Ginsenoside Rg1 Suppresses Type 2 PRRSV Infection via NF-κB Signaling Pathway In Vitro, and Provides Partial Protection against HP-PRRSV in Piglet

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Antibodies, Chemicals and Reagents
2.3. Quantitative Real-Time PCR
2.4. Cell Proliferation and Cytotoxicity Assay
2.5. Antiviral Activity Assay
2.6. Immunofluorescence Staining
2.7. Western Blotting
2.8. Rg1 Treatment on PRRSV Life Cycle Assay
2.9. Animal Experiment
2.10. Clinical Performance and Gross Lesions of Lung
2.11. Ethics Statement
2.12. Statistical Analysis
3. Results
3.1. Ginsenoside Rg1 Treatment Supressed PRRSV Replication in Marc-145 Cells and PAMs
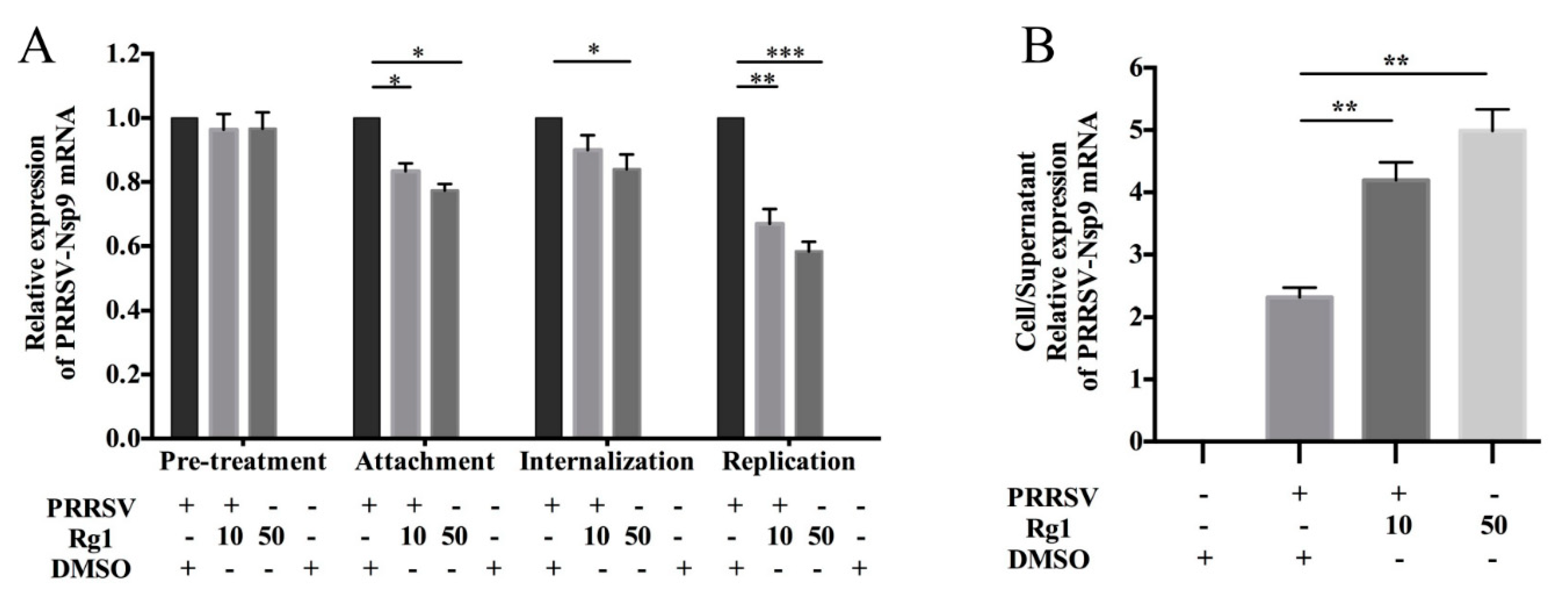
3.2. Rg1 Treatment Affect PRRSV Attachment, Replication and Release in Marc-145 Cells
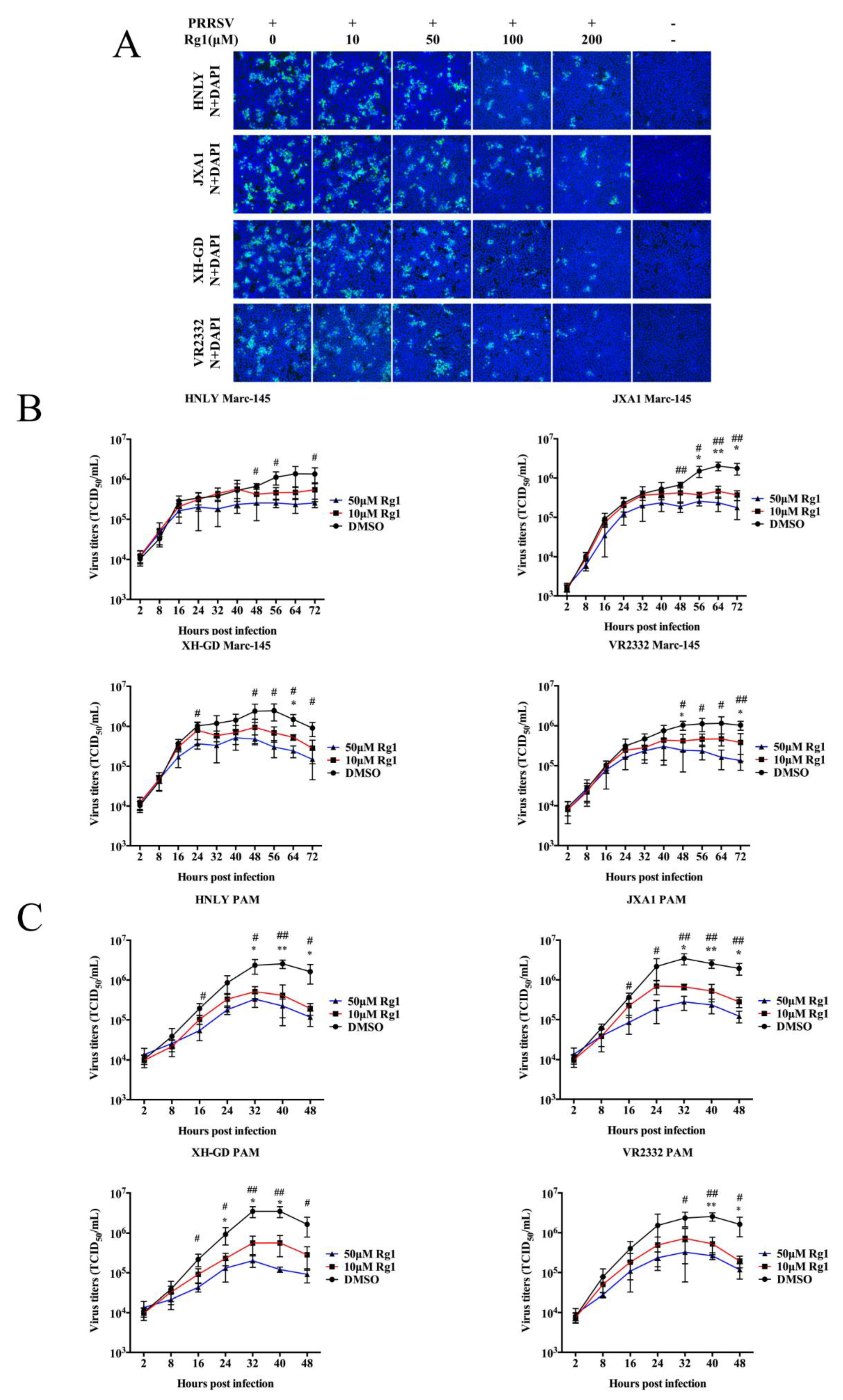
3.3. The Anti-PRRSV Ability of Rg1 Treatment Was Effective in Marc-145 Cells and PAMs Infected with the HP-PRRSV, NADC30-Like and Classical Strains
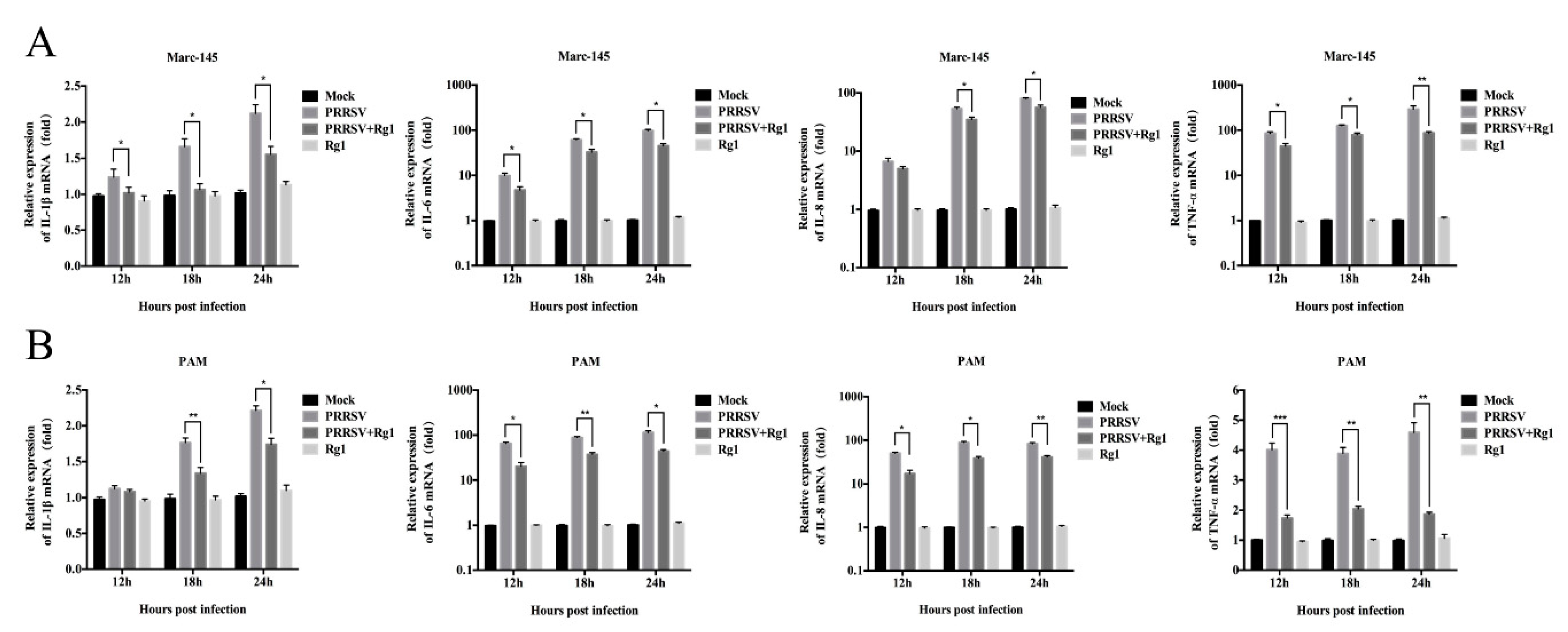
3.4. Rg1 Treatment Significantly Reduced the Pro-Inflammatory Cytokine mRNA Levels Induced by PRRSV Infection in Both Marc-145 Cells and PAMs
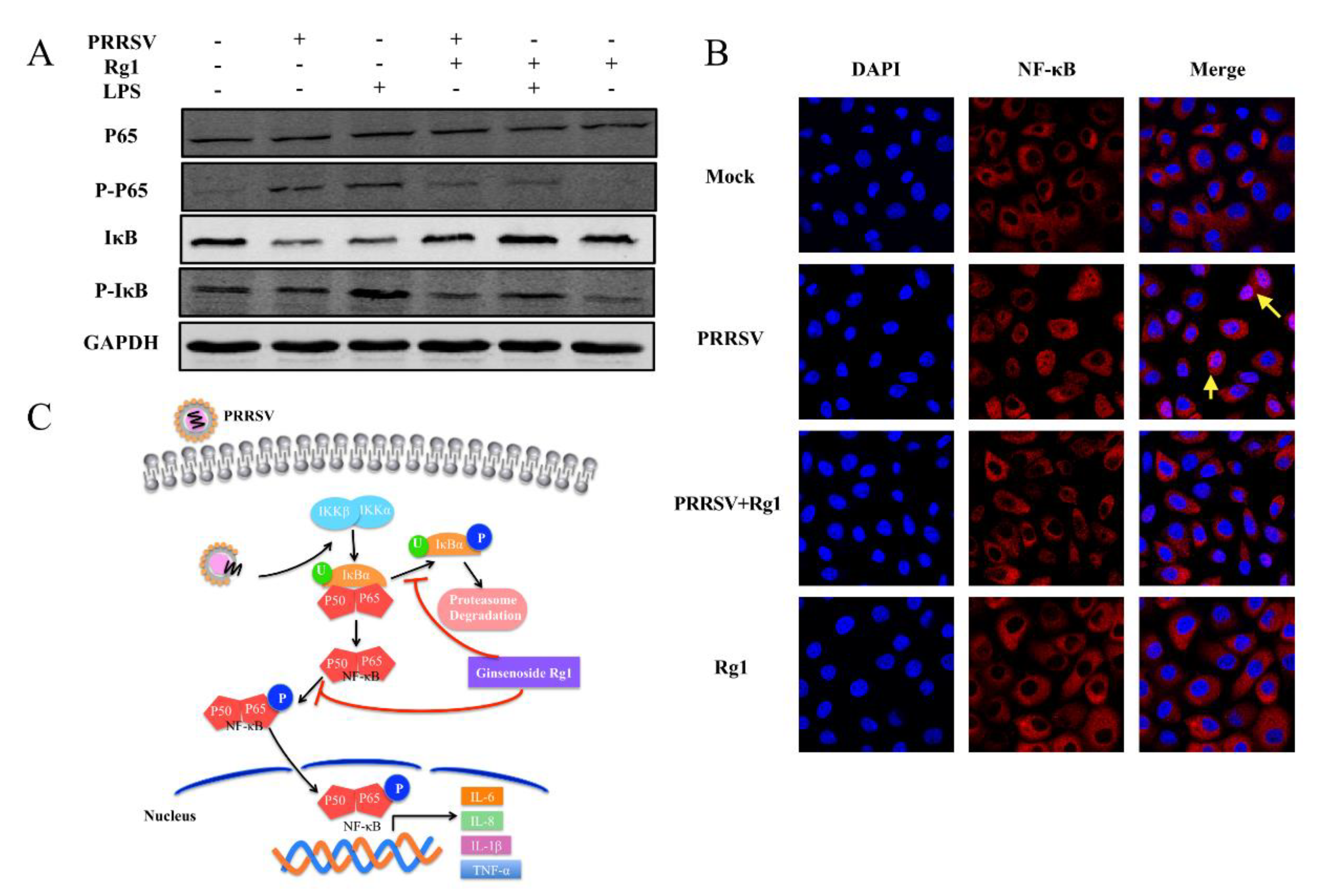
3.5. PRRSV-Infection Triggered NF-κB Activation Was Inhibited by Rg1 Treatment in Marc-145 Cells
3.6. Rg1 Treatment Exhibits Antiviral Activity in Piglets
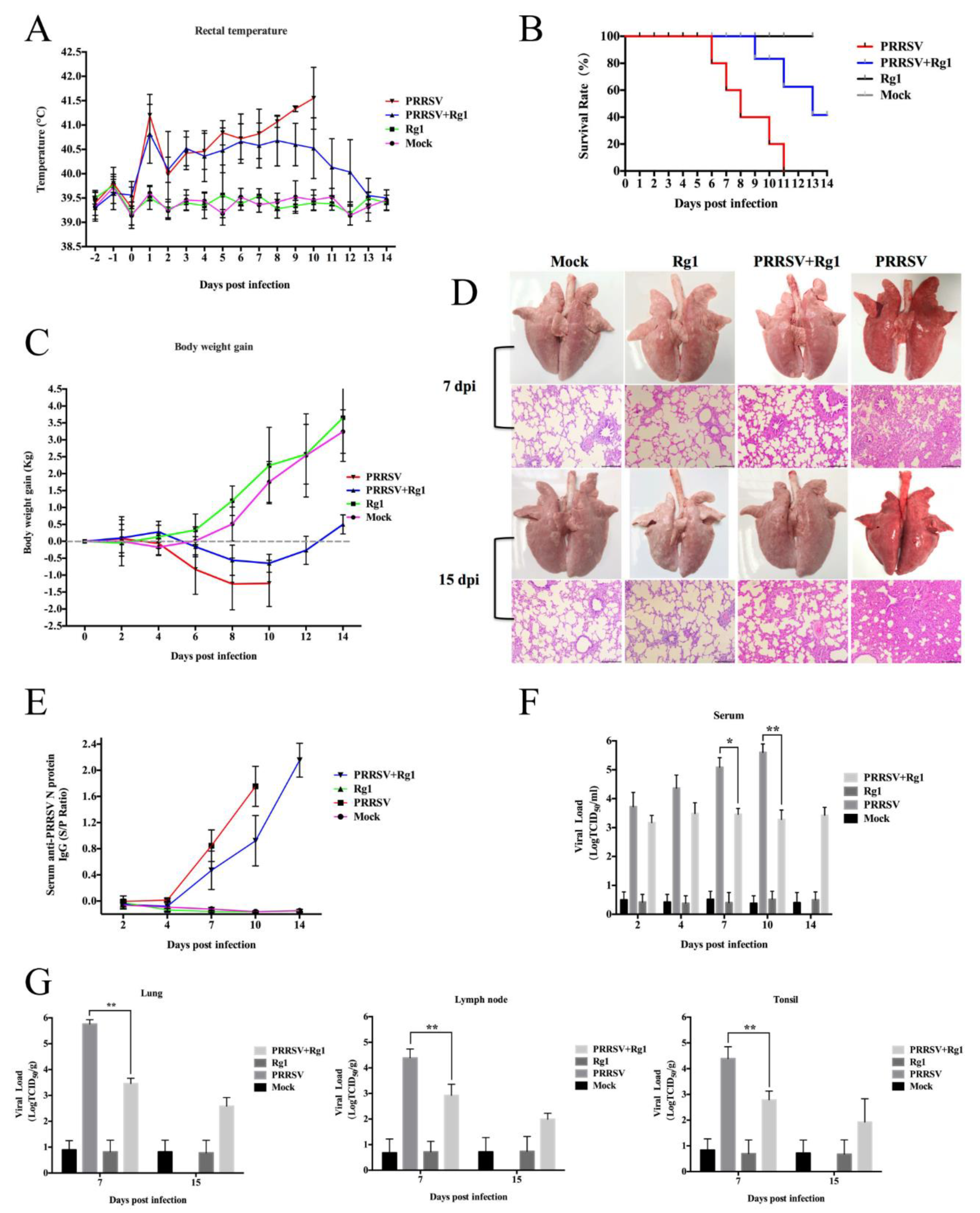
3.6.1. Clinical Signs and Mortality
3.6.2. Pathological Examination
3.6.3. Viremia and Tissue Viral Load
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Music, N.; Gagnon, C.A. The role of porcine reproductive and respiratory syndrome (PRRS) virus structural and non-structural proteins in virus pathogenesis. Anim. Health Res. Rev. 2010, 11, 135–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, C.J.; Murtaugh, M.P.; Faaberg, K.S. Porcine reproductive and respiratory syndrome virus comparison: Divergent evolution on two continents. J. Virol. 1999, 73, 270–280. [Google Scholar] [PubMed]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lemey, P.; Singh Brar, M.; Suchard, M.A.; Murtaugh, M.P.; Carman, S.; D’Allaire, S.; Delisle, B.; Lambert, M.E.; Gagnon, C.A.; et al. The spread of type 2 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) in North America: A phylogeographic approach. Virology 2013, 447, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Ait-Ali, T.; Wilson, A.D.; Westcott, D.G.; Clapperton, M.; Waterfall, M.; Mellencamp, M.A.; Drew, T.W.; Bishop, S.C.; Archibald, A.L. Innate immune responses to replication of porcine reproductive and respiratory syndrome virus in isolated Swine alveolar macrophages. Viral Immunol. 2007, 20, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.K.; Li, Q.; Yu, Z.Q.; Han, X.L.; Wei, Y.F.; Ji, C.H.; Lu, G.; Ma, C.Q.; Zhang, G.H.; Wang, H. Emergence of novel recombination lineage 3 of porcine reproductive and respiratory syndrome viruses in Southern China. Transbound. Emerg. Dis. 2019, 66, 578–587. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, J.; Bai, X.; Ji, G.; Yan, H.; Li, Y.; Wang, Y.; Tan, F.; Xiao, Y.; Li, X.; et al. Pathogenicity comparison between highly pathogenic and NADC30-like porcine reproductive and respiratory syndrome virus. Arch. Virol. 2016, 161, 2257–2261. [Google Scholar] [CrossRef]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Tong, G.Z.; Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; An, T.Q.; Qiu, H.J. Highly pathogenic porcine reproductive and respiratory syndrome, China. Emerg. Infect. Dis. 2007, 13, 1434–1436. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Z.; Ding, Y.; Ge, X.; Guo, X.; Yang, H. NADC30-like Strain of Porcine Reproductive and Respiratory Syndrome Virus, China. Emerg. Infect. Dis. 2015, 21, 2256–2257. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Gao, L.; Si, J.; Sun, Y.; Liu, J.; Cao, L.; Feng, W.H. Inhibition of porcine reproductive and respiratory syndrome virus replication by flavaspidic acid AB. Antivir. Res. 2013, 97, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Duan, E.; Wang, D.; Fang, L.; Ma, J.; Luo, J.; Chen, H.; Li, K.; Xiao, S. Suppression of porcine reproductive and respiratory syndrome virus proliferation by glycyrrhizin. Antivir. Res. 2015, 120, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Du, T.; Long, F.; Yang, X.; Sun, Y.; Duan, M.; Zhang, G.; Liu, Y.; Zhou, E.M.; Chen, W.; et al. Platycodin D Suppresses Type 2 Porcine Reproductive and Respiratory Syndrome Virus in Primary and Established Cell Lines. Viruses 2018, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Im, D.S.; Nah, S.Y. Yin and Yang of ginseng pharmacology: Ginsenosides vs gintonin. Acta Pharm. Sin. 2013, 34, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.F.; Yu, H.J.; Liu, Z.; Zhang, D.F.; Zhou, Q.J.; Zhang, H.L.; Du, A.F. Ginsenoside Rg1 enhances immune response induced by recombinant Toxoplasma gondii SAG1 antigen. Vet. Parasitol. 2011, 179, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Yuan, L.; Zhang, L.; Hu, S. Ginsenosides Rg1 and Re act as adjuvant via TLR4 signaling pathway. Vaccine 2012, 30, 4106–4112. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Song, X.; Hu, S. Ginsenoside Rg1 and aluminum hydroxide synergistically promote immune responses to ovalbumin in BALB/c mice. Clin. Vaccine Immunol. 2008, 15, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Kou, J.P.; Yu, B.Y. Ginsenoside Rg1 protects against hydrogen peroxide-induced cell death in PC12 cells via inhibiting NF-kappaB activation. Neurochem. Int. 2011, 58, 119–125. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Rong, L.; Qiao, J.O. Antiinflammatory effects of Panax notoginseng saponins ameliorate acute lung injury induced by oleic acid and lipopolysaccharide in rats. Mol. Med. Rep. 2014, 10, 1400–1408. [Google Scholar] [CrossRef]
- Chen, Y.; He, S.; Sun, L.; Luo, Y.; Sun, Y.; Xie, J.; Zhou, P.; Su, S.; Zhang, G. Genetic variation, pathogenicity, and immunogenicity of highly pathogenic porcine reproductive and respiratory syndrome virus strain XH-GD at different passage levels. Arch. Virol. 2016, 161, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.C.; Du, J.; But, P.P.; Deng, X.L.; Zhang, Y.W.; Ooi, V.E.; Xu, H.X.; Lee, S.H.; Lee, S.F. Antiviral Chinese medicinal herbs against respiratory syncytial virus. J. Ethnopharmacol. 2002, 79, 205–211. [Google Scholar] [CrossRef]
- Xie, J.X.; Cui, T.T.; Cui, J.; Chen, Y.; Zhang, M.Z.; Zhou, P.; Deng, S.C.; Su, S.; Zhang, G.H. Epidemiological and evolutionary characteristics of the PRRSV in Southern China from 2010 to 2013. Microb. Pathog. 2014, 75, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Y.F.; Lu, Q.; Bai, J.; Wang, X.Y.; Jiang, P. A new porcine reproductive and respiratory syndrome virus strain with highly conserved molecular characteristics in its parental and attenuated strains. Virus Genes 2014, 49, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Gao, J.C.; Xiong, J.Y.; Guo, J.C.; Yang, Y.B.; Jiang, C.G.; Tang, Y.D.; Tian, Z.J.; Cai, X.H.; Tong, G.Z.; et al. Two Residues in NSP9 Contribute to the Enhanced Replication and Pathogenicity of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2018, 92, e02209-17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liang, Y.; Han, J.; Burkhart, K.M.; Vaughn, E.M.; Roof, M.B.; Faaberg, K.S. Attenuation of porcine reproductive and respiratory syndrome virus strain NM184 using chimeric construction with vaccine sequence. Virology 2008, 371, 418–429. [Google Scholar] [CrossRef]
- Halbur, P.G.; Paul, P.S.; Meng, X.J.; Lum, M.A.; Andrews, J.J.; Rathje, J.A. Comparative pathogenicity of nine US porcine reproductive and respiratory syndrome virus (PRRSV) isolates in a five-week-old cesarean-derived, colostrum-deprived pig model. J. Vet. Diagn. Investig. 1996, 8, 11–20. [Google Scholar] [CrossRef]
- Nauwynck, H.J.; Duan, X.; Favoreel, H.W.; Van Oostveldt, P.; Pensaert, M.B. Entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages via receptor-mediated endocytosis. J. Gen. Virol. 1999, 80 Pt 2, 297–305. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef]
- Wang, D.; Cao, L.; Xu, Z.; Fang, L.; Zhong, Y.; Chen, Q.; Luo, R.; Chen, H.; Li, K.; Xiao, S. MiR-125b reduces porcine reproductive and respiratory syndrome virus replication by negatively regulating the NF-kappaB pathway. PLoS ONE 2013, 8, e55838. [Google Scholar]
- Chen, Y.; Yu, Z.Q.; Yi, H.Y.; Wei, Y.F.; Han, X.L.; Li, Q.; Ji, C.H.; Huang, J.M.; Deng, Q.W.; Liu, Y.X.; et al. The phosphorylation of the N protein could affect PRRSV virulence in vivo. Vet. Microbiol. 2019, 231, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; Christianson, W.T.; Morrison, R.B.; Gorcyca, D.; Chladek, D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 1992, 4, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kleiboeker, S.B. Porcine arterivirus activates the NF-kappaB pathway through IkappaB degradation. Virology 2005, 342, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Fritz, E.R.; Reecy, J.M.; Kuhar, D.; Prucnal, E.; Molina, R.; Christopher-Hennings, J.; Zimmerman, J.; Rowland, R.R. Interleukin-8, interleukin-1beta, and interferon-gamma levels are linked to PRRS virus clearance. Viral. Immunol. 2010, 23, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Thanawongnuwech, R.; Thacker, B.; Halbur, P.; Thacker, E.L. Increased production of proinflammatory cytokines following infection with porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae. Clin. Diagn. Lab Immunol. 2004, 11, 901–908. [Google Scholar] [CrossRef]
- Bao, S.; Zou, Y.; Wang, B.; Li, Y.; Zhu, J.; Luo, Y.; Li, J. Ginsenoside Rg1 improves lipopolysaccharide-induced acute lung injury by inhibiting inflammatory responses and modulating infiltration of M2 macrophages. Int. Immunopharm. 2015, 28, 429–434. [Google Scholar] [CrossRef]
- Cao, L.; Zou, Y.; Zhu, J.; Fan, X.; Li, J. Ginsenoside Rg1 attenuates concanavalin A-induced hepatitis in mice through inhibition of cytokine secretion and lymphocyte infiltration. Mol. Cell. Biochem. 2013, 380, 203–210. [Google Scholar] [CrossRef]
- Ning, C.; Gao, X.; Wang, C.; Huo, X.; Liu, Z.; Sun, H.; Yang, X.; Sun, P.; Ma, X.; Meng, Q.; et al. Protective effects of ginsenoside Rg1 against lipopolysaccharide/d-galactosamine-induced acute liver injury in mice through inhibiting toll-like receptor 4 signaling pathway. Int. Immunopharm. 2018, 61, 266–276. [Google Scholar] [CrossRef]
- Vanderheijden, N.; Delputte, P.L.; Favoreel, H.W.; Vandekerckhove, J.; Van Damme, J.; van Woensel, P.A.; Nauwynck, H.J. Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages. J. Virol. 2003, 77, 8207–8215. [Google Scholar] [CrossRef]
- Van Breedam, W.; Delputte, P.L.; Van Gorp, H.; Misinzo, G.; Vanderheijden, N.; Duan, X.; Nauwynck, H.J. Porcine reproductive and respiratory syndrome virus entry into the porcine macrophage. J. Gen. Virol. 2010, 91 Pt 7, 1659–1667. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Dudziak, D.; Dirmeier, U.; Hobom, G.; Riedel, A.; Schlee, M.; Staudt, L.M.; Rosenwald, A.; Behrends, U.; Bornkamm, G.W.; et al. Active NF-kappaB signalling is a prerequisite for influenza virus infection. J. Gen. Virol. 2004, 85 Pt 8, 2347–2356. [Google Scholar] [CrossRef]
- Williams, S.A.; Kwon, H.; Chen, L.F.; Greene, W.C. Sustained induction of NF-kappa B is required for efficient expression of latent human immunodeficiency virus type 1. J. Virol. 2007, 81, 6043–6056. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Zhang, X.Y.; Xu, L.H.; Ouyang, D.Y.; Liu, K.P.; Pan, H.; He, J.; He, X.H. Ginsenoside Rg1 regulates innate immune responses in macrophages through differentially modulating the NF-kappaB and PI3K/Akt/mTOR pathways. Int. Immunopharm. 2014, 23, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Chi, X.; Zhang, Y.; Ma, X.; Liang, S.; Wang, Y.; Hu, S.H. Ginsenoside Rg1 enhanced immune responses to infectious bursal disease vaccine in chickens with oxidative stress induced by cyclophosphamide. Poult. Sci. 2018, 97, 2698–2707. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Yuan, Q.; Cui, Q.; Liu, C.; Zhou, Z.; Zhao, H.; Dun, Y.; Wang, T.; Zhang, C. Vaccine adjuvant ginsenoside Rg1 enhances immune responses against hepatitis B surface antigen in mice. Can. J. Physiol. Pharm. 2016, 94, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Lopez, O.J.; Osorio, F.A. Role of neutralizing antibodies in PRRSV protective immunity. Vet. Immunol. Immunopathol. 2004, 102, 155–163. [Google Scholar] [CrossRef]
- Charerntantanakul, W. Porcine reproductive and respiratory syndrome virus vaccines: Immunogenicity, efficacy and safety aspects. World J. Virol. 2012, 1, 23–30. [Google Scholar] [CrossRef]
- Zuckermann, F.A.; Garcia, E.A.; Luque, I.D.; Christopher-Hennings, J.; Doster, A.; Brito, M.; Osorio, F. Assessment of the efficacy of commercial porcine reproductive and respiratory syndrome virus (PRRSV) vaccines based on measurement of serologic response, frequency of gamma-IFN-producing cells and virological parameters of protection upon challenge. Vet. Microbiol. 2007, 123, 69–85. [Google Scholar] [CrossRef]
- Li, X.; Galliher-Beckley, A.; Pappan, L.; Trible, B.; Kerrigan, M.; Beck, A.; Hesse, R.; Blecha, F.; Nietfeld, J.C.; Rowland, R.R.; et al. Comparison of Host Immune Responses to Homologous and Heterologous Type II Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Challenge in Vaccinated and Unvaccinated Pigs. Biomed. Res. Int. 2014, 2014, 416727. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence (5′-3′) |
|---|---|
| PRRSV Nsp9 | F: CCTGCAATTGTCCGCTGGTTTG |
| R: GACGACAGGCCACCTCTCTTAG | |
| GAPDH | F: GCAAAGACTGAACCCACTAATT |
| R: TTGCCTCTGTTGTTACTTGGAG | |
| Marc-145 IL-6 | F: GAGGCACTGGCAGAAAAC |
| R: TGCAGGAACTGGATCAGGAC | |
| PAM IL-6 | F: CCTTCAGTCCAGTCGCCTTCTC |
| R: CATCACCTTTGGCATCTTCTTC | |
| Marc-145 IL-8 | F: AGGACAAGAGCCAGGAAG |
| R: CTGCACCTTCACACAGAGC | |
| PAM IL-8 | F: CACTGTGAAAATTCAGAAATCATTGT |
| R: CTTCACAAATACCTGCACAACC | |
| Marc-145 TNF-α | F: TCTGTCTGCTGCACTTTGGAGTG |
| R: TTGAGGGTTTGCTACAACATGG | |
| PAM TNFα | F: TGGTGGTGCCGACAGATGG |
| R: GGCTGATGGTGTGAGTGAGG | |
| Marc-145 IL-1β | F: GGAAGACAAATTGCATGG |
| R: CCCAACTGGTACATCAGC | |
| PAM IL-1β | F: ACCTGGACCTTGGTTCTCTG |
| R: CATCTGCCTGATGCTCTTG | |
| Probe JXA1 Nsp9 | ACTGCTGCCACGACTTACTGGTCACGCAGT |
| PRRSV Strain | ||||
|---|---|---|---|---|
| XH-GD | JXA1 | HNLY | VR2332 | |
| EC50 (μM) a | 75.05 ± 13.52 | 71.33 ± 13.43 | 94.21 ± 8.27 | 55.05 ± 4.535 |
| Groups | Clinical Signs Scores (±S.D.) b | Lung Lesions Scores (±S.D.) c |
|---|---|---|
| PRRSV (Challenge control) | 13.270 ± 1.89311 | 76.0 ± 18.1711 |
| PRRSV+Rg1 | 8.807 ± 2.53021 | 34.0 ± 11.4021 |
| Mock | 02 | 02 |
| Rg1 | 02 | 02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.-q.; Yi, H.-y.; Ma, J.; Wei, Y.-f.; Cai, M.-k.; Li, Q.; Qin, C.-x.; Chen, Y.-j.; Han, X.-l.; Zhong, R.-t.; et al. Ginsenoside Rg1 Suppresses Type 2 PRRSV Infection via NF-κB Signaling Pathway In Vitro, and Provides Partial Protection against HP-PRRSV in Piglet. Viruses 2019, 11, 1045. https://0-doi-org.brum.beds.ac.uk/10.3390/v11111045
Yu Z-q, Yi H-y, Ma J, Wei Y-f, Cai M-k, Li Q, Qin C-x, Chen Y-j, Han X-l, Zhong R-t, et al. Ginsenoside Rg1 Suppresses Type 2 PRRSV Infection via NF-κB Signaling Pathway In Vitro, and Provides Partial Protection against HP-PRRSV in Piglet. Viruses. 2019; 11(11):1045. https://0-doi-org.brum.beds.ac.uk/10.3390/v11111045
Chicago/Turabian StyleYu, Zhi-qing, He-you Yi, Jun Ma, Ying-fang Wei, Meng-kai Cai, Qi Li, Chen-xiao Qin, Yong-jie Chen, Xiao-liang Han, Ru-ting Zhong, and et al. 2019. "Ginsenoside Rg1 Suppresses Type 2 PRRSV Infection via NF-κB Signaling Pathway In Vitro, and Provides Partial Protection against HP-PRRSV in Piglet" Viruses 11, no. 11: 1045. https://0-doi-org.brum.beds.ac.uk/10.3390/v11111045