- Willoughby, I.; Jinks, R.L.; Kerr, G.; Gosling, P.G. Factors affecting the success of direct seeding for lowland afforestation in the UK. Forestry 2004, 77, 467–482. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Garcia, E.; Grater, G.; Bravo, F. Climatic variability and other site factors influence on natural regeneration of Pinus pinaster Ait. in Mediterranean forests. Ann. For. Sci. 2011, 68, 811–823. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Temperature (°C) | Precipitation (mm) | Aridity Index | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2014 | 2015 | 2016 | 2017 | 2014 | 2015 | 2016 | 2017 | 2014 | 2015 | 2016 | 2017 | |
| January | −4.3 | −0.5 | −5.2 | −0.6 | 57.4 | 62.6 | 39.8 | 33.6 | 119.74 | 78.99 | 98.42 | 42.85 |
| February | −0.7 | 0.7 | 1.8 | 0.4 | 30.9 | 51.9 | 146.4 | 68.0 | 39.78 | 66.69 | 148.87 | 78.60 |
| March | 4.4 | 4.0 | 2.1 | 4.2 | 5.1 | 94.5 | 30.0 | 57.1 | 4.21 | 81.24 | 29.82 | 48.31 |
| April | 8.2 | 6.6 | 7.6 | 6.8 | 60.3 | 69.1 | 62.3 | 98.3 | 39.72 | 49.96 | 42.57 | 70.23 |
| May | 12.8 | 11.5 | 13.0 | 13.5 | 42.9 | 60.7 | 82.6 | 31.1 | 22.61 | 33.91 | 43.18 | 15.87 |
| June | 14.4 | 15.4 | 17.5 | 17.9 | 65.9 | 37.6 | 33.4 | 28.6 | 32.49 | 17.76 | 14.61 | 12.32 |
| July | 19.6 | 17.7 | 18.0 | 18.2 | 180.2 | 99.4 | 81.3 | 166.2 | 73.10 | 43.01 | 34.90 | 70.83 |
| August | 17.0 | 19.8 | 16.8 | 18.6 | 126.8 | 6.9 | 123.8 | 93.7 | 56.38 | 2.76 | 55.44 | 39.30 |
| September | 13.5 | 14.1 | 13.9 | 14.1 | 43.7 | 77.2 | 18.0 | 228.3 | 22.31 | 38.46 | 9.05 | 113.77 |
| October | 8.0 | 6.1 | 6.6 | 8.8 | 22.8 | 12.5 | 166.2 | 171.0 | 15.18 | 9.31 | 119.89 | 109.19 |
| November | 2.5 | 4.1 | 2.4 | 4.3 | 9.5 | 138.1 | 103.1 | 55.0 | 9.06 | 117.51 | 99.80 | 46.06 |
| December | −1.2 | 3.2 | 1.2 | 1.7 | 33.2 | 115.7 | 94.8 | 106.8 | 45.10 | 105.30 | 101.86 | 109.09 |
| Variable | Source of Variation | Wald F Statistic | Degree of Freedom | p Value |
|---|---|---|---|---|
| Seedling height | Seedling density | 4.1049 | 1 | 0.04276 * |
| MSP method (M) | 3.0034 | 3 | 0.3911 | |
| Seedling age (A) | 4132.959 | 3 | ˂0.0001 *** | |
| M × A | 15.2052 | 9 | 0.04545 * | |
| Seedling density | MSP method (M) | 237.867 | 3 | ˂0.0001 *** |
| Seedling age (A) | 169.495 | 3 | ˂0.0001 *** | |
| M × A | 14.299 | 9 | 0.1121 | |
| Seedling survival | Seedling density | 32.8115 | 1 | 0.0001 *** |
| MSP method (M) | 10.7168 | 3 | 0.01336 * | |
| Seedling age (A) | 229.5242 | 2 | ˂0.0001 *** | |
| M × A | 2.4329 | 6 | 0.8759 |
| Feature | Forest Plough | Active Plough | Forest Mill | Without MSP | p Value | |||
|---|---|---|---|---|---|---|---|---|
| Ridge | Furrow | Ridge | Furrow | Strip | Outside the Strip | |||
| Total N (g kg−1) | 1.64 (0.104)d | 0.46 (0.034)a | 2.23 (0.288)e | 0.54 (0.042)ab | 1.07 (0.081)c | 0.96 (0.092)bc | 0.96 (0.044)bc | ˂0.0001 |
| Total C (g kg−1) | 31.6 (1.787)c | 9.3 (0.710)a | 46.1 (6.603)d | 10.9 (0.803)a | 22.4 (1.41)b | 20.2 (1.922)b | 20.7 (1.009)b | ˂0.0001 |
| C/N | 19.4 (0.368)a | 20.3 (0.710)a | 20.5 (0.438)a | 20.5 (0.711)a | 21.0 (0.383)a | 21.1 (0.553)a | 21.5 (0.215)a | 0.1136 |
| P2O5 (mg 100 g−1) | 6.37 (0.522)a | 8.59 (0.434)b | 6.75 (0.423)ab | 7.92 (0.348)ab | 8.27 (0.508)ab | 6.99 (0.597)ab | 7.05 (0.546)ab | 0.0149 |
| Ca2+ (mg 100 g−1) | 11.31 (1.776)b | 1.57 (0.128)a | 18.25 (3.079)c | 1.97 (0.328)a | 6.32 (1.095)ab | 5.96 (0.610)ab | 5.51 (0.569)ab | ˂0.0001 |
| K+ (mg 100 g−1) | 5.38 (0.533)b | 1.16 (0.101)a | 7.32 (1.094)b | 1.28 (0.087)a | 2.79 (0.271)a | 2.68 (0.202)a | 2.50 (0.168)a | ˂0.0001 |
| Mg2+ (mg 100 g−1) | 1.681 (0.216)b | 0.235 (0.016)a | 2.757 (0.431)c | 0.274 (0.031)a | 0.957 (0.148)ab | 0.914 (0.088)ab | 0.820 (0.073)a | ˂0.0001 |
| Na+ (mg 100 g−1) | 0.272 (0.035)a | 0.170 (0.031)a | 0.242 (0.034)a | 0.131 (0.046)a | 0.223 (0.028)a | 0.219 (0.028)a | 0.145 (0.029)a | 0.0564 |
| Bd (g cm−3) | 0.74 (0.084)a | 1.36 (0.023)c | 0.85 (0.111)ab | 1.29 (0.036)c | 0.92 (0.058)ab | 1.12 (0.044)bc | 1.10 (0.064)bc | ˂0.0001 |
| Am (g 100 cm−3) | 17.2 (3.366)a | 12.6 (1.238)a | 16.1 (2.357)a | 13.7 (1.399)a | 15.6 (2.394)a | 19.4 (3.414)a | 20.6 (3.025)a | 0.3010 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleksandrowicz-Trzcińska, M.; Drozdowski, S.; Studnicki, M.; Żybura, H. Effects of Site Preparation Methods on the Establishment and Natural-Regeneration Traits of Scots Pines (Pinus sylvestris L.) in Northeastern Poland. Forests 2018, 9, 717. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110717
Aleksandrowicz-Trzcińska M, Drozdowski S, Studnicki M, Żybura H. Effects of Site Preparation Methods on the Establishment and Natural-Regeneration Traits of Scots Pines (Pinus sylvestris L.) in Northeastern Poland. Forests. 2018; 9(11):717. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110717
Chicago/Turabian StyleAleksandrowicz-Trzcińska, Marta, Stanisław Drozdowski, Marcin Studnicki, and Henryk Żybura. 2018. "Effects of Site Preparation Methods on the Establishment and Natural-Regeneration Traits of Scots Pines (Pinus sylvestris L.) in Northeastern Poland" Forests 9, no. 11: 717. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110717