Succession of Mite Assemblages (Acari, Mesostigmata) during Decomposition of Tree Leaves in Forest Stands Growing on Reclaimed Post-Mining Spoil Heap and Adjacent Forest Habitats

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Site Description and Experimental Design
2.2. Soil Mite Investigation
2.3. Statistical Analysis
3. Results
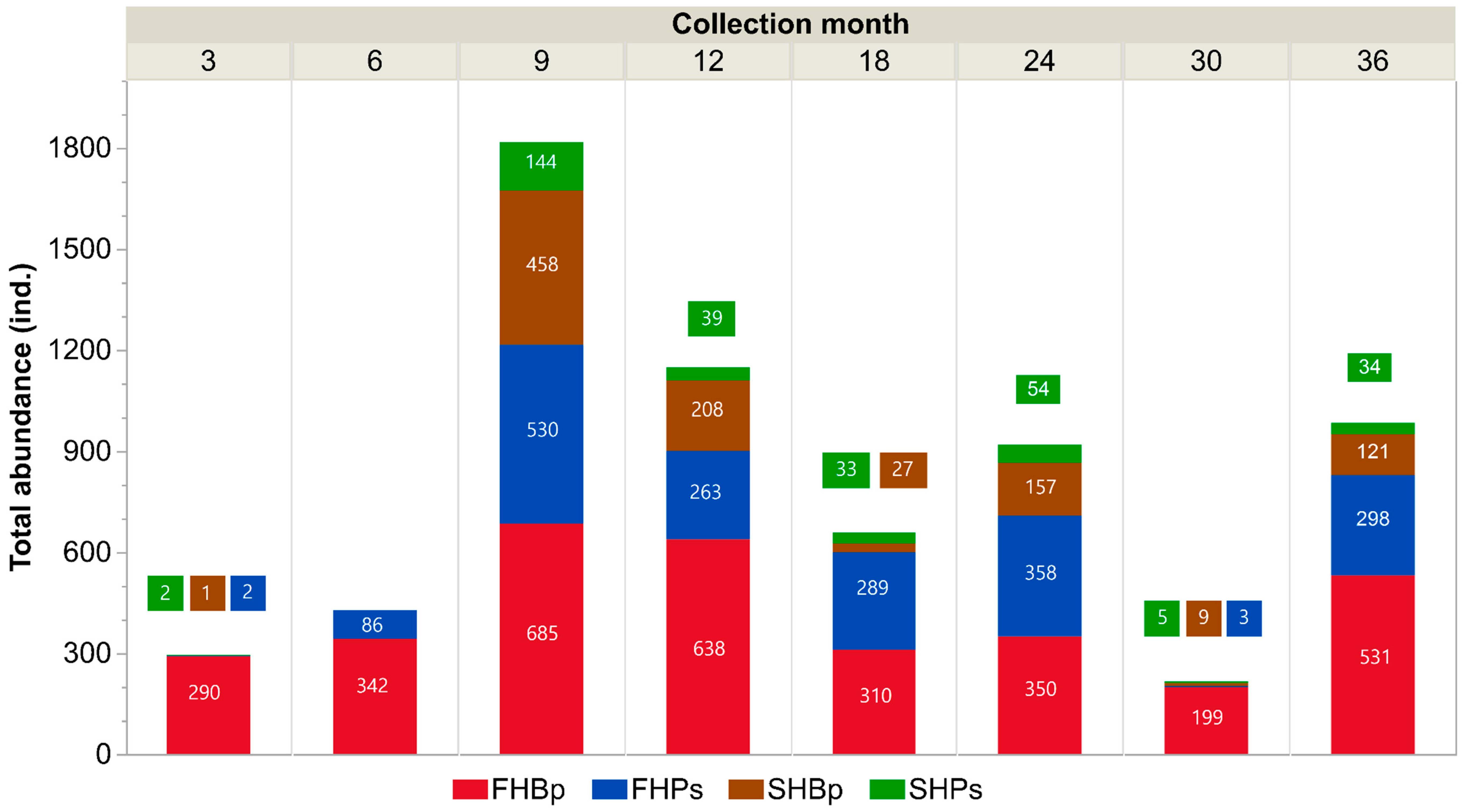
3.1. Mite Abundance
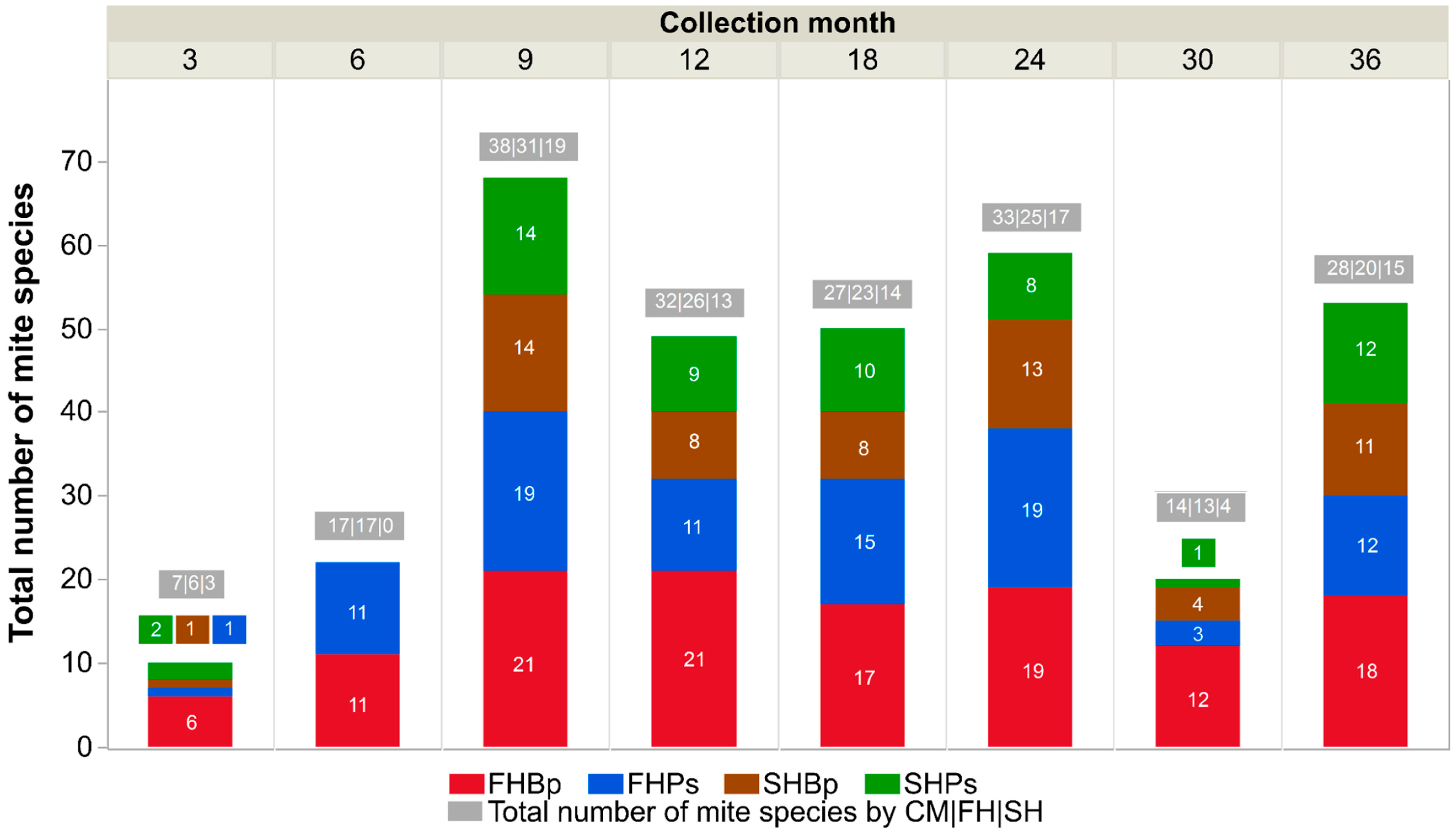
3.2. Mite Species Richness
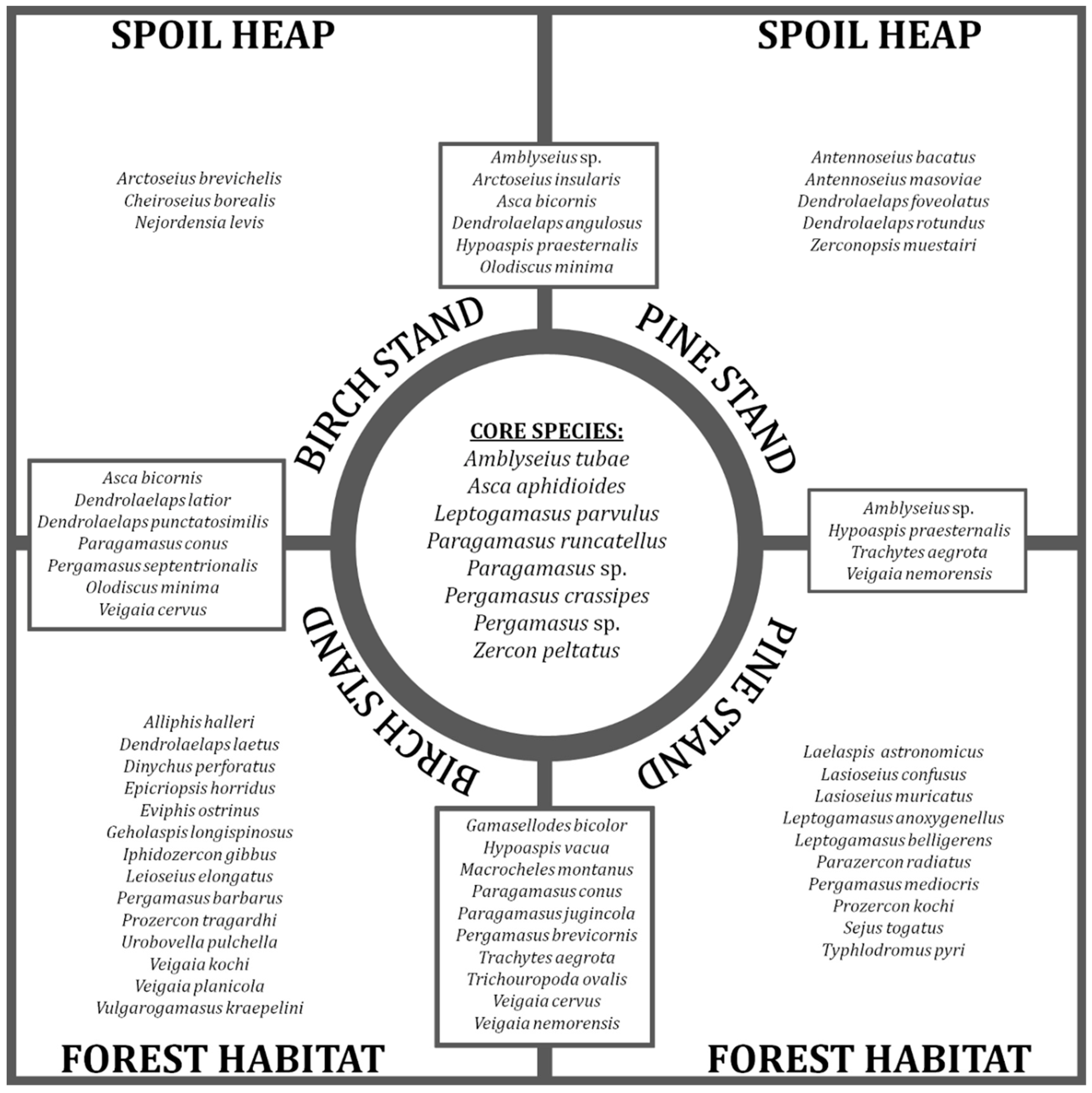
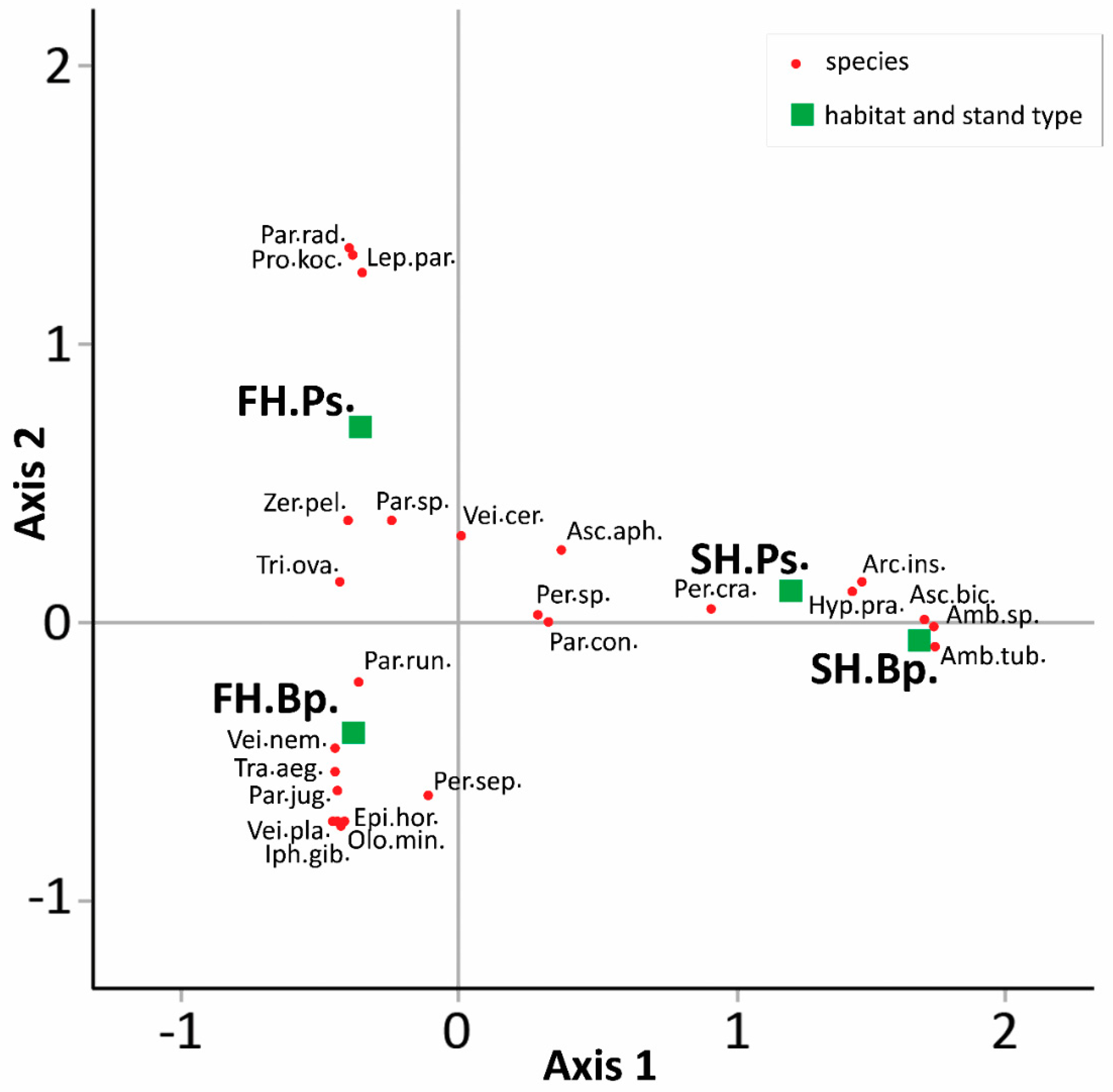
3.3. Mite Assemblages
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot No. | Coordinates | Plot Area (m2) | Stand Type | Habitat Type | Soil Texture | Humus Type | pH H2O (O Layer) | Stand Age (years) | Average DBH (cm) * ± SE | Average H (m) * ± SE | BA (m2 ha−1) *,** | Stocking Density (ind. ha−1) *,*** |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 | 51.2207, 19.4339 | 604.5 | Bp | SH | CL | FPMD | 6.10 | 22 | 8.87 (0.30) | 9.21 (0.21) | 8.54 (98.31) | 1274 (74.76) |
| 7 | 51.2208, 19.4339 | 604.5 | CL | FPMD | 6.12 | 22 | 8.96 (0.34) | 9.09 (0.22) | 7.4 (95.55) | 1075 (69.15) | ||
| 15 | 51.2105, 19.4384 | 900.0 | Ps | S | FPM | 5.01 | 18 | 5.49 (0.07) | 4.86 (0.04) | 12.76 (99.83) | 5022 (99.12) | |
| 16 | 51.2117, 19.4265 | 810.0 | S | FPM | 4.93 | 17 | 6.81 (0.11) | 6.05 (0.05) | 20.61 (99.96) | 5123 (99.28) | ||
| 105 | 51.2744, 19.4324 | 646.5 | Bp | FH | SL | FMD | 5.55 | 20 | 10.40 (0.21) | 13.66 (0.11) | 15.79 (99.95) | 1779 (92.00) |
| 106 | 51.2747, 19.4326 | 450.0 | CL | FMD | 5.57 | 20 | 10.10 (0.20) | 13.44 (0.12) | 18.32 (99.99) | 2200 (95.19) | ||
| 107 | 51.1738, 19.4323 | 750.0 | Ps | S | FMM | 4.68 | 21 | 8.02 (0.15) | 8.90 (0.08) | 23.41 (100.00) | 4200 (99.68) | |
| 108 | 51.1746, 19.4321 | 750.0 | S | FM | 4.37 | 21 | 8.46 (0.16) | 9.16 (0.08) | 21.22 (99.81) | 3467 (99.24) |
| Habitat Type | Stand Type | Litter Type | Total | |||
|---|---|---|---|---|---|---|
| Birch | Oak | Alder | Pine | |||
| FH | Bp | 760 | 942 | 860 | 783 | 3345 |
| Ps | 436 | 525 | 442 | 426 | 1829 | |
| SH | Bp | 210 | 487 | 129 | 155 | 981 |
| Ps | 60 | 109 | 51 | 91 | 311 | |
| Total | 1466 | 2063 | 1482 | 1455 | - | |
| Habitat Type | Stand Type | Litter Type | Total | |||
|---|---|---|---|---|---|---|
| Birch | Oak | Alder | Pine | |||
| FH | Bp | 22 | 23 | 25 | 26 | 37 |
| Ps | 21 | 22 | 19 | 19 | 30 | |
| SH | Bp | 15 | 19 | 8 | 16 | 22 |
| Ps | 12 | 15 | 10 | 13 | 21 | |
| Total | 39 | 43 | 33 | 39 | - | |
| No. | Species | Family | Total No. of Specimens | FH | SH | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bp | Ps | Bp | Ps | ||||||||||||||||||||
| F | M | D | P | L | F | M | D | P | L | F | M | D | P | L | F | M | D | P | L | ||||
| 1 | Alliphis halleri (G. et R. Canestrini, 1881) | Eviphidae | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | Amblyseius sp. | Phytoseiidae | 35 | - | - | - | - | - | 2 | - | - | - | - | 27 | 2 | - | - | - | 3 | - | - | 1 | - |
| 3 | Amblyseius tubae Karg, 1970 | Phytoseiidae | 775 | 26 | - | - | - | - | 8 | - | - | - | 1 | 596 | 58 | - | 24 | 1 | 58 | 2 | - | 1 | - |
| 4 | Antennoseius bacatus Athias-Henriot, 1961 | Ascidae | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | - |
| 5 | Antennoseius masoviae Sellnick, 1943 | Ascidae | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| 6 | Arctoseius brevichelis Karg, 1969 | Ascidae | 1 | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| 7 | Arctoseius insularis (Willmann, 1952) | Ascidae | 45 | - | - | - | - | - | - | - | - | - | - | 8 | - | - | - | - | 32 | 5 | - | - | - |
| 8 | Asca aphidioides (Linneaeus, 1758) | Ascidae | 307 | 88 | - | - | - | - | 100 | - | - | 4 | - | 69 | - | 1 | 4 | - | 36 | 2 | 1 | 2 | - |
| 9 | Asca bicornis (Canestrini et Fanzago, 1887) | Ascidae | 150 | 1 | - | - | - | - | - | - | - | - | - | 75 | 19 | - | 2 | - | 44 | 8 | - | 1 | - |
| 10 | Cheiroseius borealis (Berlese, 1904) | Ascidae | 1 | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| 11 | Dendrolaelaps angulosus (Willmann, 1936) | Digamasellidae | 8 | - | - | - | - | - | - | - | - | - | - | 4 | - | - | - | - | - | - | 4 | - | - |
| 12 | Dendrolaelaps foveolatus (Leitner, 1949) | Digamasellidae | 8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | 5 | - | - |
| 13 | Dendrolaelaps laetus Shcherbak, 1980 | Digamasellidae | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 14 | Dendrolaelaps latior (Leitner, 1949) | Digamasellidae | 2 | 1 | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| 15 | Dendrolaelaps punctatosimilis Hirschmann, 1960 | Digamasellidae | 7 | 2 | - | 2 | - | - | - | - | - | - | - | - | - | 3 | - | - | - | - | - | - | - |
| 16 | Dendrolaelaps rotundus Hirschmann, 1960 | Digamasellidae | 4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 3 | - | - |
| 17 | Dinychus perforatus Kramer, 1882 | Uropodiade | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 18 | Epicriopsis horridus Kramer, 1876 | Ameroseiidae | 13 | 10 | 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 19 | Eviphis ostrinus (C.L. Koch, 1836) | Eviphidae | 4 | 3 | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 20 | Gamasellodes bicolor (Berlese, 1918) | Ascidae | 4 | 1 | - | - | - | - | 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 21 | Geholaspis longispinosus (Kramer, 1876) | Macrochelidae | 8 | 6 | - | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 22 | Hypoaspis praesternalis Willmann, 1949 | Laelapidae | 15 | - | - | - | - | - | 1 | - | - | - | - | 6 | - | - | - | - | 6 | 2 | - | - | - |
| 23 | Hypoaspis vacua (Michael, 1891) | Laelapidae | 6 | 2 | - | - | - | - | 2 | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| 24 | Iphidozercon gibbus (Berlese, 1903) | Ascidae | 18 | 15 | 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 25 | Laelaspis astronomicus (C.L. Koch, 1839) | Laelapidae | 3 | - | - | - | - | - | 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 26 | Lasioseius confusus Evans, 1958 | Ascidae | 1 | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 27 | Lasioseius muricatus (C.L. Koch, 1839) | Ascidae | 1 | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 28 | Leioseius elongatus Evans, 1958 | Ascidae | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 29 | Leptogamasus anoxygenenellus (Micherdziński, 1969) | Parasitidae | 1 | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 30 | Leptogamasus belligerens Witaliński, 1973 | Parasitidae | 6 | - | - | - | - | - | 3 | 3 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 31 | Leptogamasus parvulus (Berlese, 1903) | Parasitidae | 112 | 1 | 1 | - | - | - | 44 | 62 | 1 | - | - | 1 | 1 | - | - | - | 1 | - | - | - | - |
| 32 | Macrocheles montanus (Willmann, 1951) | Macrochelidae | 9 | 5 | - | 2 | 1 | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| 33 | Nejordensia levis (Oudemans et Voigts, 1904) | Ascidae | 1 | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| 52 | Olodiscus minima Kramer, 1882 | Uropodidae | 287 | 215 | 1 | 64 | 3 | - | - | - | - | - | - | 1 | - | - | - | - | 3 | - | - | - | - |
| 34 | Paragamasus conus (Karg, 1971) | Parasitidae | 12 | 3 | 2 | - | - | - | 3 | - | - | - | - | - | 4 | - | - | - | - | - | - | - | - |
| 35 | Paragamasus jugincola Athias-Henriot, 1967 | Parasitidae | 72 | 39 | 27 | - | - | - | 1 | 5 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 36 | Paragamasus runcatellus (Berlese, 1903) | Parasitidae | 90 | 51 | 15 | - | - | - | 14 | 7 | - | - | - | 1 | 1 | - | - | - | 1 | - | - | - | - |
| 37 | Paragamasus sp. | Parasitidae | 71 | - | - | 17 | 7 | 5 | - | - | 21 | 15 | - | - | - | 3 | 1 | - | - | - | - | 2 | - |
| 38 | Parazercon radiatus Berlese, 1910 | Zerconidae | 16 | - | - | - | - | - | 16 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 39 | Pergamasus barbarus (Berlese, 1904) | Parasitidae | 5 | - | - | 4 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 40 | Pergamasus brevicornis (Berlese, 1903) | Parasitidae | 5 | 1 | - | - | - | - | 4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 41 | Pergamasus crassipes (Linneaeus, 1758) | Parasitidae | 62 | 9 | 4 | - | - | - | 2 | 4 | - | - | - | 9 | 4 | - | - | - | 10 | 19 | 1 | - | - |
| 42 | Pergamasus mediocris Berlese, 1904 | Parasitidae | 4 | - | - | - | - | - | 2 | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| 43 | Pergamasus septentrionalis (Oudemans, 1902) | Parasitidae | 52 | 39 | 3 | - | 2 | - | - | - | - | - | - | 7 | 1 | - | - | - | - | - | - | - | - |
| 44 | Pergamasus sp. | Parasitidae | 138 | - | - | 20 | 35 | 2 | - | - | 19 | 11 | - | - | - | 8 | 8 | - | - | - | 17 | 18 | - |
| 45 | Prozercon kochi Sellnick, 1943 | Zerconidae | 39 | - | - | - | - | - | 20 | 10 | 7 | 2 | - | - | - | - | - | - | - | - | - | - | - |
| 46 | Prozercon tragardhi (Halbert, 1923) | Zerconidae | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 47 | Sejus togatus C.L. Koch, 1836 | Sejoidea | 1 | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 48 | Trachytes aegrota (C.L. Koch, 1841) | Trachytidae | 710 | 350 | - | 123 | 173 | 27 | 15 | - | 10 | 8 | - | - | - | - | - | - | - | - | 1 | 1 | 2 |
| 49 | Trichouropoda ovalis (C.L. Koch, 1839) | Trematuridae | 14 | 1 | 1 | 6 | - | - | 2 | - | 4 | - | - | - | - | - | - | - | - | - | - | - | - |
| 50 | Typhlodromus pyri Scheuten, 1857 | Phytoseiidae | 1 | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 51 | Urobovella pulchella (Berlese, 1904) | Urodinychidae | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 53 | Veigaia cervus (Kramer, 1876) | Veigaiaiidae | 114 | 37 | - | 3 | 2 | - | 26 | - | 20 | 5 | - | 14 | - | 5 | 2 | - | - | - | - | - | - |
| 54 | Veigaia kochi (Trägarth, 1901) | Veigaiaiidae | 2 | - | - | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 55 | Veigaia nemorensis (C.L. Koch, 1839) | Veigaiaiidae | 892 | 432 | - | 223 | 122 | - | 50 | - | 39 | 25 | - | - | - | - | - | - | - | - | - | 1 | - |
| 56 | Veigaia planicola (Berlese, 1892) | Veigaiaiidae | 41 | 27 | - | 9 | 5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 57 | Vulgarogamasus kraepelini (Berlese, 1904) | Parasitidae | 2 | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 58 | Zercon peltatus C.L. Koch, 1836 | Zerconidae | 2281 | 509 | 321 | 126 | 68 | 25 | 671 | 52 | 250 | 98 | 144 | 4 | - | 3 | - | - | 5 | 1 | 3 | 1 | - |
| 59 | Zerconopsis muestairi (Schweizer, 1949) | Ascidae | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| In total | 6466 | F | M | D | P | L | F | M | D | P | L | F | M | D | P | L | F | M | D | P | L | ||
| 1881 | 382 | 602 | 421 | 59 | 995 | 147 | 373 | 169 | 145 | 826 | 90 | 23 | 41 | 1 | 201 | 45 | 35 | 28 | 2 | ||||
| 3345 | 1829 | 981 | 311 | ||||||||||||||||||||
| Habitat Type | Stand Type | Species Rank | Taxon | Collection Month (Date) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 6 | 9 | 12 | 18 | 24 | 30 | 36 | Total | ||||
| (10.03.2014) | (09.06.2014) | (10.09.2014) | (10.12.2014) | (10.06.2015) | (09.12.2015) | (09.06.2016) | (13-14.12.2016) | |||||
| FH | Bp | 1 | Z. peltatus | 280 | 141 | 163 | 111 | 68 | 104 | 109 | 73 | 1049 |
| 2 | V. nemorensis | 5 | 10 | 145 | 234 | 106 | 55 | 44 | 178 | 777 | ||
| 3 | T. aegrota | 1 | 139 | 225 | 128 | 32 | 42 | 3 | 103 | 673 | ||
| 4 | O. minima | 34 | 51 | 31 | 40 | 61 | 19 | 47 | 283 | |||
| 5 | A. aphidioides | 1 | 5 | 9 | 21 | 9 | 43 | 88 | ||||
| 6 | P. jugincola | 4 | 1 | 22 | 1 | 38 | 66 | |||||
| 6 | P. runcatellus | 13 | 38 | 5 | 2 | 1 | 7 | 66 | ||||
| 7 | Pergamasus sp. | 2 | 21 | 1 | 26 | 6 | 1 | 57 | ||||
| 8 | P. septentrionalis | 11 | 29 | 4 | 44 | |||||||
| 9 | V. cervus | 3 | 23 | 6 | 2 | 3 | 5 | 42 | ||||
| 10 | V. planicola | 8 | 18 | 15 | 41 | |||||||
| Ps | 1 | Z. peltatus | 2 | 68 | 339 | 228 | 158 | 202 | 1 | 217 | 1215 | |
| 2 | V. nemorensis | 25 | 6 | 21 | 47 | 15 | 114 | |||||
| 3 | L. parvulus | 2 | 9 | 16 | 4 | 46 | 30 | 107 | ||||
| 4 | A. aphidioides | 2 | 72 | 1 | 15 | 3 | 11 | 104 | ||||
| 5 | V. cervus | 19 | 3 | 22 | 2 | 5 | 51 | |||||
| 6 | P. kochi | 2 | 1 | 10 | 16 | 10 | 39 | |||||
| 7 | Paragamasus sp. | 2 | 5 | 27 | 1 | 1 | 36 | |||||
| 8 | T. aegrota | 26 | 4 | 3 | 0 | 33 | ||||||
| 9 | Pergamasus sp. | 3 | 10 | 14 | 2 | 1 | 30 | |||||
| 10 | P. runcatellus | 4 | 2 | 8 | 5 | 2 | 21 | |||||
| SH | Bp | 1 | A. tubae | 295 | 153 | 15 | 116 | 6 | 94 | 679 | ||
| 2 | A. bicornis | 42 | 36 | 1 | 14 | 3 | 96 | |||||
| 3 | A. aphidioides | 73 | 1 | 74 | ||||||||
| 4 | Amblyseius sp. | 9 | 8 | 1 | 2 | 1 | 8 | 29 | ||||
| 5 | V. cervus | 13 | 2 | 1 | 5 | 21 | ||||||
| 6 | Pergamasus sp. | 13 | 1 | 1 | 1 | 16 | ||||||
| 7 | P. crassipes | 1 | 4 | 3 | 2 | 3 | 13 | |||||
| Ps | 1 | A. tubae | 19 | 3 | 1 | 35 | 3 | 61 | ||||
| 2 | A. bicornis | 41 | 4 | 6 | 2 | 53 | ||||||
| 3 | A. aphidioides | 24 | 1 | 14 | 1 | 1 | 41 | |||||
| 4 | A. insularis | 1 | 2 | 20 | 2 | 6 | 6 | 37 | ||||
| 5 | Pergamasus sp. | 29 | 4 | 2 | 35 | |||||||
| 6 | P. crassipes | 17 | 6 | 2 | 2 | 3 | 30 | |||||
| 7 | Z. peltatus | 3 | 2 | 5 | 10 | |||||||
| 8 | D. foveolatus | 1 | 7 | 8 | ||||||||
| 9 | H. praesternalis | 1 | 1 | 4 | 2 | 8 | ||||||
| 10 | Amblyseius sp. | 1 | 1 | 2 | 4 | |||||||
| 11 | D. angulosus | 2 | 2 | 4 | ||||||||
| 12 | D. rotundus | 1 | 3 | 4 | ||||||||
| 13 | T. aegrota | 1 | 1 | 2 | 4 | |||||||
References
- Berg, B.; McClaugherty, C. Plant Litter, Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Hobbie, S.E. Plant species effects on nutrient cycling: Revisiting litter feedbacks. Trends Ecol. Evol. 2015, 30, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Takeda, H. Succession of soil microarthropod communities during the aboveground and belowground litter decomposition processes. Soil Biol. Biochem. 2017, 110, 95–102. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Antunes, P.M.; Bennett, A.E.; Birkhofer, K.; Bissett, A.; Bowker, M.A.; Caruso, T.; Chen, B.; Coleman, D.C.; de Boer, W.; et al. Priorities for research in soil ecology. Pedobiologia 2017, 63, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, R.K.; Singh, J.S. Restoration of Mine Spoil in a Dry Tropical Region: A Review. Proc. Indian Natl. Sci. Acad. 2017, 83, 789–844. [Google Scholar] [CrossRef]
- Sako, A.; Semde, S.; Wenmenga, U. Geochemical evaluation of soil, surface water and groundwater around the Tongon gold mining area, northern Cote d’Ivoire, West Africa. J. Afr. Earth Sci. 2018, 145, 297–316. [Google Scholar] [CrossRef]
- Horodecki, P.; Nowiński, M.; Rawlik, K.; Jagodziński, A.M. Rozkład liści drzew w początkowych etapach dekompozycji w drzewostanach sosnowych i brzozowych rosnących na rekultywowanym zwałowisku pokopalnianym i terenach leśnych (Early stages of tree leaves decomposition in pine and birch stands growing on a reclaimed lignite mine spoil heap and forest areas). Studia i Materiały Centrum Edukacji Przyrodniczo-Leśnej w Rogowie 2015, 17, 262–278. [Google Scholar]
- Frouz, J. The effect of soil macrofauna on litter decomposition and soil organic matter accumulation during soil formation in spoil heaps after brown coal mining: A preliminary results. Ekol. Bratisl. 2002, 21, 363–369. [Google Scholar]
- Frouz, J. The effect of litter type and macrofauna community on litter decomposition and organic matter accumulation in post-mining sites. Biologia 2008, 63, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Frouz, J.; Roubíčková, A.; Heděnec, P.; Tajovský, K. Do soil fauna really hasten litter decomposition? A meta-analysis of enclosure studies. Eur. J. Soil Biol. 2015, 68, 18–24. [Google Scholar] [CrossRef]
- Camizuli, E.; Scheifler, R.; Garnier, S.; Monna, F.; Losno, R.; Gourault, C.; Hamm, G.; Lachiche, C.; Delivet, G.; Chateau, C.; et al. Trace metals from historical mining sites and past metallurgical activity remain bioavailable to wildlife today. Sci. Rep. 2018, 8, 3436. [Google Scholar] [CrossRef] [PubMed]
- Dunger, W.; Wanner, M.; Hauser, H.; Hohberg, K.; Schulz, H.-J.; Schwalbe, T.; Seifert, B.; Vogel, J.; Voigtländer, K.; Zimdars, B.; et al. Development of soil fauna at mine sites during 46 years after afforestation. Pedobiologia 2001, 45, 243–271. [Google Scholar] [CrossRef]
- Russell, D.J.; Hohberg, K.; Elmer, M. Primary colonisation of newly formed soils by actinedid mites. Soil Org. 2010, 82, 237–251. [Google Scholar]
- Murvanidze, M.; Mumladze, L.; Arabuli, T.; Kvavadze, E. Oribatid Mite Colonization of Sand and Manganese Tailing Sites. Acarologia 2013, 53, 203–216. [Google Scholar] [CrossRef]
- Wang, S.; Ruan, H. Effects of soil mesofauna and microclimate on nitrogen dynamics in leaf litter decomposition along an elevation gradient. Afr. J. Biotechnol. 2011, 10, 6732–6742. [Google Scholar]
- Berg, M.P.; Kniese, J.P.; Bedaux, J.J.M.; Verhoef, H.A. Dynamics and stratification of functional groups of micro- and mesoarthropods in the organic layer of a Scots pine forest. Biol. Fertil. Soils 1998, 26, 268–284. [Google Scholar] [CrossRef]
- Boczek, J.; Błaszak, C. Roztocze (Acari)—Znaczenie w Życiu i Gospodarce Człowieka; SGGW: Warszawa, Poland, 2005. [Google Scholar]
- Koehler, H.H. Predatory mites (Gamasina, Mesostigmata). Agric. Ecosyst. Environ. 1999, 74, 395–410. [Google Scholar] [CrossRef]
- Salmane, I.; Brumelis, G. The importance of the moss layer in sustaining biological diversity of Gamasina mites in coniferous forest soil. Pedobiologia 2008, 52, 69–76. [Google Scholar] [CrossRef]
- Koehler, H.H. Mesostigmata (Gamasina, Uropodina), efficient predators in agroecosystems. Agric. Ecosyst. Environ. 1997, 62, 105–117. [Google Scholar] [CrossRef]
- Reeves, W.K.; Dowling, A.P.G.; Dasch, G.A. Rickettsial agents from parasitic Dermanyssoidea (Acari: Mesostigmata). Exp. Appl. Acarol. 2006, 38, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Wissuwa, J.; Salamon, J.-A.; Frank, T. Effects of habitat age and plant species on predatory mites (Acari, Mesostigmata) in grassy arable fallows in Eastern Austria. Soil Biol. Biochem. 2012, 50, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Ruf, A.; Beck, L. The use of predatory soil mites in ecological soil classification and assessment concepts, with perspectives for oribatid mites. Ecotox. Environ. Saf. 2005, 62, 290–299. [Google Scholar] [CrossRef] [PubMed]
- VreekenBuijs, M.J.; Brussaard, L. Soil mesofauna dynamics, wheat residue decomposition and nitrogen mineralization in buried litterbags. Biol. Fertil. Soils 1996, 23, 374–381. [Google Scholar] [CrossRef]
- Sławska, M. Springtails Collembola, Hexapoda and Mites Acari, Arachnida as indicators of healthy forest soils. Studia i Materiały Centrum Edukacji Przyrodniczo-Leśnej 2007, 9, 208–217. [Google Scholar]
- Beaulieu, F.; Weeks, A.R. Free-living mesostigmatic mites in Australia: Their roles in biological control and bioindication. Aust. J. Exp. Agric. 2007, 47, 460–478. [Google Scholar] [CrossRef]
- Koehler, H.H. Gamasina in a succession of thirteen years. In Ecology and Evolution of the Acari; Series Entomologica 55; Springer: Dordrecht, The Netherlands, 1999; pp. 531–539. [Google Scholar]
- Koehler, H.; Melecis, V. Long-term observations of soil mesofauna. In Long-Term Ecological Research; Muller, F., Baessler, C., Schubert, H., Klotz, S., Eds.; Springer: New York, NY, USA, 2010; ISBN 978-90-481-8781-2. [Google Scholar]
- Madej, G. Rozwój Zgrupowań Roztoczy Mesostigmata (Arachnida. Acari) na Nieużytkach Poprzemysłowych; Uniwersytet Śląski: Katowice, Poland, 2004. [Google Scholar]
- Madej, G.; Skubała, P. Colonization of a dolomitic dump by mesostigmatid mites (Acari, Mesostigmata). In Acarid Phylogeny and Evolution: Adaptation in Mites and Ticks; Bernini, F., Nannelli, R., Nuzaci, G., DeLillo, E., Eds.; Springer: Dordrecht, The Netherlands, 2002; ISBN 978-1-4020-0465-0. [Google Scholar]
- Madej, G.; Stodółka, A. Successional changes and diversity of mesostigmatid mite communities (Acari: Mesostigmata) on reclaimed power plant waste dumps. Ann. Zool. 2008, 58, 267–278. [Google Scholar] [CrossRef]
- Pastwik, E.; Skorupski, M.; Piasta, A.; Jagodziński, A.M. Mesostigmata mites of afforested post-industrial habitats on lignite mine spoil heap in Bełchatów—A preliminary study. In Proceedings of the Science for Sustainability—International Scientific Conference for PhD Students, University of West Hungary, Győr, Hungary, 19–20 Marth 2013; Neményi, M., Varga, L., Facskó, F., Lőrincz, I., Eds.; University of West Hungary Press: Sopron, Hungary, 2013; pp. 251–257. [Google Scholar]
- Skorupski, M.; Horodecki, P.; Jagodziński, A.M. Roztocze z rzędu Mesostigmata (Arachnida, Acari) na terenach przemysłowych i poprzemysłowych w Polsce. Nauka Przyr. Technol. 2013, 7, 11. [Google Scholar]
- Piasta, A.; Skorupski, M.; Horodecki, P.; Jagodziński, A.M. Zgrupowania roztoczy (Acari) pod drzewostanami sosnowymi na terenach leśnych i rekultywowanym zwałowisku zewnętrznym w Nadleśnictwie Bełchatów (Soil mite communities under Scots pine stands growing on forest sites and reclaimed lignite mine spoil heap in Bełchatów Forest District). Studia i Materiały Centrum Edukacji Przyrodniczo-Leśnej w Rogowie 2015, 17, 279–294. [Google Scholar]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Żytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Mueller, K.E.; Eisenhauer, N.; Reich, P.B.; Hobbie, S.E.; Chadwick, O.A.; Chorover, J.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; Kałucka, I.; et al. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol. Biochem. 2016, 92, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Skorupski, M. Influence of Selected Tree Species on Forest Ecosystem Biodiversity for the Example of Mesostigmata Mites in a Common-Garden Experiment; Uniwersytet Przyrodniczy w Poznaniu: Poznań, Poland, 2010. [Google Scholar]
- Gergocs, V.; Hufnagel, L. The effect of microarthropods on litter decomposition depends on litter quality. Eur. J. Soil Biol. 2016, 75, 24–30. [Google Scholar] [CrossRef]
- Gao, M.; Li, J.; Zhang, X. Responses of soil fauna structure and leaf litter decomposition to effective microorganism treatments in Da Hinggan Mountains, China. Chin. Geogr. Sci. 2012, 22, 647–658. [Google Scholar] [CrossRef]
- Kaneko, N.; Salamanca, E. Mixed leaf litter effects on decomposition rates and soil microarthropod communities in an oak–pine stand in Japan. Ecol. Res. 1999, 14, 131–138. [Google Scholar] [CrossRef]
- Riutta, T.; Slade, E.M.; Bebber, D.P.; Taylor, M.E.; Malhi, Y.; Riordan, P.; Macdonald, D.W.; Morecroft, M.D. Experimental evidence for the interacting effects of forest edge, moisture and soil macrofauna on leaf litter decomposition. Soil Biol. Biochem. 2012, 49, 124–131. [Google Scholar] [CrossRef]
- Slade, E.M.; Riutta, T. Interacting effects of leaf litter species and macrofauna on decomposition in different litter environments. Basic Appl. Ecol. 2012, 13, 423–431. [Google Scholar] [CrossRef]
- Singh, K.P.; Singh, P.K.; Tripathi, S.K. Litterfall, litter decomposition and nutrient release patterns in four native tree species raised on coal mine spoil at Singrauli, India. Biol. Fertil. Soil. 1999, 29, 371–378. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Oleksyn, J.; Karolewski, P.; Horodecki, P.; Rawlik, K.; Żytkowiak, R.; Chałupka, W.; Chmura, D.J.; Guzicka, M.; Misiorny, A.; et al. Raport Końcowy z Realizacji Usługi Badawczej pt. “Środowiskowo-Genetyczne Uwarunkowania Produktywności Ekosystemów Leśnych na Gruntach Leśnych i Poprzemysłowych”.; Na Podstawie Umowy nr OR/2717/3/11 z dnia 2 Lutego 2011 roku.; Institute of Dendrology, Polish Academy of Sciences: Kórnik, Poland, 2015. [Google Scholar]
- Horodecki, P.; Jagodzinski, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Hopkins, D.W.; Badalucco, L.; English, L.C.; Meli, S.M.; Chudek, J.A.; Ioppolo, A. Plant litter decomposition and microbial characteristics in volcanic soils (Mt Etna, Sicily) at different stages of development. Biol. Fertil. Soil 2007, 43, 461–469. [Google Scholar] [CrossRef]
- Esperschuetz, J.; Zimmermann, C.; Duemig, A.; Welzl, G.; Buegger, F.; Elmer, M.; Munch, J.C.; Schloter, M. Dynamics of microbial communities during decomposition of litter from pioneering plants in initial soil ecosystems. Biogeosciences 2013, 10, 5115–5124. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Wierzcholska, S.; Dyderski, M.K.; Horodecki, P.; Rusińska, A.; Gdula, A.K.; Kasprowicz, M. Tree species effects on bryophyte guilds on a reclaimed post-mining site. Ecol. Eng. 2018, 110, 117–127. [Google Scholar] [CrossRef]
- Rawlik, M.; Kasprowicz, M.; Jagodziński, A.M.; Kaźmierowski, C.; Łukowiak, R.; Grzebisz, W. Canopy tree species determine herb layer biomass and species composition on a reclaimed mine spoil heap. Sci. Total Environ. 2018, 635, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Pietrzykowski, M. Scots pine (Pinus sylvestris L.) ecosystem macronutrients budget on reclaimed mine sites—Stand trees supply and stability. Nat. Sci. 2010, 2, 590–599. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Daniels, W.L. Estimation of carbon sequestration by pine (Pinus sylvestris L.) ecosystems developed on reforested post-mining sites in Poland on differing mine soil substrates. Ecol. Eng. 2014, 73, 209–218. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Gdula, A.K.; Rawlik, K.; Kasprowicz, M. Zróżnicowanie flory roślin naczyniowych runa pod drzewostanami powstałymi w wyniku rekultywacji zwałowiska pokopalnianego (Differentiation of the flora of herbaceous layer under tree stands established for the reclamation of the lignite mine spoil heap). Studia i Materiały Centrum Edukacji Przyrodniczo-Leśnej w Rogowie 2015, 17, 249–261. [Google Scholar]
- Horodecki, P.; Nowiński, M.; Jagodziński, A.M. Advantages of mixed tree stands in restoration of upper soil layers on post-mining sites: A five-year leaf litter decomposition experiment. Land Degrad. Dev. 2018. [Google Scholar] [CrossRef]
- Kamczyc, J.; Urbanowski, C.; Pers-Kamczyc, E. Mite communities (Acari: Mesostigmata) in young and mature coniferous forests after surface wildfire. Exp. Appl. Acarol. 2017, 72, 145–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skubała, P. Development of oribatid mite communities (Acari, Oribatida) on a mine dump. In Acarid Phylogeny and Evolution: Adaptation in Mites and Ticks; Bernini, F., Nannelli, R., Nuzaci, G., DeLillo, E., Eds.; Springer: Dordrecht, The Netherlands, 2002; ISBN 978-1-4020-0465-0. [Google Scholar]
- Ulrich, W.; Zalewski, M. Abundance and co-occurrence patterns of core and satellite species of ground beetles on small lake islands. Oikos 2006, 114, 338–348. [Google Scholar] [CrossRef]
- Seniczak, S.; Graczyk, R.; Seniczak, A.; Faleńczyk-Koziróg, K.; Kaczmarek, S.; Marquardt, T. Microhabitat preferences of Oribatida and Mesostigmata (Acari) inhabiting lowland beech forest in Poland and the trophic interactions between these mites. Eur. J. Soil Biol. 2018, 87, 25–32. [Google Scholar] [CrossRef]
- Helingerová, M.; Frouz, J.; Šantrůčková, H. Microbial activity in reclaimed and unreclaimed post-mining sites near Sokolov (Czech Republic). Ecol. Eng. 2010, 36, 768–776. [Google Scholar] [CrossRef]
- Hagvar, S. From Litter to Humus in a Norwegian Spruce Forest: Long-Term Studies on the Decomposition of Needles and Cones. Forests 2016, 7, 186. [Google Scholar] [CrossRef]
- Gonzalez, G.; Seastedt, T.R.; Donato, Z. Earthworms, arthropods and plant litter decomposition in aspen (Populus tremuloides) and lodgepole pine (Pinus contorta) forests in Colorado, USA. Pedobiologia 2003, 47, 863–869. [Google Scholar] [CrossRef]
- Gergocs, V.; Rethati, G.; Hufnagel, L. Litter quality indirectly influences community composition, reproductive mode and trophic structure of oribatid mite communities: A microcosm experiment. Exp. Appl. Acarol. 2015, 67, 335–356. [Google Scholar] [CrossRef] [PubMed]
- Karg, W. Synökologische Untersuchungen von Bodenmilben aus forstwirtschaftlich und landwirtschaftlich genutzten Böden. Pedobiologia 1967, 7, 198–214. [Google Scholar]
- Jurksiene, G.; Janusauskaite, D.; Armolaitis, K.; Baliuckas, V. Leaf litterfall decomposition of pedunculate (Quercus robur L.) and sessile (Q. petraea [Matt.] Liebl.) oaks and their hybrids and its impact on soil microbiota. Dendrobiology 2017, 78, 51–62. [Google Scholar] [CrossRef]
- Osler, G.H.R.; Gauci, C.S.; Abbott, L.K. Limited evidence for short-term succession of microarthropods during early phases of surface litter decomposition. Pedobiologia 2004, 48, 37–49. [Google Scholar] [CrossRef]
- Schmidt, R.A. Leaf structures affect predatory mites (Acari: Phytoseiidae) and biological control: A review. Exp. Appl. Acarol. 2014, 62, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Maraun, M.; Scheu, S. The structure of oribatid mite communities (Acari, Oribatida): Patterns, mechanisms and implications for future research. Ecography 2000, 23, 374–382. [Google Scholar] [CrossRef]
- Erdmann, G.; Scheu, S.; Maraun, M. Regional factors rather than forest type drive the community structure of soil living oribatid mites (Acari, Oribatida). Exp. Appl. Acarol. 2012, 57, 157–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonari, G.; Migliorini, M.; Landi, M.; Protano, G.; Fanciulli, P.P.; Angiolini, C. Concordance between plant species, oribatid mites and soil in a Mediterranean stone pine forest. Arthropod-Plant Interact. 2017, 11, 61–69. [Google Scholar] [CrossRef]
- Karg, W. Acari (Acarina) Milben Parasitiformes (Anactinochaeta), Cohors Gamasina Leach Raubmmilben—Die Tierwelt Deutschlands; VEB Gustav Fischer Ver Lag: Jena, Germany, 1993; Volume 59. [Google Scholar]




| Fixed Factor | DF | Wald Chi-Square | Prob > Chi-Square | |||
|---|---|---|---|---|---|---|
| Abundance | Species Richness | Abundance | Species Richness | Abundance | Species Richness | |
| CM | 7 | 7 | 141.972 | 15.689 | <0.0001 | 0.0281 |
| LT | 3 | 3 | 8.461 | 0.282 | 0.0374 | 0.9633 |
| HT | 1 | 1 | 22.590 | 1.393 | <0.0001 | 0.2379 |
| ST | 1 | 1 | 7.111 | 0.025 | 0.0077 | 0.8752 |
| CM × LT | 21 | 21 | 78.628 | 10.756 | <0.0001 | 0.9673 |
| CM × HT | 7 | 7 | 1365.612 | 7.949 | <0.0001 | 0.3371 |
| CM × ST | 7 | 7 | 69.675 | 3.709 | <0.0001 | 0.8126 |
| LT × HT | 3 | 3 | 6.848 | 2.115 | 0.0769 | 0.5489 |
| LT × ST | 3 | 3 | 10.256 | 1.578 | 0.0165 | 0.6644 |
| HT × ST | 1 | 1 | 0.797 | 1.159 | 0.3719 | 0.2818 |
| CM × LT × HT | 21 | 21 | 917.881 | 14.963 | <0.0001 | 0.8248 |
| CM × LT × ST | 21 | 21 | 237.841 | 12.251 | <0.0001 | 0.9326 |
| CM × HT × ST | 7 | 7 | 12.618 | 1.931 | 0.082 | 0.9635 |
| LT × HT × ST | 3 | 3 | 5.563 | 1.153 | 0.1349 | 0.7644 |
| CM × LT × HT × ST | 21 | 21 | 69.846 | 14.247 | <0.0001 | 0.8588 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbanowski, C.K.; Horodecki, P.; Kamczyc, J.; Skorupski, M.; Jagodziński, A.M. Succession of Mite Assemblages (Acari, Mesostigmata) during Decomposition of Tree Leaves in Forest Stands Growing on Reclaimed Post-Mining Spoil Heap and Adjacent Forest Habitats. Forests 2018, 9, 718. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110718
Urbanowski CK, Horodecki P, Kamczyc J, Skorupski M, Jagodziński AM. Succession of Mite Assemblages (Acari, Mesostigmata) during Decomposition of Tree Leaves in Forest Stands Growing on Reclaimed Post-Mining Spoil Heap and Adjacent Forest Habitats. Forests. 2018; 9(11):718. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110718
Chicago/Turabian StyleUrbanowski, Cezary K., Paweł Horodecki, Jacek Kamczyc, Maciej Skorupski, and Andrzej M. Jagodziński. 2018. "Succession of Mite Assemblages (Acari, Mesostigmata) during Decomposition of Tree Leaves in Forest Stands Growing on Reclaimed Post-Mining Spoil Heap and Adjacent Forest Habitats" Forests 9, no. 11: 718. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110718