Cynomolgus Monkeys (Macaca fascicularis) as an Experimental Infection Model for Human Group A Rotavirus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Virus Inoculum
2.3. Experimental Design
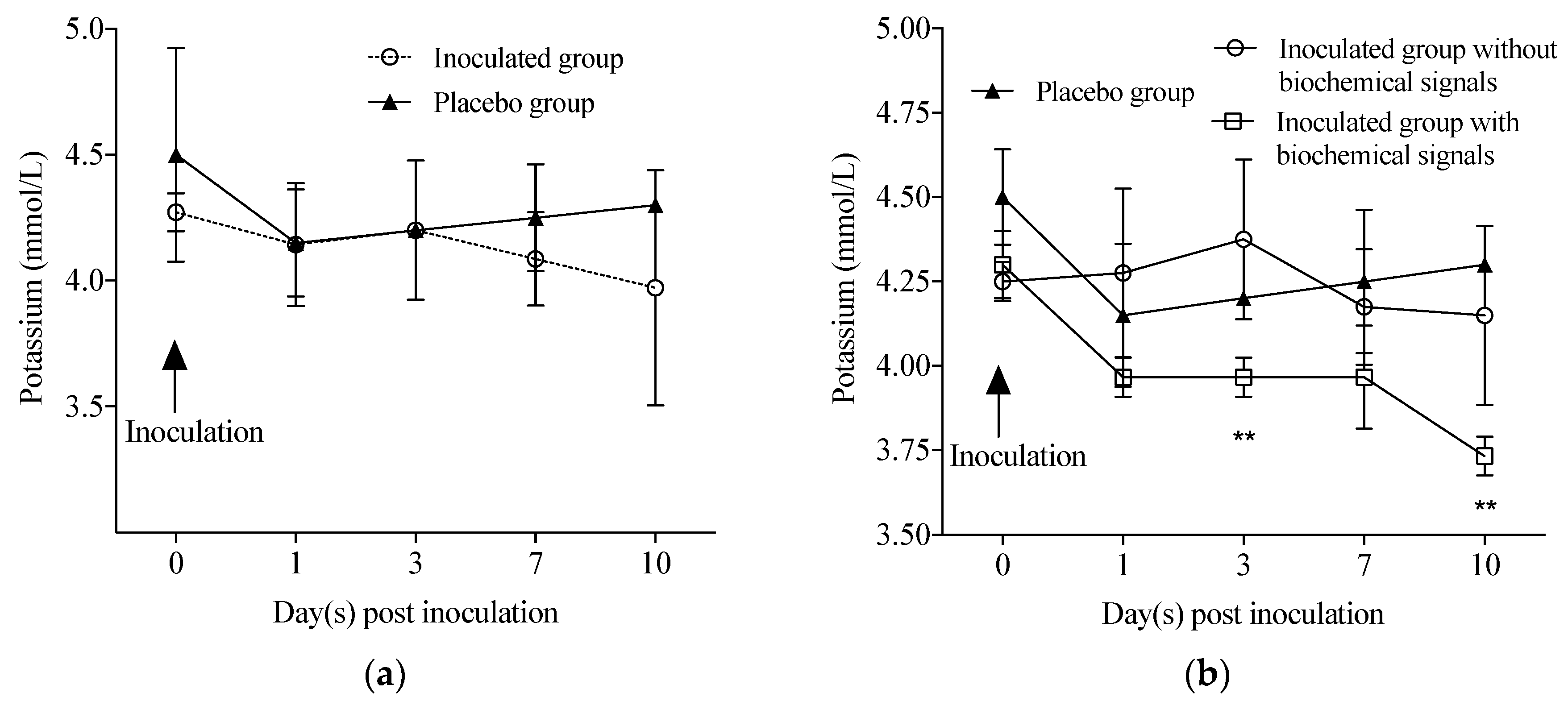
2.4. Haematological and Biochemistry Analyses
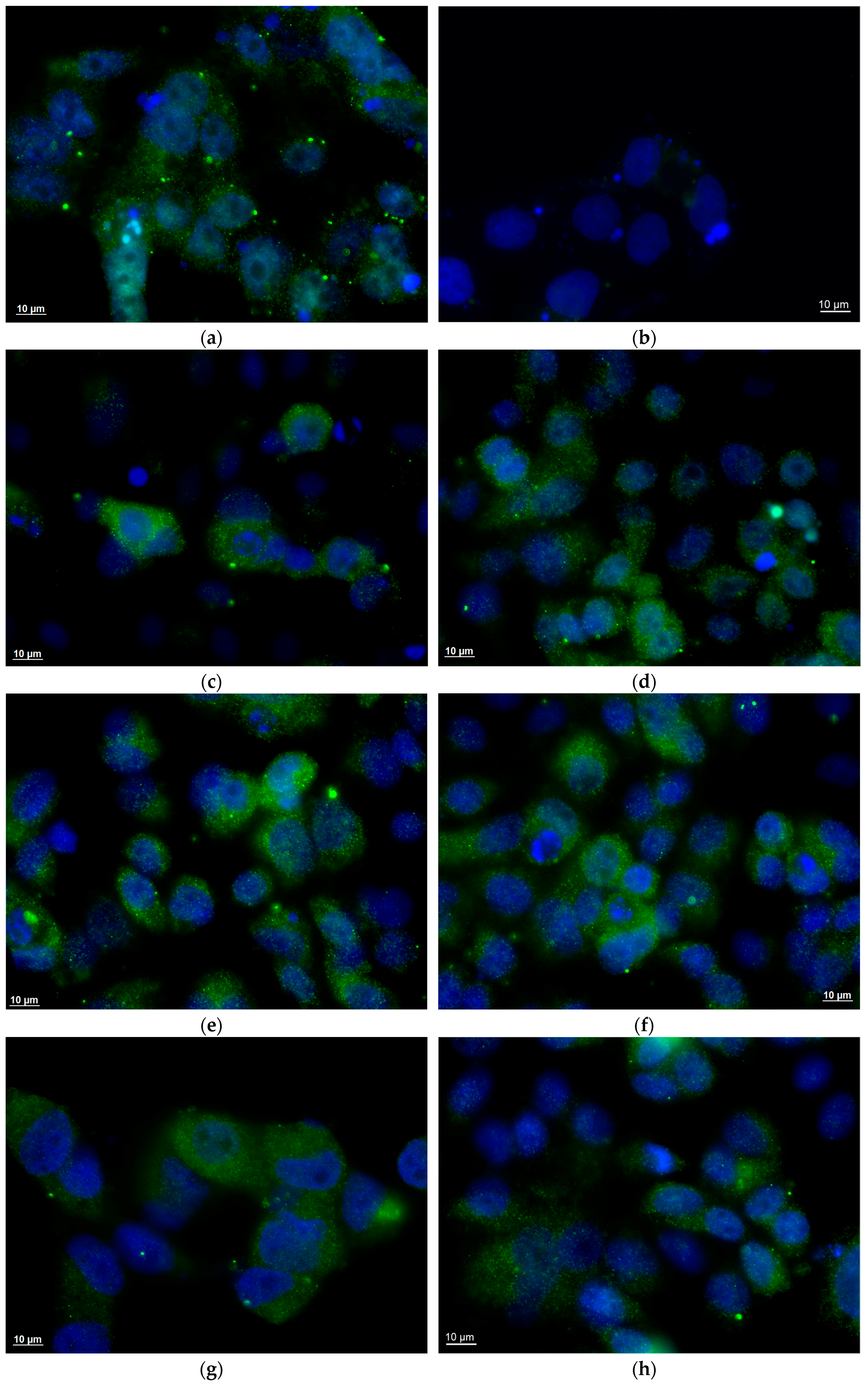
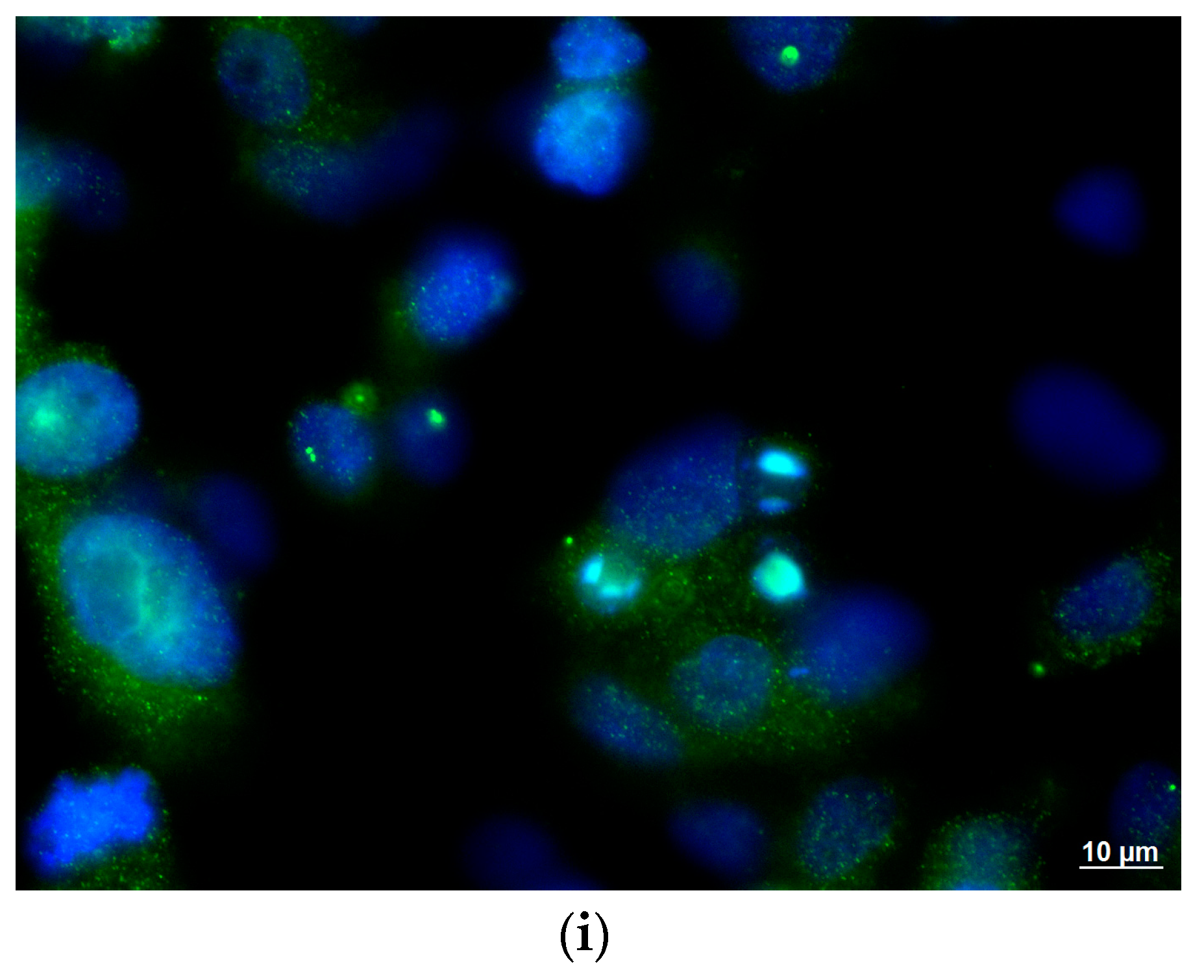
2.5. Human Group A Rotavirus Detection by Enzyme Immunoassay
2.6. Qualitative and Quantitative Human Group A Rotavirus RNA Detection by Molecular Amplification Procedures
2.7. Evidence of Infectious Particles from Faeces
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Madsen, L.B.; Ustrup, M.; Hansen, K.S.; Nyasulu, P.S.; Bygbjerg, I.C.; Konradsen, F. Estimating the costs of implementing the rotavirus vaccine in the national immunisation programme: The case of Malawi. Trop. Med. Int. Health 2014, 19, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Rotavirus vaccines: WHO position paper—January 2013. Wkly. Epidemiol. Rec. 2013, 88, 49–64.
- Parashar, U.D.; Nelson, E.A.S.; Kang, G. Diagnosis, management, and prevention of rotavirus gastroenteritis in children. BMJ 2013, 347, f7204. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.M.; Glass, R.; Desai, R.; Tate, J.E.; Parashar, U.D. Fulfilling the promise of rotavirus vaccines: How far have we come since licensure? Lancet Infect. Dis. 2012, 12, 561–570. [Google Scholar] [CrossRef]
- Velázquez, F.R.; Matson, D.O.; Calva, J.J.; Guerrero, L.; Morrow, A.L.; Carter-Campbell, S.; Glass, R.I.; Estes, M.K.; Pickering, L.K.; Ruiz-Palacios, G.M. Rotavirus infection in infants as protection against subsequent infections. N. Engl. J. Med. 1996, 335, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Gentsch, J.R.; Glass, R.I. The role of serum antibodies in the protection against rotavirus disease: An overview. Clin. Infect. Dis. 2002, 34, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, D.R.; Wells, P.W. Passive immunity in rotaviral infections. J. Am. Vet. Med. Assoc. 1978, 173, 565–568. [Google Scholar] [PubMed]
- Fernandez, F.M.; Conner, M.E.; Hodgins, D.C.; Parwani, A.V.; Nielsen, P.R.; Crawford, S.E.; Estes, M.K.; Saif, L.J. Passive immunity to bovine rotavirus in newborn calves fed colostrum supplements from cows immunized with recombinant SA11 rotavirus core-like particle (CLP) or virus-like particle (VLP) vaccines. Vaccine 1998, 16, 507–516. [Google Scholar] [CrossRef]
- Yuan, L.; Iosef, C.; Azevedo, M.S.; Kim, Y.; Qian, Y.; Geyer, A.; Nguyen, T.V.; Chang, K.O.; Saif, L.J. Protective immunity and antibody-secreting cell responses elicited by combined oral attenuated Wa human rotavirus and intranasal Wa 2/6-VLPs with mutant Escherichia coli heat-labile toxin in gnotobiotic pigs. J. Virol. 2001, 75, 9229–9238. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, M.S.; Yuan, L.; Jeong, K.-I.; Gonzalez, A.; Nguyen, T.V.; Pouly, S.; Gochnauer, M.; Zhang, W.; Azevedo, A.; Saif, L.J. Viremia and nasal and rectal shedding of rotavirus in gnotobiotic pigs inoculated with Wa human rotavirus. J. Virol. 2005, 79, 5428–5436. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Li, G.; Zhang, W.; Azevedo, M.S.P.; Saif, L.J.; Liu, F.; Bui, T.; Yousef, A.; Yuan, L. Development of γδ T cell subset responses in gnotobiotic pigs infected with human rotaviruses and colonized with probiotic lactobacilli. Vet. Immunol. Immunopathol. 2011, 141, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarlet, M.; Crawford, S.E.; Barone, C.; Bertolotti-Ciarlet, A.; Ramig, R.F.; Estes, M.K.; Conner, M.E. Subunit rotavirus vaccine administered parenterally to rabbits induces active protective immunity. J. Virol. 1998, 72, 9233–9246. [Google Scholar] [PubMed]
- Preidis, G.A.; Saulnier, D.M.; Blutt, S.E.; Mistretta, T.-A.; Riehle, K.P.; Major, A.M.; Venable, S.F.; Barrish, J.P.; Finegold, M.J.; Petrosino, J.F.; et al. Host response to probiotics determined by nutritional status of rotavirus-infected neonatal mice. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Ciarlet, M.; Conner, M.E.; Finegold, M.J.; Estes, M.K. Group a rotavirus infection and age-dependent diarrheal disease in rats: A new animal model to study the pathophysiology of rotavirus infection. J. Virol. 2002, 76, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Ramani, S.; Crawford, S.E.; Blutt, S.E.; Estes, M.K. Human organoid cultures: Transformative new tools for human virus studies. Curr. Opin. Virol. 2018, 29, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Chege, G.K.; Snodgrass, D.R.; Campbell, I.; Hart, C.; Omollo, E.; Mwenda, J.M. Prevalence of rotavirus antibodies in a non-human primate colony. Afr. J. Health Sci. 1995, 2, 296–299. [Google Scholar] [PubMed]
- Otsyula, M.; Yee, J.; Suleman, M.; Tarara, R.; Martins, J.; Woods, P.; Glass, R.; Jennings, M. Rotavirus infection in African, non-human primates. Ann. Trop. Med. Parasitol. 1996, 90, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Kojima, K.; Taniguchi, K.; Kobayashi, N. Species-specific and interspecies relatedness of NSP1 sequences in human, porcine, bovine, feline, and equine rotavirus strains. Arch. Virol. 1996, 141, 1–12. [Google Scholar] [CrossRef] [PubMed]
- El-Attar, L.; Dhaliwal, W.; Howard, C.R.; Bridger, J.C. Rotavirus cross-species pathogenicity: Molecular characterization of a bovine rotavirus pathogenic for pigs. Virology 2001, 291, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Malik, Y.S.; Sharma, K.; Dhama, K.; Ghosh, S.; Bányai, K.; Kobayashi, N.; Singh, R.K. Molecular characterization of unusual bovine rotavirus A strains having high genetic relatedness with human rotavirus: Evidence for zooanthroponotic transmission. Zoonoses Public Health 2018. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, O.; Nakagomi, T. Interspecies transmission of rotaviruses studied from the perspective of genogroup. Microbiol. Immunol. 1993, 37, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Iturriza Gómara, M.; Kang, G.; Mammen, A.; Jana, A.K.; Abraham, M.; Desselberger, U.; Brown, D.; Gray, J. Characterization of G10P[11] rotaviruses causing acute gastroenteritis in neonates and infants in Vellore, India. J. Clin. Microbiol. 2004, 42, 2541–2547. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, R.G.; Sly, D.L.; London, W.T.; Palmer, A.E.; Kalica, A.R.; Van Kirk, D.H.; Chanock, R.M.; Kapikian, A.Z. Induction of diarrhea in colostrum-deprived newborn rhesus monkeys with the human reovirus-like agent of infantile gastroenteritis. Arch. Virol. 1976, 50, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Majer, M.; Behrens, F.; Weinmann, E.; Mauler, R.; Maass, G.; Baumeister, H.G.; Luthardt, T. Diarrhea in newborn cynomologus monkeys infected with human rotavirus. Infection 1978, 6, 71–72. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Awang, A. Experimental group A rotaviral infection in cynomolgus monkeys raised on formula diet. Microbiol. Immunol. 1990, 34, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Chege, G.K.; Steele, A.D.; Hart, C.A.; Snodgrass, D.R.; Omolo, E.O.; Mwenda, J.M. Experimental infection of non-human primates with a human rotavirus isolate. Vaccine 2005, 23, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Nayak, M.K.; De, P.; Manna, B.; Dutta, S.; Bhadra, U.K.; Chawla-Sarkar, M. Species A rotaviruses isolated from hospitalized patients over 5 years of age in Kolkata, India, in 2012/13. Arch. Virol. 2018, 163, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Bruijning-Verhagen, P.; Nipshagen, M.D.; de Graaf, H.; Bonten, M.J.M. Rotavirus disease course among immunocompromised patients; 5-year observations from a tertiary care medical centre. J. Infect. 2017, 75, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Raini, S.K.; Nyangao, J.; Kombich, J.; Sang, C.; Gikonyo, J.; Ongus, J.R.; Odari, E.O. Human rotavirus group a serotypes causing gastroenteritis in children less than 5 years and HIV-infected adults in Viwandani slum, Nairobi. Ethiop. J. Health Sci. 2015, 25, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Saulsbury, F.T.; Winkelstein, J.A.; Yolken, R.H. Chronic rotavirus infection in immunodeficiency. J. Pediatr. 1980, 97, 61–65. [Google Scholar] [CrossRef]
- Glass, R.I.; Parashar, U.; Patel, M.; Gentsch, J.; Jiang, B. Rotavirus vaccines: Successes and challenges. J. Infect. 2014, 68 (Suppl. S1), S9–S18. [Google Scholar] [CrossRef] [PubMed]
- Amado, L.A.; Marchevsky, R.S.; de Paula, V.S.; Hooper, C.; Freire, M.D.S.; Gaspar, A.M.C.; Pinto, M.A. Experimental hepatitis A virus (HAV) infection in cynomolgus monkeys (Macaca fascicularis): Evidence of active extrahepatic site of HAV replication. Int. J. Exp. Pathol. 2010, 91, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.R.; Yee, J.; Barry, P.; Spinner, A.; Roberts, J.A.; Cabello, P.H.; Leite, J.P.; Lerche, N.W. Prevalence of antibodies to selected viruses in a long-term closed breeding colony of rhesus macaques (Macaca mulatta) in Brazil. Am. J. Primatol. 2003, 59, 123–128. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, L.G.; Marchevsky, R.S.; dos Santos, D.R.L.; de Oliveira, J.M.; de Paula, V.S.; Lopes, L.M.; Van der Poel, W.H.M.; González, J.E.; Munné, M.S.; Moran, J.; et al. Infection by Brazilian and Dutch swine hepatitis E virus strains induces haematological changes in Macaca fascicularis. BMC Infect. Dis. 2013, 13, 495. [Google Scholar] [CrossRef] [PubMed]
- Gardinali, N.R.; Guimarães, J.R.; Melgaço, J.G.; Kevorkian, Y.B.; de Oliveira Bottino, F.; Vieira, Y.R.; da Silva, A.C.; Pinto, D.P.; da Fonseca, L.B.; Vilhena, L.S.; et al. Cynomolgus monkeys are successfully and persistently infected with hepatitis E virus genotype 3 (HEV-3) after long-term immunosuppressive therapy. PLoS ONE 2017, 12, e0174070. [Google Scholar] [CrossRef] [PubMed]
- Leon, L.A.; Marchevsky, R.S.; Gaspar, A.M.; Garcia, R.D.; Almeida, A.J.; Pelajo-Machado, M.; Castro, T.X.; Nascimento, J.P.; Brown, K.E.; Pinto, M.A. Cynomolgus monkeys (Macaca fascicularis) experimentally infected with B19V and hepatitis A virus: No evidence of the co-infection as a cause of acute liver failure. Mem. Inst. Oswaldo Cruz 2016, 111, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Espejo, R.T.; López, S.; Arias, C. Structural polypeptides of simian rotavirus SA11 and the effect of trypsin. J. Virol. 1981, 37, 156–160. [Google Scholar] [PubMed]
- Zárate, S.; Espinosa, R.; Romero, P.; Méndez, E.; Arias, C.F.; López, S. The VP5 domain of VP4 can mediate attachment of rotaviruses to cells. J. Virol. 2000, 74, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Patton, J.T.; McDonald, S.M. Culturing, storage, and quantification of rotaviruses. Curr. Protoc. Microbiol. 2009. [Google Scholar] [CrossRef]
- Rosso, M.C.; Badino, P.; Ferrero, G.; Costa, R.; Cordero, F.; Steidler, S. Biologic Data of Cynomolgus Monkeys Maintained under Laboratory Conditions. PLoS ONE 2016, 11, e0157003. [Google Scholar] [CrossRef] [PubMed]
- Iturriza Gómara, M.; Wong, C.; Blome, S.; Desselberger, U.; Gray, J. Molecular characterization of VP6 genes of human rotavirus isolates: Correlation of genogroups with subgroups and evidence of independent segregation. J. Virol. 2002, 76, 6596–6601. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.Q.; Halkosalo, A.; Salminen, M.; Szakal, E.D.; Puustinen, L.; Vesikari, T. One-step quantitative RT-PCR for the detection of rotavirus in acute gastroenteritis. J. Virol. Methods 2008, 153, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Fumian, T.M.; Victoria, M.; Vieira, C.B.; Fioretti, J.M.; Rocha, M.S.; Prado, T.; Guimarães, F.R.; da Gama, N.P.; de Oliveira, J.M.; Mendes, A.C.O.; et al. Enteric viruses’ dissemination in a private reserve of natural heritage. Lett. Appl. Microbiol. 2018, 66, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.M.; Kerin, T.; Hull, J.; McCaustland, K.; Gentsch, J. Enhancement of detection and quantification of rotavirus in stool using a modified real-time RT-PCR assay. J. Med. Virol. 2008, 80, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Montero, H.; Rojas, M.; Arias, C.F.; López, S. Rotavirus infection induces the phosphorylation of eIF2α but prevents the formation of stress granules. J. Virol. 2008, 82, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- McNeal, M.M.; Sestak, K.; Choi, A.H.-C.; Basu, M.; Cole, M.J.; Aye, P.P.; Bohm, R.P.; Ward, R.L. Development of a rotavirus-shedding model in rhesus macaques, using a homologous wild-type rotavirus of a new P genotype. J. Virol. 2005, 79, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Weber, S.G. Rotavirus infection in adults. Lancet Infect. Dis. 2004, 4, 91–99. [Google Scholar] [CrossRef]
- Pickering, L.K.; Bartlett, A.V.; Reves, R.R.; Morrow, A. Asymptomatic excretion of rotavirus before and after rotavirus diarrhea in children in day care centers. J. Pediatr. 1988, 112, 361–365. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Sarkar, R.; Menon, V.K.; Babji, S.; Paul, A.; Rajendran, P.; Sowmyanarayanan, T.V.; Moses, P.D.; Iturriza-Gomara, M.; Gray, J.J.; et al. Rotavirus shedding in symptomatic and asymptomatic children using reverse transcription-quantitative PCR. J. Med. Virol. 2013, 85, 1661–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollet, J.J.; Ericsson, C.D.; Gibson, G.; Pickering, L.K.; DuPont, H.L.; Kohl, S.; Conklin, R.H. Human rotavirus in an adult population with travelers’ diarrhea and its relationship to the location of food consumption. J. Med. Virol. 1979, 4, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Blutt, S.E.; Kirkwood, C.D.; Parreño, V.; Warfield, K.L.; Ciarlet, M.; Estes, M.K.; Bok, K.; Bishop, R.F.; Conner, M.E. Rotavirus antigenaemia and viraemia: A common event? Lancet Lond. Engl. 2003, 362, 1445–1449. [Google Scholar] [CrossRef]
- Blutt, S.E.; Matson, D.O.; Crawford, S.E.; Staat, M.A.; Azimi, P.; Bennett, B.L.; Piedra, P.A.; Conner, M.E. Rotavirus antigenemia in children is associated with viremia. PLoS Med. 2007, 4, e121. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Liu, B.; Kohira, R.; Fuchigami, T.; Mugishima, H.; Izumi, H.; Kuzuya, M.; Fujii, R.; Hamano, M.; Ogura, H. Rotavirus antigenemia and genomia in children with rotavirus gastroenteritis. Jpn. J. Infect. Dis. 2010, 63, 83–86. [Google Scholar] [PubMed]
- Ahmed, K.; Bozdayi, G.; Mitui, M.T.; Ahmed, S.; Kabir, L.; Buket, D.; Bostanci, I.; Nishizono, A. Circulating rotaviral RNA in children with rotavirus antigenemia. J. Negat. Results Biomed. 2013, 12, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemming, M.; Huhti, L.; Räsänen, S.; Salminen, M.; Vesikari, T. Rotavirus antigenemia in children is associated with more severe clinical manifestations of acute gastroenteritis. Pediatr. Infect. Dis. J. 2014, 33, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Gilger, M.A.; Matson, D.O.; Conner, M.E.; Rosenblatt, H.M.; Finegold, M.J.; Estes, M.K. Extraintestinal rotavirus infections in children with immunodeficiency. J. Pediatr. 1992, 120, 912–917. [Google Scholar] [CrossRef]
- Nishimura, S.; Ushijima, H.; Nishimura, S.; Shiraishi, H.; Kanazawa, C.; Abe, T.; Kaneko, K.; Fukuyama, Y. Detection of rotavirus in cerebrospinal fluid and blood of patients with convulsions and gastroenteritis by means of the reverse transcription polymerase chain reaction. Brain Dev. 1993, 15, 457–459. [Google Scholar] [CrossRef]
- Ushijima, H.; Xin, K.Q.; Nishimura, S.; Morikawa, S.; Abe, T. Detection and sequencing of rotavirus VP7 gene from human materials (stools, sera, cerebrospinal fluids, and throat swabs) by reverse transcription and PCR. J. Clin. Microbiol. 1994, 32, 2893–2897. [Google Scholar] [PubMed]
- Morrison, C.; Gilson, T.; Nuovo, G.J. Histologic distribution of fatal rotaviral infection: An immunohistochemical and reverse transcriptase in situ polymerase chain reaction analysis. Hum. Pathol. 2001, 32, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; Fenaux, M.; Sharma, S.; Malik, J.; Subodh, S.; Bhatnagar, S.; Greenberg, H.; Glass, R.I.; Gentsch, J.; Bhan, M.K. Quantitative evaluation of rotaviral antigenemia in children with acute rotaviral diarrhea. J. Infect. Dis. 2006, 194, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Dung, T.T.N.; Phat, V.V.; Nga, T.V.T.; My, P.V.T.; Duy, P.T.; Campbell, J.I.; Thuy, C.T.; Hoang, N.V.M.; Van Minh, P.; Le Phuc, H.; et al. The validation and utility of a quantitative one-step multiplex RT real-time PCR targeting rotavirus A and norovirus. J. Virol. Methods 2013, 187, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-Q.; Li, D.-D.; Duan, Z.-J. Research progress of real-time quantitative PCR method for group A rotavirus detection. Bing Xue Bao Chin. J. Virol. 2013, 29, 651–654. [Google Scholar]
- Gautam, R.; Lyde, F.; Esona, M.D.; Quaye, O.; Bowen, M.D. Comparison of PremierTM Rotaclone®, ProSpecTTM, and RIDASCREEN® rotavirus enzyme immunoassay kits for detection of rotavirus antigen in stool specimens. J. Clin. Virol. 2013, 58, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Wilde, J.; Yolken, R.; Willoughby, R.; Eiden, J. Improved detection of rotavirus shedding by polymerase chain reaction. Lancet Lond. Engl. 1991, 337, 323–326. [Google Scholar] [CrossRef]
- Barnes, G.L.; Callaghan, S.L.; Kirkwood, C.D.; Bogdanovic-Sakran, N.; Johnston, L.J.; Bishop, R.F. Excretion of serotype G1 rotavirus strains by asymptomatic staff: A possible source of nosocomial infection. J. Pediatr. 2003, 142, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Del Castillo, J.R.; Ludert, J.E.; Sanchez, A.; Ruiz, M.C.; Michelangeli, F.; Liprandi, F. Rotavirus infection alters Na+ and K+ homeostasis in MA-104 cells. J. Gen. Virol. 1991, 72 Pt 3, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.F.; Ruiz, M.C.; Chemello, M.E.; Michelangeli, F. Characterization of a membrane calcium pathway induced by rotavirus infection in cultured cells. J. Virol. 1999, 73, 2481–2490. [Google Scholar] [PubMed]
- Lundgren, O.; Svensson, L. Pathogenesis of rotavirus diarrhea. Microbes Infect. 2001, 3, 1145–1156. [Google Scholar] [CrossRef]
- Sack, D.A.; Chowdhury, A.M.; Eusof, A.; Ali, M.A.; Merson, M.H.; Islam, S.; Black, R.E.; Brown, K.H. Oral hydration rotavirus diarrhoea: A double blind comparison of sucrose with glucose electrolyte solution. Lancet Lond. Engl. 1978, 2, 280–283. [Google Scholar] [CrossRef]
- Molla, A.M.; Rahman, M.; Sarker, S.A.; Sack, D.A.; Molla, A. Stool electrolyte content and purging rates in diarrhea caused by rotavirus, enterotoxigenic E. coli, and V. cholerae in children. J. Pediatr. 1981, 98, 835–838. [Google Scholar] [CrossRef]
- Holloway, G.; Coulson, B.S. Innate cellular responses to rotavirus infection. J. Gen. Virol. 2013, 94, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.A.; Yuan, L.; Rosen, B.I.; Tô, T.L.; Saif, L.J. Development of mucosal and systemic lymphoproliferative responses and protective immunity to human group A rotaviruses in a gnotobiotic pig model. Clin. Diagn. Lab. Immunol. 1996, 3, 342–350. [Google Scholar] [PubMed]
- Tabin, R.; Nusslé, D. Rotavirus enteritis in children. Helv. Paediatr. Acta. Suppl. 1980, 1–28. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Placebo Group Monkeys | Inoculated Group Monkeys | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| X11 | V7 | AB7 * | AA7 | Z7 | X9 | V11 | U13 | T7 * | |
| Age | 5 y 8 m | 6 y 10 m | 2 y 8 m | 3 y 5 m | 4 y 6 m | 5 y 9 m | 6 y 8 m | 7 y 2 m | 8 y 9 m |
| Weight (kg) | 6.29 | 6.72 | 2.43 | 2.35 | 3.98 | 4.66 | 6.70 | 5.83 | 5.19 |
| DPI | Faeces (RNA copies/mg) | ||||||||
| 0 | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) |
| 1 | (-) | (-) | (-) | (-) | (-) | (-) | (-) | 6.137 × 103 | 5.709 × 103 |
| 2 | (-) | (-) | ††† 3.0 × 103 | 5.4 × 103 | 5.691 × 103 | (-) | 6.5 × 106 | 4.646 × 103 | 1.749 × 103 |
| 3 | (-) | (-) | ††† (-) | 1.041 × 104 | 5.211 × 103 | 3.531 × 103 | 3.6 × 103 | 4.594 × 103 | 8.229 × 103 |
| 4 | (-) | (-) | 4.714 × 103 | 3.669 × 103 | (-) | 4.269 × 103 | 1.068 × 104 | (-) | (-) |
| 5 | (-) | (-) | 3.686 × 103 | (-) | (-) | (-) | (-) | (-) | (-) |
| 6 | (-) | (-) | 3.12 × 103 | 1.174 × 104 | 4.217 × 103 | 2.160 × 103 | 9.909 × 103 | 8.674 × 103 | 4.251 × 103 |
| 7 | (-) | (-) | 2.451 × 103 | 4.149 × 103 | 1.01 × 104 | (-) | 1.239 × 104 | (-) | †† 1.903 × 103 |
| 8 | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) | †† (-) |
| 9 | (-) | (-) | 3.497 × 103 | 6.703 × 103 | (-) | 3.36 × 103 | (-) | (-) | † (-) |
| 10 | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) | 3.051 × 103 |
| DPI | Sera (RNA copies/mL) | ||||||||
| 0 | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) |
| 1 | (-) | (-) | (-) | (-) | 1.455 × 103 | 1.589 × 103 | (-) | 2.253 × 103 | (-) |
| 3 | (-) | (-) | 2.157 × 103 | (-) | 9.549 × 102 | (-) | 9.206 × 102 | (-) | (-) |
| 7 | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) | (-) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bentes, G.A.; Guimarães, J.R.; Volotão, E.D.M.; Fialho, A.M.; Hooper, C.; Ganime, A.C.; Gardinali, N.R.; Lanzarini, N.M.; Da Silva, A.D.S.; Pitcovski, J.; et al. Cynomolgus Monkeys (Macaca fascicularis) as an Experimental Infection Model for Human Group A Rotavirus. Viruses 2018, 10, 355. https://0-doi-org.brum.beds.ac.uk/10.3390/v10070355
Bentes GA, Guimarães JR, Volotão EDM, Fialho AM, Hooper C, Ganime AC, Gardinali NR, Lanzarini NM, Da Silva ADS, Pitcovski J, et al. Cynomolgus Monkeys (Macaca fascicularis) as an Experimental Infection Model for Human Group A Rotavirus. Viruses. 2018; 10(7):355. https://0-doi-org.brum.beds.ac.uk/10.3390/v10070355
Chicago/Turabian StyleBentes, Gentil Arthur, Juliana Rodrigues Guimarães, Eduardo De Mello Volotão, Alexandre Madi Fialho, Cleber Hooper, Ana Carolina Ganime, Noemi Rovaris Gardinali, Natália Maria Lanzarini, Alexandre Dos Santos Da Silva, Jacob Pitcovski, and et al. 2018. "Cynomolgus Monkeys (Macaca fascicularis) as an Experimental Infection Model for Human Group A Rotavirus" Viruses 10, no. 7: 355. https://0-doi-org.brum.beds.ac.uk/10.3390/v10070355