The Roles of Ebola Virus Soluble Glycoprotein in Replication, Pathogenesis, and Countermeasure Development


1
National Microbiology Laboratory, Public Health Agency of Canada, Winnipeg, MB R3E 3R2, Canada
2
Department of Medical Microbiology and Infectious Diseases, Max Rady College of Medicine, University of Manitoba, Winnipeg, MB R3E 0J9, Canada
3
Institute Pasteur of Shanghai, Chinese Academy of Sciences, Shanghai, China
4
Département de microbiologie-infectiologie et d’immunologie, Université Laval, Québec, QC G1V 0A6, Canada
*
Authors to whom correspondence should be addressed.
Viruses 2019, 11(11), 999; https://0-doi-org.brum.beds.ac.uk/10.3390/v11110999
Submission received: 7 October 2019
/
Revised: 28 October 2019
/
Accepted: 29 October 2019
/
Published: 31 October 2019
(This article belongs to the Special Issue Pathogenesis of Emerging Viral Infections)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Ebola virus (EBOV) is a highly lethal pathogen that has caused several outbreaks of severe hemorrhagic fever in humans since its emergence in 1976. The EBOV glycoprotein (GP1,2) is the sole viral envelope protein and a major component of immunogenicity; it is encoded by the GP gene along with two truncated versions: soluble GP (sGP) and small soluble GP (ssGP). sGP is, in fact, the primary product of the GP gene, and it is secreted in abundance during EBOV infection. Since sGP shares large portions of its sequence with GP1,2, it has been hypothesized that sGP may subvert the host immune response by inducing antibodies against sGP rather than GP1,2. Several reports have shown that sGP plays multiple roles that contribute to the complex pathogenesis of EBOV. In this review, we focus on sGP and discuss its possible roles with regards to the pathogenesis of EBOV and the development of specific antiviral drugs.
1. Introduction
Filoviruses are negative stranded, nonsegmented RNA viruses that belong to the family Filoviridae, which is further subdivided into five genera: Marburgvirus, Ebolavirus, Cuevavirus, Striavirus, and Thamnovirus [1]. The Ebolavirus genus consists of five viruses representing five distinct species: Ebola virus (EBOV; species Zaire ebolavirus), Sudan virus (SUDV; Sudan ebolavirus), Bundibugyo virus (BDBV; species Bundibugyo ebolavirus), Taï Forest virus (TAFV; species Taï Forest ebolavirus), and Reston virus (RESTV; species Reston ebolavirus). The recently described Bombali virus (BOMV) has been proposed to belong to a sixth species, tentatively called Bombali ebolavirus [1,2]. EBOV attracted global attention following a large outbreak in West Africa during 2013–2016, which led to over 10,000 deaths [3]. EBOV causes severe hemorrhagic fever in humans and non-human primates (NHPs), with a fatality rate up to 90% [3,4]. Infected patients typically develop fever, headache, vomiting and diarrhea, progressing to shock and multiorgan failure in severe cases. Hemorrhage is also observed in a subset of victims. Neurological symptoms such as meningoencephalitis, seizures, and coma have also been reported [5,6,7]. Bats are suspected to be the reservoir of EBOV, and introduction into humans is thought to occur following direct contact with bats or bat excreta or through contact with other susceptible animals, such as NHPs [3]. Direct transmission between humans is the major mode of EBOV spread because the virus can be shed in many bodily fluids, including blood, saliva, tears, urine, semen, and sweat [3].
The EBOV genome is approximately 19 kb in size, and it encodes seven structural proteins, including the nucleoprotein (NP), virion protein (VP) 35, VP40, glycoprotein (GP1,2), VP30, VP24, and the RNA-dependent RNA polymerase (L) (Figure 1A) [4,8]. NP, which binds to the viral genome, is the major component of the nucleocapsid, along with VP35 and VP24. VP30 is also a component of the nucleocapsid, in addition to its role as a transcription factor. VP40 is the matrix protein, which drives the formation of new virus particles, and L, along with VP35 as a polymerase cofactor, facilitates genome replication and transcription. GP1,2, a class I membrane protein residing in the viral envelope, serves as the major viral attachment and entry factor, although the complex mechanisms governing EBOV entry into the host cell are only partially understood [9]. Intriguingly, through a co-transcriptional editing mechanism, the GP gene of all ebolaviruses encodes two additional, nonstructural proteins known as soluble GP (sGP) and small soluble GP (ssGP) (Figure 1) [10,11,12]. Both sGP and ssGP have been proposed to play a number of different roles during infection [10,13,14,15,16,17]. This review presents our current understanding of sGP, and it describes the roles of this protein during the various phases of EBOV pathogenesis, with the view of connecting structure to function and ultimately informing the rational development of antiviral therapies.
2. EBOV GP Gene Products
The GP gene encodes three proteins, GP1,2, sGP, and ssGP, that are the product of a unique co-transcriptional editing strategy (Figure 1) [10,11,12,18]. sGP mRNA is the primary product of the GP gene, accounting for approximately 70% of transcripts [10,11]. Occasionally, however, the viral polymerase stutters during transcription at a stretch of seven uridines present in the viral genome just upstream of the sGP stop codon. This stuttering results in the addition of a non-templated adenosine residue in the nascent transcript, thus producing a frame shift that alters the position of the stop codon. The resulting longer mRNA, which accounts for approximately 25% of the transcripts derived from the GP gene, encodes the GP1,2 precursor [10,11]. The co-transcriptional addition of two adenosine residues, or the subtraction of a single adenosine, results in a different frame shift that produces a much shorter transcript, which encodes ssGP [10]. Thus, transcripts that encode sGP contain a stretch of seven adenosine residues (7A), transcripts that encode GP1,2 contain a stretch of eight adenosine residues (8A), and transcripts that encode ssGP contain a stretch of six or nine adenosine residues (6A/9A) [11,12]. Notably, while this co-transcriptional strategy is employed by all ebolaviruses and cuevaviruses, it is not employed by the marburgviruses, including Marburg virus (MARV) and Ravn virus, which consequently do not produce sGP or ssGP [19].
Following transcription, GP1,2 is produced as a precursor protein, known as pre-GP or GP0, which is then cleaved post-translationally by furin-like proteases to yield the ectodomains GP1 and GP2 (Figure 2) [9,20,21]. GP1 and GP2 then form dimers, which in turn form trimers to produce the mature and functional heterotrimeric GP1,2. As the sole viral protein expressed on the surface of the virion envelope, GP1,2 facilitates attachment to host cells by interacting with a variety of cell surface factors, including carbohydrate-binding receptors [9]. Following internalization of the virion by macropinocytosis and progression through the endocytic pathways, cleavage of the GP1,2 mucin-like domain and glycan cap by cathepsins L and B reveals the receptor binding site. Interaction between the GP1,2 receptor binding site and the filovirus receptor Niemann–Pick C1 (NPC1) in the late endosome/early lysosome, results in a conformational change in GP1,2 that drives fusion of the virion and endosomal membranes, resulting in the release of the virus genome into the cytoplasm [22]. Interestingly, proteolytic cleavage of surface-expressed GP1,2 by the cellular tumor necrosis factor α converting enzyme (TACE) removes the transmembrane anchor and liberates GP1 in complex with a truncated GP2, a product known as shed GP [23]. Shed GP has been reported to activate the Toll-like receptor (TLR) 4 signaling pathway, resulting in the expression of pro-inflammatory cytokines, maturation of macrophages and dendritic cells [24]. Shed GP was also demonstrated to increase endothelial permeability and block the activity of neutralizing antibodies [23,25].
sGP, the most abundant product from the GP gene, is initially synthesized as a precursor protein known as pre-sGP (Figure 2) [26]. Subsequent post-translational proteolytic cleavage by furin and dimerization produces mature sGP, which is secreted from cells. The byproduct of furin cleavage at amino acid position 324 in the C-terminus of sGP produces the Δ-peptide, which is also secreted from cells [26]. Although sGP shares the first 295 N-terminal amino acids with GP1,2, the structure and function are markedly different: sGP forms 110 kDa homodimers in a parallel orientation, whereas GP forms anti-parallel trimers [10,27]. Since sGP is the primary product of the GP gene and shares common residues with GP1,2, it may play a role in the replication cycle and pathogenesis of EBOV. sGP may also be an excellent biomarker for specific diagnosis of EBOV since it is secreted in abundance into the blood during the early stages of infection. Moreover, sGP constitutes a promising target for vaccines and antiviral therapeutics due to its many functions on the host immune system.
3. sGP May Substitute as a Structural Protein
sGP, which shares the same N-terminus as GP1,2, is commonly thought of as a nonstructural, secretory glycoprotein. Both sGP and GP1 share a cysteine residue at position 53, which forms a disulfide bond with cysteine 609 in GP2 [12]. Data suggest that sGP can substitute for GP1, forming a complex with GP2 to create a functional glycoprotein [13]. Indeed, vesicular stomatitis virus (VSV) pseudotyped with the sGP–GP2 complex resulted in an infectious virus. However, sGP was also shown to result in a reduction in virus titer when overexpressed with GP1,2. Together, these data suggest not only that sGP may have a novel role as a structural protein by replacing GP1, but they also imply that sGP may have a role in limiting the cytotoxicity of GP1,2. The GP1 mucin-like domain is known to have cytotoxic effects, and it has been previously reported that a recombinant EBOV encoding only GP1,2 and not sGP showed significantly increased cytotoxicity [14]. It is therefore tempting to speculate that by replacing a certain amount of GP1, sGP may decrease the overall cytotoxicity of EBOV to the host cells and, in so doing, facilitate more efficient replication and promote infectivity.
4. sGP Serves as a Virulence Factor
Previous studies have shown that a recombinant virus encoding eight uridines (8U) at the GP transcriptional editing site (producing an 8A transcript) quickly reverts to the wild type 7U (7A transcript) in guinea pigs [28]. This finding suggests that there is a strong selective pressure in vivo for sGP production and that sGP plays a critical role during EBOV infection. Moreover, passage of EBOV in Vero E6 cells quickly leads to an eight uridine genotype, whereas passage in Huh7 cells favors the seven uridine genotype [29], indicating that sGP expression may also be cell type dependent. Indeed, although recombinant EBOV devoid of sGP exhibited increased synthesis of GP1,2 and enhanced cytotoxicity [14], the virus proved less pathogenic in animals [15]. Guinea pigs infected with sGP-deficient EBOV had lower viral loads throughout the course of the experiments compared to animals infected with wild type EBOV, although both groups developed an antibody response against GP [15]. This finding suggests that sGP may play a role in the pathogenicity of EBOV, and it provides evidence supporting the idea that GP1,2 cytotoxicity may limit virus spread. Conversely, however, another study found that the absence of sGP did not influence the infectivity of a recombinant, sGP-deficient EBOV, as this virus did not show significant attenuation in guinea pigs [30]. The reason for the discrepancy between these two studies remains unclear, although it may berelated to the differences in the recombinant viruses used or differences in the outbred guinea pigs. Further studies will be required to investigate the effect of sGP on EBOV virulence in other animal models, such as NHPs, which more closely resemble humans.
Interestingly, Whitmer et al. investigated persistent infections in EBOV survivors and sequenced EBOV from semen samples [31]. In virus sequences obtained from one patient, the authors found a cytidine insertion at the GP editing site that resulted in the genome sequence encoding a full-length GP1,2 with the frequency of 34%–65%. This is the first published case from a human patient to show that selective pressure from specific tissue compartments, such as immune privileged sites, may cause mutations that decrease sGP production [31]. However, whether this insertion mutation resulted in a significant decrease in sGP production, and whether this might have contributed to patient survival or virus persistence, remains unclear.
5. sGP Alters the Immune Response
Previous studies have suggested that sGP may exhibit anti-inflammatory activities [17]. Kindzelskii et al. showed that sGP dramatically reduces the amount of CD16b receptor on human neutrophils in a dose-dependent manner [32]. They also found that sGP could induce a conformational change in this receptor, helping prevent the activation of neutrophils and stunt the innate immune response by preventing immune complexes to activate neutrophil metabolic flux [32]. Indeed, during EBOV infection, many areas of focal tissue destruction in multiple organs, including the liver and kidneys, lack leukocyte infiltration, although neutrophil aggregation can still be observed within the vascular system [17]. This may be explained by the inhibition of the transmigration process of leukocytes through the endothelium due to the anti-inflammatory effect of sGP. Wahl-Jensen et al. showed that VP40 and GP1,2 were able to activate endothelial cells and decrease barrier function—which was enhanced by the cytokine tumor necrosis factor alpha (TNF-α)—while sGP induced a recovery of endothelial barrier function following treatment with TNF-α [17]. Thus, sGP seems to play an anti-inflammatory role by protecting the integrity of the endothelium and promoting the recovery of barrier function, which may support virus replication. sGP was also shown to reduce the production of pro-inflammatory, but not anti-inflammatory, cytokines by macrophages [33]. Moreover, although sGP was shown not to affect phagocytosis, it significantly diminished the chemotaxis of activated macrophages [33]. Together, these data suggest that sGP impairs effector activities of immune cells before they get infected, thereby creating a pool of susceptible macrophages primed for infection [33]. The preservation of the phagocytic ability of macrophages also ensures that these cells can continue to take up virions and promote cell infection [33]. Indeed, the anti-inflammatory functions of sGP may provide a relatively stable environment for EBOV-infected cells and support virus replication and transmission to other cells. More studies will be needed to address the mechanism of the potential anti-inflammatory function of sGP, and whether the effect is specific for TNF-α or other components in the signaling pathway. It would also be interesting to determine whether the sGP functions observed in cell culture are retained in vivo, and whether eliminating sGP might help restore the normal function of immune cells.
6. sGP Is an Important Target for the Immune Response
sGP has been shown to act as a decoy antigen through binding specific antibodies against GP1,2, possibly resulting in antigenic subversion and contributing to systemic viral spread in the host [34]. High amounts of sGP are thought to divert the host humoral immune response away from GP1,2 and towards sGP, a phenomenon that is supported by data showing that sGP is able to efficiently compete for anti-GP1,2 antibodies from mice immunized by sGP [34]. Additionally, sGP can interfere with viral neutralization by antisera from mice immunized with GP1,2 and sGP [34]. However, when mice were immunized with GP1,2 alone (in which expression of sGP was disabled), anti-GP1,2 antibodies showed low cross-reactivity with sGP, consistent with previous studies demonstrating that antibodies generated in response to GP1,2 do not share many epitopes with sGP [35,36]. One recent study showed that a chimeric EBOV expressing BDBV GP but not sGP, displayed increased susceptibility to a monoclonal antibody isolated from a survivor of BDBV infection, further demonstrating a role for sGP in evasion of antibody neutralization [37].
Somewhat paradoxically, Liu et al. recently showed that intradermal or intramuscular immunization with vaccines containing the sGP subunit plus adjuvant can confer effective protection to mice against lethal EBOV challenge by producing antibodies that can neutralize both GP1,2 and sGP [38]. These data demonstrate that sGP elicits sGP/GP1,2 cross-reactive antibodies, and they may suggest that neutralization of sGP itself offers some therapeutic benefit. Indeed, recent analysis of a number of different sGP/GP1,2 cross-reactive antibodies demonstrated that some of these antibodies could more effectively neutralize wild type EBOV (which expresses sGP) than VSV pseudotyped with EBOV GP (which does not express sGP) [39]. This study therefore raises the intriguing possibility that certain sGP/GP1,2 cross-reactive antibodies may depend on an interaction with sGP in order to achieve their therapeutic benefit. Notably, 13C6, a monoclonal antibody that is part of the ZMapp cocktail, has been shown to bind both sGP and GP1,2 [16,40], and it is tempting to speculate that binding to sGP may be critical for this antibody’s therapeutic function. Moreover, analysis of the immune effector functions of GP1,2- and sGP-specific IgA1 antibodies isolated from human survivors revealed an association with antibody-dependent phagocytosis by neutrophils (ADNP) [41], demonstrating the important role an immune response against sGP may play during EBOV infection.
With respect to the cellular immune response, a recent study investigated the EBOV-specific T-cell memory responses in patients infected during the 2013-2016 EBOV outbreak [42]. The study revealed that 8/10 individuals had T-cell responses to both GP1,2 and sGP in their peripheral blood mononuclear cells (PBMCs), while only 1/10 responded to GP1,2 but not sGP. These data therefore suggest that the portion of GP1 shared with sGP may be more immunogenic than GP1,2 in inducing an effector T-cell response, thereby diverting the immune response away from GP1,2 [42].
7. Δ-Peptide May Have a Role in Viral Replication and Pathogenicity
Cleavage of the sGP protein by furin yields Δ-peptide, a 40 aa carboxy-terminal fragment that is released from cells more slowly and at lower amounts than sGP [26,43]. The function of the Δ-peptide is still unknown, but sequence analysis suggests it could be a candidate viroporin, capable of permeabilizing cellular membranes. Δ-peptide possesses a motif similar to one found in known viroporins, such as rotavirus NSP4 and HIV LLP-1, suggesting that this peptide may act as a lysin or cytotoxin [26]. Indeed, a recent study showed that the Δ-peptide has a high abundance of aromatic and cationic residues, which are found in many membrane-permeabilizing peptides [44]. Another study has shown that Δ-peptide is able to permeabilize mammalian cell plasma membranes at micromolar concentrations, so it may alter the permeation of ionic compounds and small molecules into the cells, contributing to viral pathology by causing disruption of cell function and cell death [45]. On the other hand, Δ-peptide may function to modulate virus entry and infection of certain cell types. When Δ-peptide-Fc domain chimeras were added externally to cells, cell viability was not affected; however, the chimeras inhibited the entry of all filoviruses, including MARV [43]. These data suggest that Δ-peptide inhibits GP1,2 interaction with cellular attachment factors, and while the implications of this function are unclear, it may help facilitate virion budding or prevent superinfection [43].
8. The Roles of ssGP in Pathogenesis Remain Unknown
ssGP is the second, nonstructural glycoprotein produced through RNA editing during ebolavirus infection, with less than 5% of GP gene transcripts thought to be specific for ssGP mRNA [46]. ssGP possesses the same 295 N-terminal amino acid sequence with GP1,2 and sGP, differing at their C-terminal sites [10]. Based on their shared sequence, it was previously postulated that ssGP may function similarly to sGP; however, this appears not to be the case. Unlike sGP, ssGP did not exhibit anti-inflammatory function without effect on the endothelial barrier restoration [10]. This may be explained by the observation that ssGP does not contain the cysteine 306 which sGP did. Moreover, Mehedi et al. investigated the binding of ssGP to peripheral blood mononuclear cells and found that it did not interact with CD16b, suggesting that ssGP does not possess the same anti-inflammatory functions as sGP [10]. Thus, the role of ssGP in ebolavirus pathogenesis remains unclear, and more studies are needed to unravel its potential as an antiviral target.
9. Perspectives on the Role of sGP as a Biomarker for Diagnosis and a Target for Antiviral Therapy
At present, the development of post-exposure treatments for Ebola virus disease is mostly focused on neutralizing antibodies against GP1,2 [47]. Indeed, one monoclonal antibody, mAb114, and two cocktails of monoclonal antibodies, ZMapp and REGN-EB3, have been evaluated in a randomized clinical trial during the ongoing EBOV outbreak in the Democratic Republic of the Congo [48]. However, as one of the most abundant proteins produced during EBOV infection, sGP may also be a promising target of antiviral therapy. Firstly, sGP may be targeted as part of antibody treatments. Since sGP can bind to GP/sGP cross-reactive antibodies, it may be necessary to use sGP-specific antibodies to diminish the binding of sGP to the treatment antibodies, thereby enhancing their therapeutic effect. Secondly, sGP has shown some effects on immune cells such as macrophages, inhibiting their ability to attack EBOV or remove virus infected cells. Inhibiting these deleterious functions of sGP may interfere with EBOV replication and reduce disease severity. Thirdly, sGP can subvert the host immune system by shifting the antibody response towards sGP-specific antibodies and away from GP-specific, neutralizing antibodies. Thus, it may be a promising therapeutic strategy to administer both sGP-specific antibodies along with sGP/GP1,2-cross-reactive antibodies to limit the decoy effect of sGP and enhance neutralization of GP1,2. Finally, sGP may act as a biomarker for diagnosis of EVD, since sGP is produced at an early stage of EBOV infection and secreted systemically in high amounts. Indeed, sGP can be detected by ELISA or similar methods, and its detection may aid in early diagnosis of infection. Overall, sGP can be considered a promising target in the treatment of EBOV infection and a key factor in EBOV pathogenesis (Figure 3).
10. Conclusions
The 2013–2016 EBOV outbreak in West Africa caused global concern, particularly in countries with endemic/imported cases [49]. There is an urgent need to design effective therapies to prevent future outbreaks. Due to its multifaceted roles in viral pathogenesis, sGP is a promising target for the diagnosis and treatment of EBOV infections. Several therapeutic antibodies targeting GP have been developed and are currently in clinical trials [47]. In addition to direct roles of GP1,2 in the virus replication cycle, sGP and its cleaved product Δ-peptide are all believed to contribute to EBOV infection and replication. However, for all reported functions of sGP, further investigations are required to definitively understand the roles of sGP in EBOV biology and determine whether it can be effectively targeted during infection.
Funding
This work was supported by Public Health Agency of Canada.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Kuhn, J.H.; Amarasinghe, G.K.; Basler, C.F.; Bavari, S.; Bukreyev, A.; Chandran, K.; Crozier, I.; Dolnik, O.; Dye, J.M.; Formenty, P.B.H.; et al. ICTV Virus Taxonomy Profile: Filoviridae. J. Gen. Virol. 2019, 100, 911–912. [Google Scholar] [CrossRef] [PubMed]
- Forbes, K.M.; Webala, P.W.; Jaaskelainen, A.J.; Abdurahman, S.; Ogola, J.; Masika, M.M.; Kivisto, I.; Alburkat, H.; Plyusnin, I.; Levanov, L.; et al. Bombali Virus in Mops condylurus Bat, Kenya. Emerg. Infect. Dis. 2019, 25, 955–957. [Google Scholar] [CrossRef] [PubMed]
- Baseler, L.; Chertow, D.S.; Johnson, K.M.; Feldmann, H.; Morens, D.M. The Pathogenesis of Ebola Virus Disease. Annu. Rev. Pathol. 2017, 12, 387–418. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, H.; Sanchez, A.; Geisbert, T.W. Fields Virology. In Filoviridae: Marburg and Ebola Viruses; Wolters Kluwer: Philadelphia, PA, USA, 2013; Chapter 32; pp. 923–956. [Google Scholar]
- Chertow, D.S.; Kleine, C.; Edwards, J.K.; Scaini, R.; Giuliani, R.; Sprecher, A. Ebola virus disease in West Africa—Clinical manifestations and management. N. Engl. J. Med. 2014, 371, 2054–2057. [Google Scholar] [CrossRef]
- Bwaka, M.A.; Bonnet, M.J.; Calain, P.; Colebunders, R.; De Roo, A.; Guimard, Y.; Katwiki, K.R.; Kibadi, K.; Kipasa, M.A.; Kuvula, K.J.; et al. Ebola hemorrhagic fever in Kikwit, Democratic Republic of the Congo: Clinical observations in 103 patients. J. Infect. Dis. 1999, 179 (Suppl. 1), S1–S7. [Google Scholar] [CrossRef]
- Sagui, E.; Janvier, F.; Baize, S.; Foissaud, V.; Koulibaly, F.; Savini, H.; Maugey, N.; Aletti, M.; Granier, H.; Carmoi, T. Severe Ebola Virus Infection With Encephalopathy: Evidence for Direct Virus Involvement. Clin. Infect. Dis. 2015, 61, 1627–1628. [Google Scholar] [CrossRef] [Green Version]
- Kirchdoerfer, R.N.; Wasserman, H.; Amarasinghe, G.K.; Saphire, E.O. Filovirus Structural Biology: The Molecules in the Machine. Curr. Top. Microbiol. Immunol. 2017, 411, 381–417. [Google Scholar]
- Davey, R.A.; Shtanko, O.; Anantpadma, M.; Sakurai, Y.; Chandran, K.; Maury, W. Mechanisms of Filovirus Entry. Curr. Top. Microbiol. Immunol. 2017, 411, 323–352. [Google Scholar]
- Mehedi, M.; Falzarano, D.; Seebach, J.; Hu, X.; Carpenter, M.S.; Schnittler, H.J.; Feldmann, H. A new Ebola virus nonstructural glycoprotein expressed through RNA editing. J. Virol. 2011, 85, 5406–5414. [Google Scholar] [CrossRef]
- Volchkov, V.E.; Becker, S.; Volchkova, V.A.; Ternovoj, V.A.; Kotov, A.N.; Netesov, S.V.; Klenk, H.D. GP mRNA of Ebola virus is edited by the Ebola virus polymerase and by T7 and vaccinia virus polymerases. Virology 1995, 214, 421–430. [Google Scholar] [CrossRef]
- Sanchez, A.; Trappier, S.G.; Mahy, B.W.; Peters, C.J.; Nichol, S.T. The virion glycoproteins of Ebola viruses are encoded in two reading frames and are expressed through transcriptional editing. Proc. Natl. Acad. Sci. USA 1996, 93, 3602–3607. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, A.; Shimojima, M.; Kawaoka, Y. sGP serves as a structural protein in Ebola virus infection. J. Infect. Dis. 2011, 204 (Suppl. 3), S897–S903. [Google Scholar] [CrossRef] [PubMed]
- Volchkov, V.E.; Volchkova, V.A.; Muhlberger, E.; Kolesnikova, L.V.; Weik, M.; Dolnik, O.; Klenk, H.D. Recovery of infectious Ebola virus from complementary DNA: RNA editing of the GP gene and viral cytotoxicity. Science 2001, 291, 1965–1969. [Google Scholar] [CrossRef] [PubMed]
- Volchkova, V.A.; Dolnik, O.; Martinez, M.J.; Reynard, O.; Volchkov, V.E. RNA Editing of the GP Gene of Ebola Virus is an Important Pathogenicity Factor. J. Infect. Dis. 2015, 212 (Suppl. 2), S226–S233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallesen, J.; Murin, C.D.; de Val, N.; Cottrell, C.A.; Hastie, K.M.; Turner, H.L.; Fusco, M.L.; Flyak, A.I.; Zeitlin, L.; Crowe, J.E., Jr.; et al. Structures of Ebola virus GP and sGP in complex with therapeutic antibodies. Nat. Microbiol. 2016, 1, 16128. [Google Scholar] [CrossRef]
- Wahl-Jensen, V.M.; Afanasieva, T.A.; Seebach, J.; Stroher, U.; Feldmann, H.; Schnittler, H.J. Effects of Ebola virus glycoproteins on endothelial cell activation and barrier function. J. Virol. 2005, 79, 10442–10450. [Google Scholar] [CrossRef]
- de La Vega, M.A.; Wong, G.; Kobinger, G.P.; Qiu, X. The multiple roles of sGP in Ebola pathogenesis. Viral. Immunol. 2015, 28, 3–9. [Google Scholar] [CrossRef]
- Anthony, S.M.; Bradfute, S.B. Filoviruses: One of These Things is (not) Like the Other. Viruses 2015, 7, 5172–5190. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.D.; Lee, J.E. The secret life of viral entry glycoproteins: Moonlighting in immune evasion. PLoS Pathog. 2013, 9, e1003258. [Google Scholar] [CrossRef] [PubMed]
- Volchkov, V.E.; Feldmann, H.; Volchkova, V.A.; Klenk, H.D. Processing of the Ebola virus glycoprotein by the proprotein convertase furin. Proc. Natl. Acad. Sci. USA 1998, 95, 5762–5767. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Qian, H.; Zhou, X.; Wu, J.; Wan, T.; Cao, P.; Huang, W.; Zhao, X.; Wang, X.; Wang, P.; et al. Structural Insights into the Niemann-Pick C1 (NPC1)-Mediated Cholesterol Transfer and Ebola Infection. Cell 2016, 165, 1467–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolnik, O.; Volchkova, V.; Garten, W.; Carbonnelle, C.; Becker, S.; Kahnt, J.; Stroher, U.; Klenk, H.D.; Volchkov, V. Ectodomain shedding of the glycoprotein GP of Ebola virus. EMBO J. 2004, 23, 2175–2184. [Google Scholar] [CrossRef] [PubMed]
- Okumura, A.; Pitha, P.M.; Yoshimura, A.; Harty, R.N. Interaction between Ebola virus glycoprotein and host toll-like receptor 4 leads to induction of proinflammatory cytokines and SOCS1. J. Virol. 2010, 84, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Escudero-Perez, B.; Volchkova, V.A.; Dolnik, O.; Lawrence, P.; Volchkov, V.E. Shed GP of Ebola virus triggers immune activation and increased vascular permeability. PLoS Pathog. 2014, 10, e1004509. [Google Scholar] [CrossRef]
- Volchkova, V.A.; Klenk, H.D.; Volchkov, V.E. Delta-peptide is the carboxy-terminal cleavage fragment of the nonstructural small glycoprotein sGP of Ebola virus. Virology 1999, 265, 164–171. [Google Scholar] [CrossRef]
- Volchkova, V.A.; Feldmann, H.; Klenk, H.D.; Volchkov, V.E. The nonstructural small glycoprotein sGP of Ebola virus is secreted as an antiparallel-orientated homodimer. Virology 1998, 250, 408–414. [Google Scholar] [CrossRef]
- Volchkova, V.A.; Dolnik, O.; Martinez, M.J.; Reynard, O.; Volchkov, V.E. Genomic RNA editing and its impact on Ebola virus adaptation during serial passages in cell culture and infection of guinea pigs. J. Infect. Dis. 2011, 204 (Suppl. 3), S941–S946. [Google Scholar] [CrossRef]
- Tsuda, Y.; Hoenen, T.; Banadyga, L.; Weisend, C.; Ricklefs, S.M.; Porcella, S.F.; Ebihara, H. An Improved Reverse Genetics System to Overcome Cell-Type-Dependent Ebola Virus Genome Plasticity. J. Infect. Dis. 2015, 212 (Suppl. 2), S129–S137. [Google Scholar] [CrossRef]
- Hoenen, T.; Marzi, A.; Scott, D.P.; Feldmann, F.; Callison, J.; Safronetz, D.; Ebihara, H.; Feldmann, H. Soluble Glycoprotein Is Not Required for Ebola Virus Virulence in Guinea Pigs. J. Infect. Dis. 2015, 212 (Suppl. 2), S242–S246. [Google Scholar] [CrossRef] [Green Version]
- Whitmer, S.L.M.; Ladner, J.T.; Wiley, M.R.; Patel, K.; Dudas, G.; Rambaut, A.; Sahr, F.; Prieto, K.; Shepard, S.S.; Carmody, E.; et al. Ebola Virus Persistence Study, G., Active Ebola Virus Replication and Heterogeneous Evolutionary Rates in EVD Survivors. Cell Rep. 2018, 22, 1159–1168. [Google Scholar] [CrossRef]
- Kindzelskii, A.L.; Yang, Z.; Nabel, G.J.; Todd, R.F., 3rd; Petty, H.R. Ebola virus secretory glycoprotein (sGP) diminishes Fc gamma RIIIB-to-CR3 proximity on neutrophils. J. Immunol. 2000, 164, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.H.; Harrison, A.; Corey, A.; Gentry, N.; Gregg, R.K. Ebola virus secreted glycoprotein decreases the anti-viral immunity of macrophages in early inflammatory responses. Cell Immunol. 2018, 324, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Mohan, G.S.; Li, W.; Ye, L.; Compans, R.W.; Yang, C. Antigenic subversion: A novel mechanism of host immune evasion by Ebola virus. PLoS Pathog. 2012, 8, e1003065. [Google Scholar] [CrossRef] [PubMed]
- Shahhosseini, S.; Das, D.; Qiu, X.; Feldmann, H.; Jones, S.M.; Suresh, M.R. Production and characterization of monoclonal antibodies against different epitopes of Ebola virus antigens. J. Virol. Methods 2007, 143, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Dowling, W.; Thompson, E.; Badger, C.; Mellquist, J.L.; Garrison, A.R.; Smith, J.M.; Paragas, J.; Hogan, R.J.; Schmaljohn, C. Influences of glycosylation on antigenicity, immunogenicity, and protective efficacy of ebola virus GP DNA vaccines. J. Virol. 2007, 81, 1821–1837. [Google Scholar] [CrossRef] [PubMed]
- Ilinykh, P.A.; Shen, X.; Flyak, A.I.; Kuzmina, N.; Ksiazek, T.G.; Crowe, J.E., Jr.; Bukreyev, A. Chimeric Filoviruses for Identification and Characterization of Monoclonal Antibodies. J. Virol. 2016, 90, 3890–3901. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ye, L.; Lin, F.; Gomaa, Y.; Flyer, D.; Carrion, R., Jr.; Patterson, J.L.; Prausnitz, M.R.; Smith, G.; Glenn, G.; et al. Intradermal Vaccination With Adjuvanted Ebola Virus Soluble Glycoprotein Subunit Vaccine by Microneedle Patches Protects Mice Against Lethal Ebola Virus Challenge. J. Infect. Dis. 2018, 218 (Suppl. 5), S545–S552. [Google Scholar] [CrossRef]
- Saphire, E.O.; Schendel, S.L.; Fusco, M.L.; Gangavarapu, K.; Gunn, B.M.; Wec, A.Z.; Halfmann, P.J.; Brannan, J.M.; Herbert, A.S.; Qiu, X.; et al. Viral Hemorrhagic Fever Immunotherapeutic, C., Systematic Analysis of Monoclonal Antibodies against Ebola Virus GP Defines Features that Contribute to Protection. Cell 2018, 174, 938–952.e13. [Google Scholar] [CrossRef]
- Qiu, X.; Wong, G.; Audet, J.; Bello, A.; Fernando, L.; Alimonti, J.B.; Fausther-Bovendo, H.; Wei, H.; Aviles, J.; Hiatt, E.; et al. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 2014, 514, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Gunn, B.M.; Roy, V.; Karim, M.M.; Hartnett, J.N.; Suscovich, T.J.; Goba, A.; Momoh, M.; Sandi, J.D.; Kanneh, L.; Andersen, K.G.; et al. Survivors of Ebola virus disease develop polyfunctional antibody responses. J. Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Sakabe, S.; Sullivan, B.M.; Hartnett, J.N.; Robles-Sikisaka, R.; Gangavarapu, K.; Cubitt, B.; Ware, B.C.; Kotliar, D.; Branco, L.M.; Goba, A.; et al. Analysis of CD8(+) T cell response during the 2013-2016 Ebola epidemic in West Africa. Proc. Natl. Acad. Sci. USA 2018, 115, E7578–E7586. [Google Scholar] [CrossRef] [PubMed]
- Radoshitzky, S.R.; Warfield, K.L.; Chi, X.; Dong, L.; Kota, K.; Bradfute, S.B.; Gearhart, J.D.; Retterer, C.; Kranzusch, P.J.; Misasi, J.N.; et al. Ebolavirus delta-peptide immunoadhesins inhibit marburgvirus and ebolavirus cell entry. J. Virol. 2011, 85, 8502–8513. [Google Scholar] [CrossRef] [PubMed]
- Gallaher, W.R.; Garry, R.F. Modeling of the Ebola virus delta peptide reveals a potential lytic sequence motif. Viruses 2015, 7, 285–305. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Melnik, L.I.; Komin, A.; Wiedman, G.; Fuselier, T.; Morris, C.F.; Starr, C.G.; Searson, P.C.; Gallaher, W.R.; Hristova, K.; et al. Ebola Virus Delta Peptide Is a Viroporin. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, H.; Volchkov, V.E.; Volchkova, V.A.; Stroher, U.; Klenk, H.D. Biosynthesis and role of filoviral glycoproteins. J. Gen. Virol. 2001, 82 Pt 12, 2839–2848. [Google Scholar] [CrossRef] [Green Version]
- Hoenen, T.; Groseth, A.; Feldmann, H. Therapeutic strategies to target the Ebola virus life cycle. Nat. Rev. Microbiol. 2019, 17, 593–606. [Google Scholar] [CrossRef]
- Lucey, D.R. New treatments for Ebola virus disease. BMJ 2019, 366, l5371. [Google Scholar] [CrossRef]
- Shears, P.; O’Dempsey, T.J. Ebola virus disease in Africa: Epidemiology and nosocomial transmission. J. Hosp. Infect. 2015, 90, 1–9. [Google Scholar] [CrossRef]
Figure 1.
Glycoprotein (GP) gene editing in Ebola virus (EBOV). Highlighted (in red) in the EBOV GP gene are seven consecutive uridine (U) residues that act as the transcriptional editing site. (A) The EBOV genome encodes seven structural proteins, including the nucleoprotein (NP), virion protein (VP) 35, VP40, glycoprotein (GP1,2), VP30, VP24, and the RNA-dependent RNA polymerase (L). (B) The majority (~70%) of transcripts produced are unedited, containing seven adenosine (7A) residues, and translated to produce soluble glycoprotein (sGP), which yields delta (Δ)-peptide upon proteolytic cleavage. (C) Approximately 25% of the transcripts contain eight adenosine (8A) residues due to the addition of a non-templated A (in blue) added by the RNA polymerase as it stutters at the editing site. The non-templated A results in a frameshift that extends the length of the open reading frame, giving rise to the surface glycoprotein (GP1,2). (D) For approximately 5% of the total transcripts, RNA polymerase stuttering at the editing site results in mRNA transcripts containing either six adenosine (6A) or nine adenosine (9A) residues, both of which encoding the small soluble glycoprotein (ssGP).
Figure 1.
Glycoprotein (GP) gene editing in Ebola virus (EBOV). Highlighted (in red) in the EBOV GP gene are seven consecutive uridine (U) residues that act as the transcriptional editing site. (A) The EBOV genome encodes seven structural proteins, including the nucleoprotein (NP), virion protein (VP) 35, VP40, glycoprotein (GP1,2), VP30, VP24, and the RNA-dependent RNA polymerase (L). (B) The majority (~70%) of transcripts produced are unedited, containing seven adenosine (7A) residues, and translated to produce soluble glycoprotein (sGP), which yields delta (Δ)-peptide upon proteolytic cleavage. (C) Approximately 25% of the transcripts contain eight adenosine (8A) residues due to the addition of a non-templated A (in blue) added by the RNA polymerase as it stutters at the editing site. The non-templated A results in a frameshift that extends the length of the open reading frame, giving rise to the surface glycoprotein (GP1,2). (D) For approximately 5% of the total transcripts, RNA polymerase stuttering at the editing site results in mRNA transcripts containing either six adenosine (6A) or nine adenosine (9A) residues, both of which encoding the small soluble glycoprotein (ssGP).

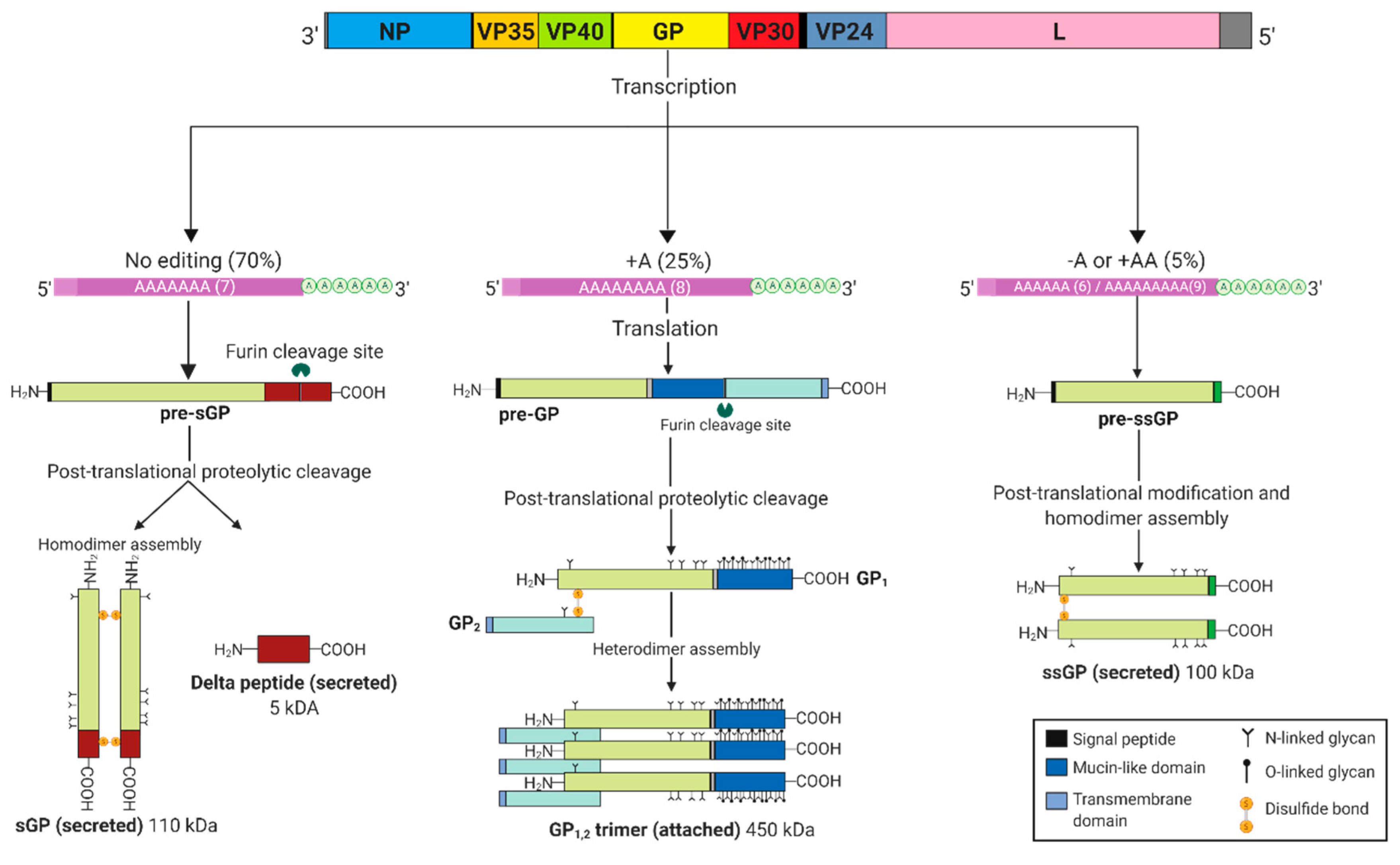
Figure 2.
Ebolavirus GP gene products. Co-transcriptional editing at the GP gene editing site results in three major transcripts that are afterwards translated to various glycoprotein products, initially synthesized as pre-sGP, pre-GP and pre-ssGP. Post-translational cleavage by furin at the carboxy terminus of pre-sGP generates a 5 kDa delta (Δ)-peptide and an sGP monomer that further assembles into a 110 kDa homodimer linked by two disulfide bonds. Similarly, pre-GP (also known as GP0) undergoes a proteolytic cleavage resulting in the formation of a disulfide linked GP1,2 heterodimer that trimerizes into a 450 kDa viral surface glycoprotein. Pre-ssGP does not undergo post-translational cleavage, but likewise homodimerizes via formation of a single disulfide bond. Additionally, all the glycoproteins are N-glycosylated, with GP1,2 experiencing further O-glycosylation.
Figure 2.
Ebolavirus GP gene products. Co-transcriptional editing at the GP gene editing site results in three major transcripts that are afterwards translated to various glycoprotein products, initially synthesized as pre-sGP, pre-GP and pre-ssGP. Post-translational cleavage by furin at the carboxy terminus of pre-sGP generates a 5 kDa delta (Δ)-peptide and an sGP monomer that further assembles into a 110 kDa homodimer linked by two disulfide bonds. Similarly, pre-GP (also known as GP0) undergoes a proteolytic cleavage resulting in the formation of a disulfide linked GP1,2 heterodimer that trimerizes into a 450 kDa viral surface glycoprotein. Pre-ssGP does not undergo post-translational cleavage, but likewise homodimerizes via formation of a single disulfide bond. Additionally, all the glycoproteins are N-glycosylated, with GP1,2 experiencing further O-glycosylation.

Figure 3.
Possible functions of sGP in the pathogenesis of EBOV. Dimeric sGP is secreted into the extracellular space where it may facilitate antigenic subversion (A), act as a decoy antigen (B), prevent activation of neutrophils (C), reduce the production of pro-inflammatory cytokines (D), and/or inhibit immune cell chemotaxis (E). Consequently, a combination of inactivated neutrophils and reduced levels of pro-inflammatory cytokines may aid in restoring endothelial barrier function (F).
Figure 3.
Possible functions of sGP in the pathogenesis of EBOV. Dimeric sGP is secreted into the extracellular space where it may facilitate antigenic subversion (A), act as a decoy antigen (B), prevent activation of neutrophils (C), reduce the production of pro-inflammatory cytokines (D), and/or inhibit immune cell chemotaxis (E). Consequently, a combination of inactivated neutrophils and reduced levels of pro-inflammatory cytokines may aid in restoring endothelial barrier function (F).

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhu, W.; Banadyga, L.; Emeterio, K.; Wong, G.; Qiu, X. The Roles of Ebola Virus Soluble Glycoprotein in Replication, Pathogenesis, and Countermeasure Development. Viruses 2019, 11, 999. https://0-doi-org.brum.beds.ac.uk/10.3390/v11110999
AMA Style
Zhu W, Banadyga L, Emeterio K, Wong G, Qiu X. The Roles of Ebola Virus Soluble Glycoprotein in Replication, Pathogenesis, and Countermeasure Development. Viruses. 2019; 11(11):999. https://0-doi-org.brum.beds.ac.uk/10.3390/v11110999
Chicago/Turabian StyleZhu, Wenjun, Logan Banadyga, Karla Emeterio, Gary Wong, and Xiangguo Qiu. 2019. "The Roles of Ebola Virus Soluble Glycoprotein in Replication, Pathogenesis, and Countermeasure Development" Viruses 11, no. 11: 999. https://0-doi-org.brum.beds.ac.uk/10.3390/v11110999
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.