Metagenomic Analysis of the Enteric RNA Virome of Infants from the Oukasie Clinic, North West Province, South Africa, Reveals Diverse Eukaryotic Viruses


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Enrichment of RNA Virome
2.3. Next Generation Sequencing
2.4. Data Analysis
3. Results
3.1. Abundance of Viral Contigs
3.2. Taxonomic Classification of Viral Contigs
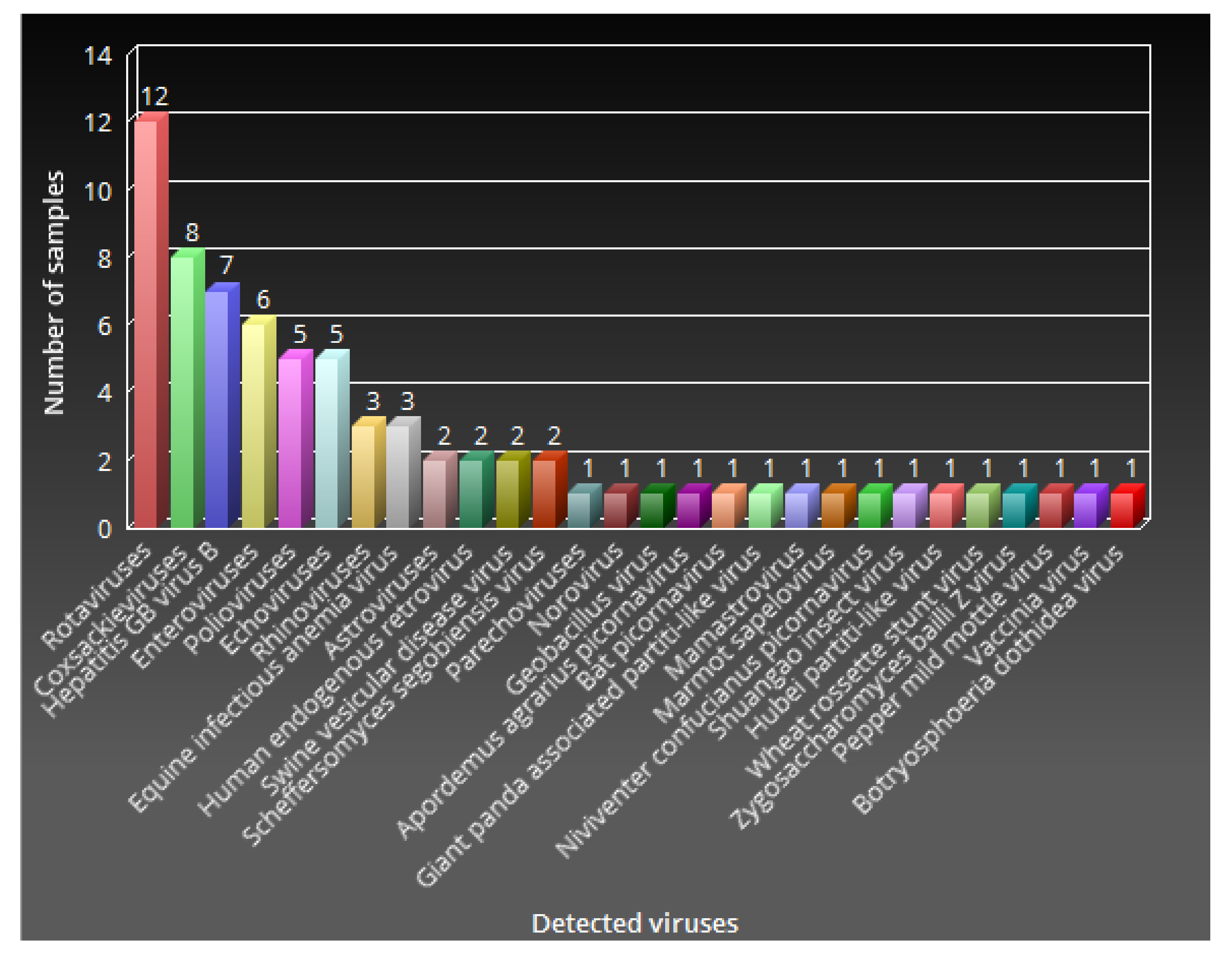
3.3. Frequency of Virus Detection and Abundance by Host Specificity
3.4. Distribution and Abundance of Mammalian Enteric RNA Viruses
3.5. Gut Virome Composition and Changes over Time for Each Study Participant
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the Human Infant Intestinal Microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef] [Green Version]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, S.T.N.S.J.C. The microbiome in early life: Implications for health outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Palacio, S.D.; Montes, S.A.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, A.; Ryan, F.J.; Stockdale, S.R.; Dalmasso, M.; Blake, T.; Ryan, C.A.; Stanton, C.; Mills, S.; Ross, P.R.; Hill, C. Viromes of one year old infants reveal the impact of birth mode on microbiome diversity. PeerJ 2018, 6, e4694. [Google Scholar] [CrossRef] [Green Version]
- Haynes, M.; Rohwer, F. The Human Virome. In Metagenomics of the Human Body; Springer: New York, NY, USA, 2010; pp. 63–77. [Google Scholar] [CrossRef]
- Minot, S.; Bryson, A.; Chehoud, C.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. Rapid evolution of the human gut virome. Proc. Natl. Acad. Sci. USA 2013, 110, 12450–12455. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, S.R.; Wang, D. Origins and challenges of viral dark matter. Virus Res. 2017, 239, 136–142. [Google Scholar] [CrossRef]
- Virgin, H.W. The Virome in Mammalian Physiology and Disease. Cell 2014, 157, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Clark, B.; McKendrick, M. A review of viral gastroenteritis. Curr. Opin. Infect. Dis. 2004, 17, 461–469. [Google Scholar] [CrossRef]
- Glass, R.I.; Parashar, U.D.; Estes, M.K. Norovirus Gastroenteritis. N. Engl. J. Med. 2009, 361, 1776–1785. [Google Scholar] [CrossRef] [Green Version]
- Eckardt, A.J. Viral Gastroenteritis in Adults. Recent Pat. Anti Infect. Drug Discov. 2011, 6, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Kapusinszky, B.; Minor, P.; Delwart, E. Nearly Constant Shedding of Diverse Enteric Viruses by Two Healthy Infants. J. Clin. Microbiol. 2012, 50, 3427–3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Focà, A.; Liberto, M.C.; Quirino, A.; Marascio, N.; Zicca, E.; Pavia, G. Gut Inflammation and Immunity: What Is the Role of the Human Gut Virome? Mediat. Inflamm. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of under-5 mortality in 2000–15: An updated systematic analysis with implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef] [Green Version]
- Black, R.E.; Cousens, S.; Johnson, H.L.; Lawn, J.E.; Rudan, I.; Bassani, D.G.; Jha, P.; Campbell, H.; Walker, C.F.; Cibulskis, R.; et al. Global, regional, and national causes of child mortality in 2008: A systematic analysis. Lancet 2010, 375, 1969–1987. [Google Scholar] [CrossRef]
- Breitbart, M.; Haynes, M.; Kelley, S.; Angly, F.; Edwards, R.A.; Felts, B.; Mahaffy, J.M.; Mueller, J.; Nulton, J.; Rayhawk, S.; et al. Viral diversity and dynamics in an infant gut. Res. Microbiol. 2008, 159, 367–373. [Google Scholar] [CrossRef]
- Reyes, A.; Haynes, M.; Hanson, N.; Angly, F.E.; Heath, A.C.; Rohwer, F.; Gordon, J.I. Viruses in the faecal microbiota of monozygotic twins and their mothers. Nat. Cell Biol. 2010, 466, 334–338. [Google Scholar] [CrossRef]
- Zhang, T.; Breitbart, M.; Lee, W.H.; Run, J.Q.; Wei, C.L.; Soh, S.W.L.; Hibberd, M.L.; Liu, E.T.; Rohwer, F.; Ruan, Y. RNA Viral Community in Human Feces: Prevalence of Plant Pathogenic Viruses. PLoS Biol. 2005, 4, e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conceição-Neto, N.; Zeller, M.; Lefrère, H.; De Bruyn, P.; Beller, L.; Deboutte, W.; Yinda, C.K.; Lavigne, R.; Maes, P.; Van Ranst, M.; et al. Modular approach to customise sample preparation procedures for viral metagenomics: A reproducible protocol for virome analysis. Sci. Rep. 2015, 5, 16532. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FASTQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 23 April 2017).
- Schmieder, R.; Edwards, R.A. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Kew, O.M.; Sutter, R.W.; Nottay, B.K.; McDonough, M.J.; Prevots, D.R.; Quick, L.; Pallansch, M.A. Prolonged Replication of a Type 1 Vaccine-Derived Poliovirus in an Immunodeficient Patient. J. Clin. Microbiol. 1998, 36, 2893–2899. [Google Scholar] [CrossRef] [Green Version]
- Hovi, T.; Lindholm, N.; Savolainen, C.; Stenvik, M. Burns Evolution of wild-type 1 poliovirus in two healthy siblings excreting the virus over a period of 6 months. J. Gen. Virol. 2004, 85, 369–377. [Google Scholar] [CrossRef]
- Bellmunt, A.; May, G.; Zell, R.; Pring-Åkerblom, P.; Verhagen, W.; Heim, A. Evolution of Poliovirus Type I during 5.5 Years of Prolonged Enteral Replication in an Immunodeficient Patient. Virology 1999, 265, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Shulman, L.M.; Manor, J.; Handsher, R.; Delpeyroux, F.; MacDonough, M.J.; Halmut, T.; Silberstein, I.; Alfandari, J.; Quay, J.; Fisher, T.; et al. Mendelson Molecular and antigenic characterisation of a highly evolved derivative of the type 2 oral polio vaccine strain isolated from sewage in Israel. J. Clin. Microbiol. 2000, 38, 3729–3734. [Google Scholar] [CrossRef] [Green Version]
- Rotbart, H.A. Viral Meningitis. Semin. Neurol. 2000, 20, 277–292. [Google Scholar] [CrossRef] [PubMed]
- Jubelt, B.; Lipton, H.L. Enterovirus/Picornavirus Infections. In Handbook of Clinical Neurology; Elsevier BV: Amsterdam, The Netherlands, 2014; Volume 123, pp. 379–416. [Google Scholar]
- Pallansch, M.A.; Oberste, M.S.; Whitton, J.L. Enteroviruses: Polioviruses, Coxsackieviruses, Echoviruses, and Newer Enteroviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2013; pp. 491–530. [Google Scholar]
- Pons-Salort, M.; Parker, E.P.K.; Grassly, N.C. The epidemiology of non-polio enteroviruses. Curr. Opin. Infect. Dis. 2015, 28, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Smuts, H.E.M.; Cronje, S.; Thomas, J.; Brink, D.; Korsman, S.; Hardie, D.R. Molecular characterization of an outbreak of enterovirus-associated meningitis in Mossel Bay, South Africa, December 2015–January 2016. BMC Infect. Dis. 2018, 18, 709. [Google Scholar] [CrossRef]
- Wolfaardt, M.; Buchner, A.; Myburgh, M.; Avenant, T.; du Plessis, N.M.; Taylor, M. Molecular characterization of enteroviruses and clinical findings from a cluster of paediatric viral meningitis cases in Tshwane, South Africa 2010–2011. J. Clin. Viral. 2014, 61, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, G.; Bines, J.E.; Brewer, T.G.; Colombara, D.V.; et al. Rotavirus Vaccination and the Global Burden of Rotavirus Diarrhea Among Children Younger Than 5 Years. JAMA Pediatr. 2018, 172, 958–965. [Google Scholar] [CrossRef] [Green Version]
- Seheri, L.M.; Page, N.A.; Mawela, M.P.; Mphahlele, M.J.; Steele, A.D. Rotavirus vaccination within the South African Expanded Programme on Immunisation. Vaccine 2012, 30, C14–C20. [Google Scholar] [CrossRef]
- Sakon, N.; Miyamoto, R.; Komano, J. An infant with acute gastroenteritis caused by a secondary infection with a Rotarix-derived strain. Eur. J. Nucl. Med. Mol. Imaging 2017, 176, 1275–1278. [Google Scholar] [CrossRef]
- Van Beek, J.; Ambert-Balay, K.; Botteldoorn, N.; Eden, J.S.; Fonager, J.; Hewitt, J.; Iritani, N.; Kroneman, A.; Vennema, H.; Vinjé, J.; et al. Indications for worldwide increased norovirus activity associated with emergence of a new variant of genotype II.4, late 2012. Eurosurveillance 2013, 18, 8–9. [Google Scholar]
- Ayukekbong, J.A.; Mesumbe, H.N.; Oyero, O.G.; Lindh, M.; Bergström, T. Role of noroviruses as aetiological agents of diarrhoea in developing countries. J. Gen. Virol. 2015, 96, 1983–1999. [Google Scholar] [CrossRef] [PubMed]
- Minot, S.; Sinha, R.; Chen, J.; Li, H.; Keilbaugh, S.A.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. The human gut virome: Inter-individual variation and dynamic response to diet. Genome Res. 2011, 21, 1616–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotton, M.; Oude Munnink, B.; Canuti, M.; Deijs, M.; Watson, S.J.; Kellam, P.; van der Hoek, L. Full genome virus detection in faecal samples using sensitive nucleic acid preparation, deep sequencing, and a novel iterative sequence classification algorithm. PLoS ONE 2014, 9, e93269. [Google Scholar]
- Lim, E.S.; Zhou, Y.; Zhao, G.; Bauer, I.K.; Droit, L.; Ndao, I.M.; Warner, B.B.; Tarr, P.I.; Wang, D.; Holtz, L.R. Early life dynamics of the human gut virome and bacterial microbiome in infants. Nat. Med. 2015, 21, 1228–1234. [Google Scholar] [CrossRef]
- Salminen, K.K.; Vuorinen, T.; Oikarinen, S.; Helminen, M.; Simell, S.; Knip, M.; Ilonen, J.; Simell, O.; Hyöty, H. Isolation of enterovirus strains from children with preclinical Type 1 diabetes. Diabet. Med. 2004, 21, 156–164. [Google Scholar] [CrossRef]
- Harvala, H.; McIntyre, C.L.; McLeish, N.J.; Kondracka, J.; Palmer, J.; Molyneaux, P.; Gunson, R.; Bennett, S.; Templeton, K.; Simmonds, P. High detection frequency and viral loads of human rhinovirus species A to C in faecal samples; diagnostic and clinical implications. J. Med. Virol. 2012, 84, 536–542. [Google Scholar] [CrossRef]
- Honkanen, H.; Oikarinen, S.; Peltonen, P.; Simell, O.; Ilonen, J.; Veijola, R.; Knip, M.; Hyöty, H. Human rhinoviruses including group C are common in stool samples of young Finnish children. J. Clin. Virol. 2013, 56, 334–338. [Google Scholar] [CrossRef]
- Issel, C.; Foil, L. Equine infectious anaemia and mechanical transmission: Man and the wee beasties. Rev. Sci. Tech. OIE 2015, 34, 513–523. [Google Scholar] [CrossRef]
- Lin, F.; Kitching, R. Swine Vesicular Disease: An Overview. Vet. J. 2000, 160, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Graul, S.; Böttcher, S.; Eibach, D.; Krumkamp, R.; Käsmaier, J.; Adu-Sarkodie, Y.; May, J.; Tannich, E.; Panning, M. High diversity of human parechovirus including novel types in stool samples from Ghanaian children. J. Clin. Virol. 2017, 96, 116–119. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Collection Time | Total Assembled Contigs | Viral Contigs | Percentage Viral Contigs |
|---|---|---|---|---|
| A1 | 6 weeks | 5549 | 68 | 1.2 |
| A2 | 10 weeks | 6255 | 28 | 0.4 |
| A3 | 36 weeks | 9805 | 489 | 5.0 |
| Total | 21,609 | 585 | 2.7% | |
| B1 | 10 weeks | 12,445 | 2504 | 20.1 |
| B2 | 14 weeks | 15,184 | 7064 | 46.5 |
| B3 | 24 weeks | 5636 | 138 | 2.4 |
| Total | 33,265 | 9706 | 29.2% | |
| C1 | 6 weeks | 4368 | 87 | 2.0 |
| C2 | 14 weeks | 8637 | 60 | 0.7 |
| C3 | 20 weeks | 3638 | 67 | 1.8 |
| Total | 16,643 | 214 | 1.3% | |
| D1 | 6 weeks | 7405 | 36 | 0.5 |
| D2 | 14 weeks | 7357 | 37 | 0.5 |
| D3 | 20 weeks | 5906 | 70 | 1.2 |
| Total | 20,668 | 143 | 0.7% | |
| Overall | 92,185 | 10,648 | 11.6% |
| Virus Family | Picornaviridae | Reoviridae | Astroviridae | Retroviridae | Caliciviridae | Flaviviridae | Poxviridae | Virgaviridae | Rhabdoviridae | Siphoviridae | Amalgaviridae | PartitiviridaE | Totiviridae | Peribunyaviridae | Unclassified picornavirales | Unclassified | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SAMPLE NUMBER | Sample ID | Sample Collection Time (Weeks) | Total Number of Contigs | Contigs Assigned to Viruses | Coxsackievirus A, B | Enterovirus A, B, C, D | Echovirus E | Poliovirus 1, 2, 3 | Parechovirus 1, 3, 8, 17, 19 | Rhinovirus A, B, C | Swine vesicular disease virus | Bat picornavirus | Marmot sapelovirus | Rotavirus A | Astrovirus | Mamastrovirus | Human endogenous retrovirus | Equine infectious anaemia virus | Norovirus GI, GII.4 | Hepatitis GB virus B | Vaccinia virus | Pepper mild mottle virus | Wheat rosette stunt virus | Geobacillus virus | Zygosaccharomyces bailli Z virus | Botryosphoeria dothidea virus | Scheffersomyces segobiensis virus | Shuangao insect virus | Apodemus agrarius picornavirus | Niniventer confucianus picornavirus | Hubei partiti-like virus | Giant panda associated partiti-like virus |
| Genomic Structure | (+)ssRNA | dsRNA | (+)ssRNA | RNA | (+)ssRNA | (+)ssRNA | (+)ssRNA | dsDNA | (+)ssRNA | (−)ssRNA | dsDNA | dsRNA | dsRNA | dsRNA | (−)ssRNA | (+)ssRNA | (+)ssRNA | RNA | RNA | |||||||||||||
| A1 | 4450 | 6 | 5549 | 68 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 58 | 0 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 |
| A2 | 6618 | 10 | 6255 | 28 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 27 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A3 | 8908 | 36 | 9805 | 489 | 84 | 34 | 309 | 0 | 0 | 0 | 2 | 0 | 0 | 59 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B1 | 8903 | 10 | 12,445 | 2504 | 1386 | 146 | 5 | 914 | 0 | 0 | 0 | 0 | 0 | 51 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B2 | 8941 | 14 | 15,184 | 7064 | 3953 | 549 | 22 | 2333 | 0 | 88 | 3 | 1 | 1 | 90 | 9 | 1 | 0 | 1 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 1 | 0 | 1 |
| B3 | 10,287 | 24 | 5636 | 138 | 2 | 1 | 0 | 4 | 94 | 0 | 0 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 4 | 6 | 1 | 1 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 1 | 0 |
| C1 | 8824 | 6 | 4368 | 87 | 5 | 4 | 20 | 1 | 0 | 0 | 0 | 0 | 0 | 52 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| C2 | 10,127 | 14 | 8637 | 60 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 57 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| C3 | 10,233 | 20 | 3638 | 67 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 43 | 0 | 0 | 0 | 17 | 0 | 2 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| D1 | 8355 | 6 | 7405 | 36 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 32 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| D2 | 8910 | 14 | 7357 | 37 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 37 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| D3 | 10,098 | 20 | 5906 | 70 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 67 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| TOTAL VIRUS CONTIGS | 10,648 | 5435 | 735 | 357 | 3253 | 94 | 90 | 5 | 1 | 1 | 593 | 11 | 1 | 3 | 19 | 4 | 21 | 1 | 1 | 1 | 1 | 1 | 1 | 10 | 1 | 1 | 1 | 1 | 1 | |||
| Sample Number | Collection Time | (+)ssRNA Viruses | (−)ssRNA Viruses | dsRNA Viruses | DNA Viruses | Unclassified RNA Viruses | Total (Different Viral Genomes) |
|---|---|---|---|---|---|---|---|
| A1 | 6 weeks | 3 | - | 2 | - | - | 5 |
| A2 | 10 weeks | 1 | - | 1 | - | - | 2 |
| A3 | 36 weeks | 5 | - | 1 | - | - | 6 |
| B1 | 10 weeks | 5 | - | 1 | - | - | 6 |
| B2 | 14 weeks | 13 | 1 | 2 | - | 1 | 17 |
| B3 | 24 weeks | 8 | - | 2 | 1 | 1 | 12 |
| C1 | 6 weeks | 5 | - | 2 | - | - | 7 |
| C2 | 14 weeks | 2 | - | 1 | - | - | 3 |
| C3 | 20 weeks | 5 | 1 | 2 | - | - | 8 |
| D1 | 6 weeks | 1 | - | 1 | 1 | 1 | 4 |
| D2 | 14 weeks | - | - | 1 | - | - | 1 |
| D3 | 20 weeks | 1 | - | 1 | - | 1 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mogotsi, M.T.; Mwangi, P.N.; Bester, P.A.; Mphahlele, M.J.; Seheri, M.L.; O’Neill, H.G.; Nyaga, M.M. Metagenomic Analysis of the Enteric RNA Virome of Infants from the Oukasie Clinic, North West Province, South Africa, Reveals Diverse Eukaryotic Viruses. Viruses 2020, 12, 1260. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111260
Mogotsi MT, Mwangi PN, Bester PA, Mphahlele MJ, Seheri ML, O’Neill HG, Nyaga MM. Metagenomic Analysis of the Enteric RNA Virome of Infants from the Oukasie Clinic, North West Province, South Africa, Reveals Diverse Eukaryotic Viruses. Viruses. 2020; 12(11):1260. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111260
Chicago/Turabian StyleMogotsi, Milton T., Peter N. Mwangi, Phillip A. Bester, M. Jeffrey Mphahlele, Mapaseka L. Seheri, Hester G. O’Neill, and Martin M. Nyaga. 2020. "Metagenomic Analysis of the Enteric RNA Virome of Infants from the Oukasie Clinic, North West Province, South Africa, Reveals Diverse Eukaryotic Viruses" Viruses 12, no. 11: 1260. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111260