Structural and Biological Basis of Alphacoronavirus nsp1 Associated with Host Proliferation and Immune Evasion

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Viruses
2.2. Plasmid Construction
2.3. Protein Purification and Crystallization
2.4. Data Collection and Structure Determination
2.5. Reporter Assay and Western Blot Analysis
2.6. Indirect Immunofluorescence Assay (IFA)
2.7. Quantitative Analysis of Rluc mRNA
2.8. RNA Sequence Analysis
2.9. DNA Content Analysis
2.10. Growth Curves of Viruses
2.11. Protein Structure Accession Numbers and Statistical Analysis
3. Results
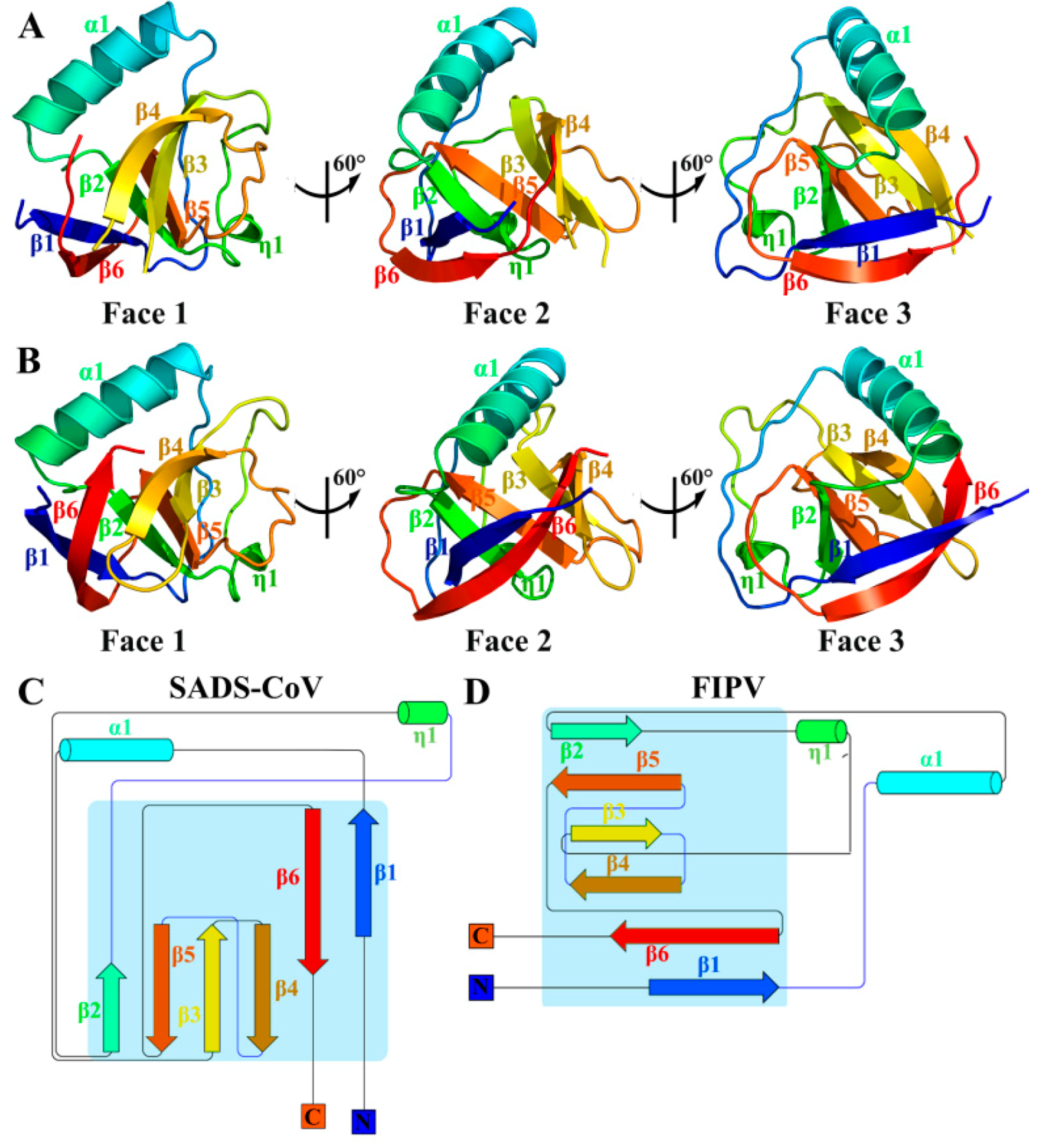
3.1. Overall Structures of SADS-CoV and FIPV nsp1
3.2. Sequence Analysis of Alphacoronaviruses nsp1
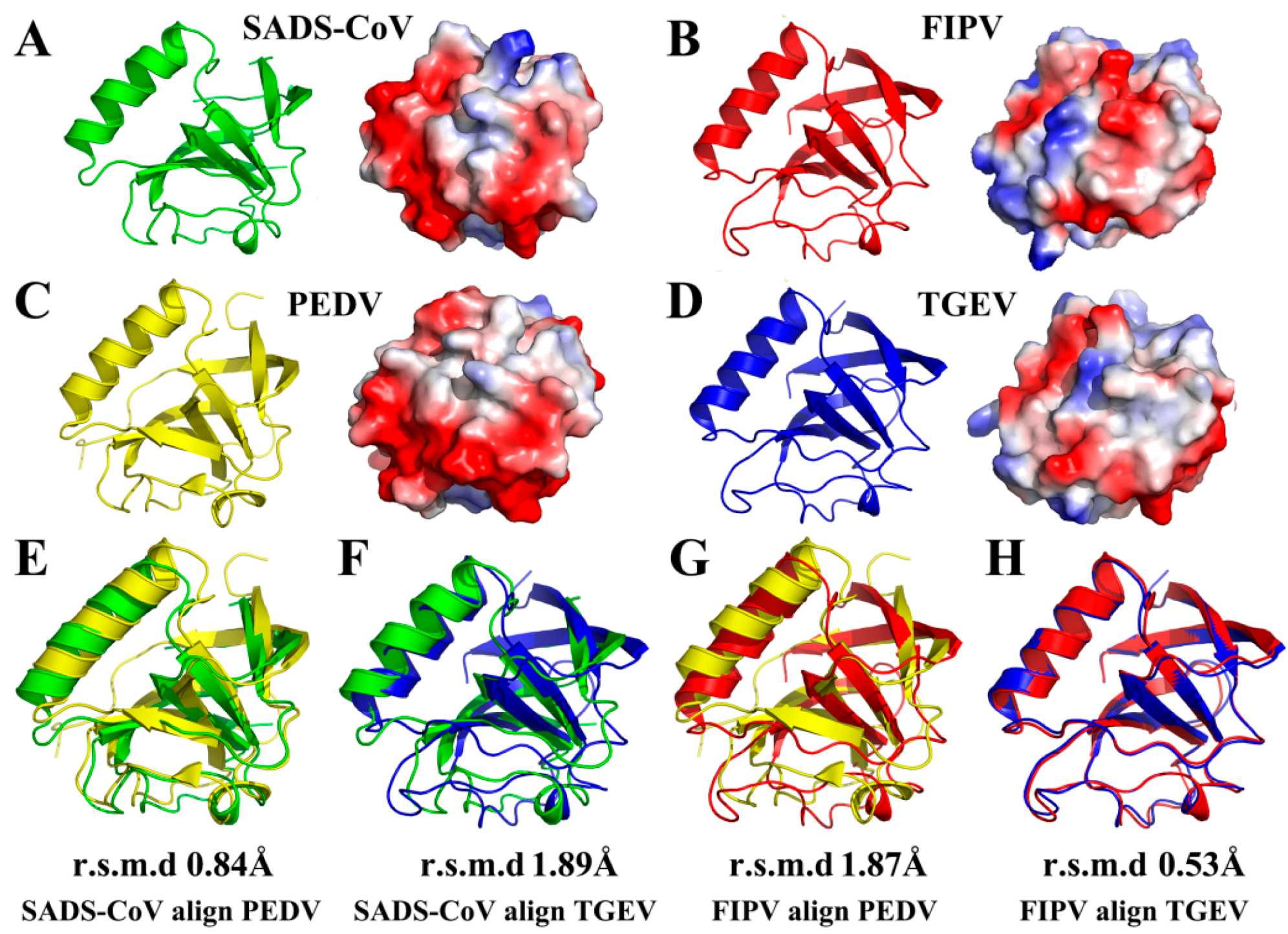
3.3. Structural Comparisons of Alphacoronavirus 1A and 1B nsp1 Proteins
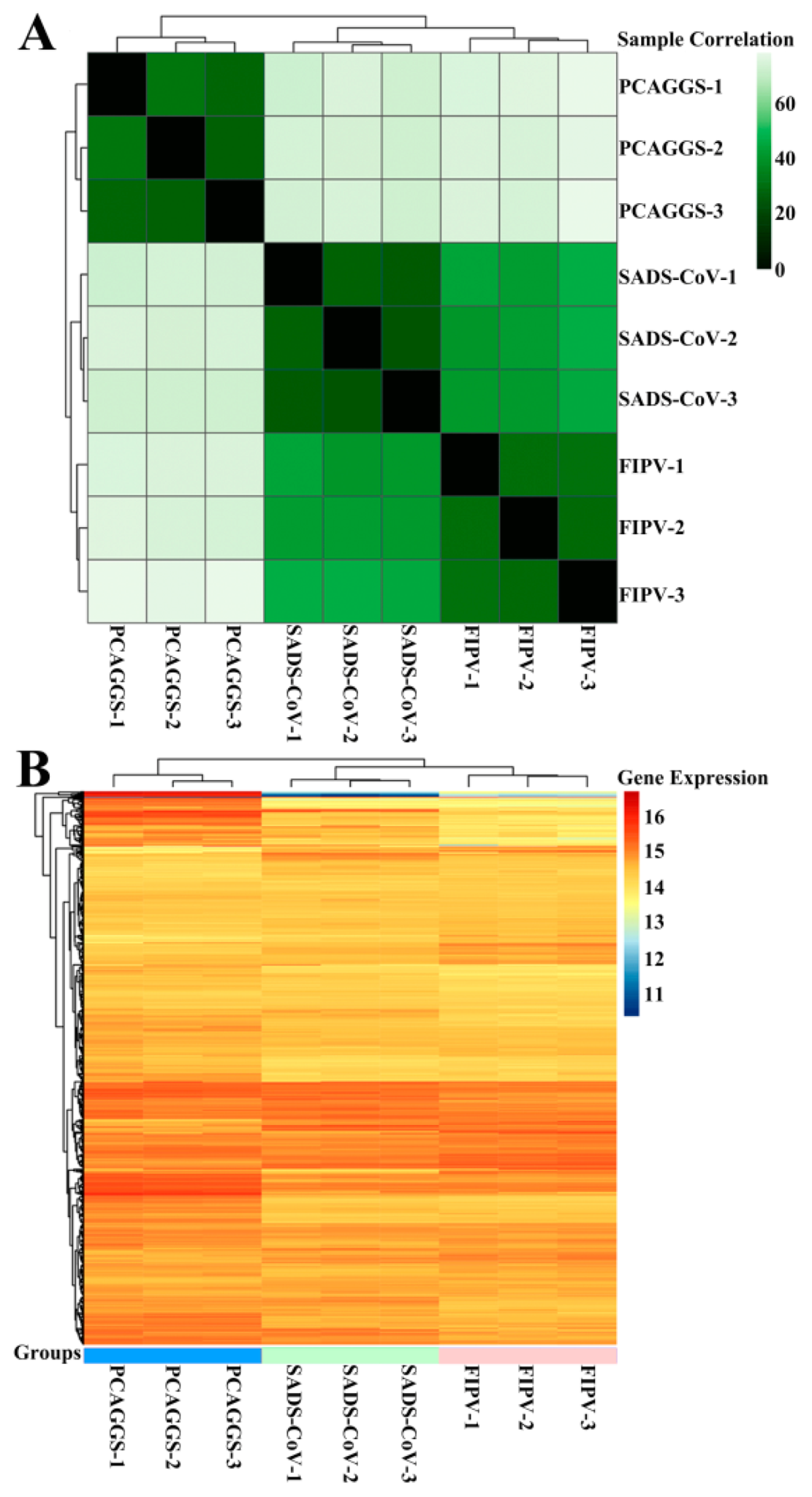
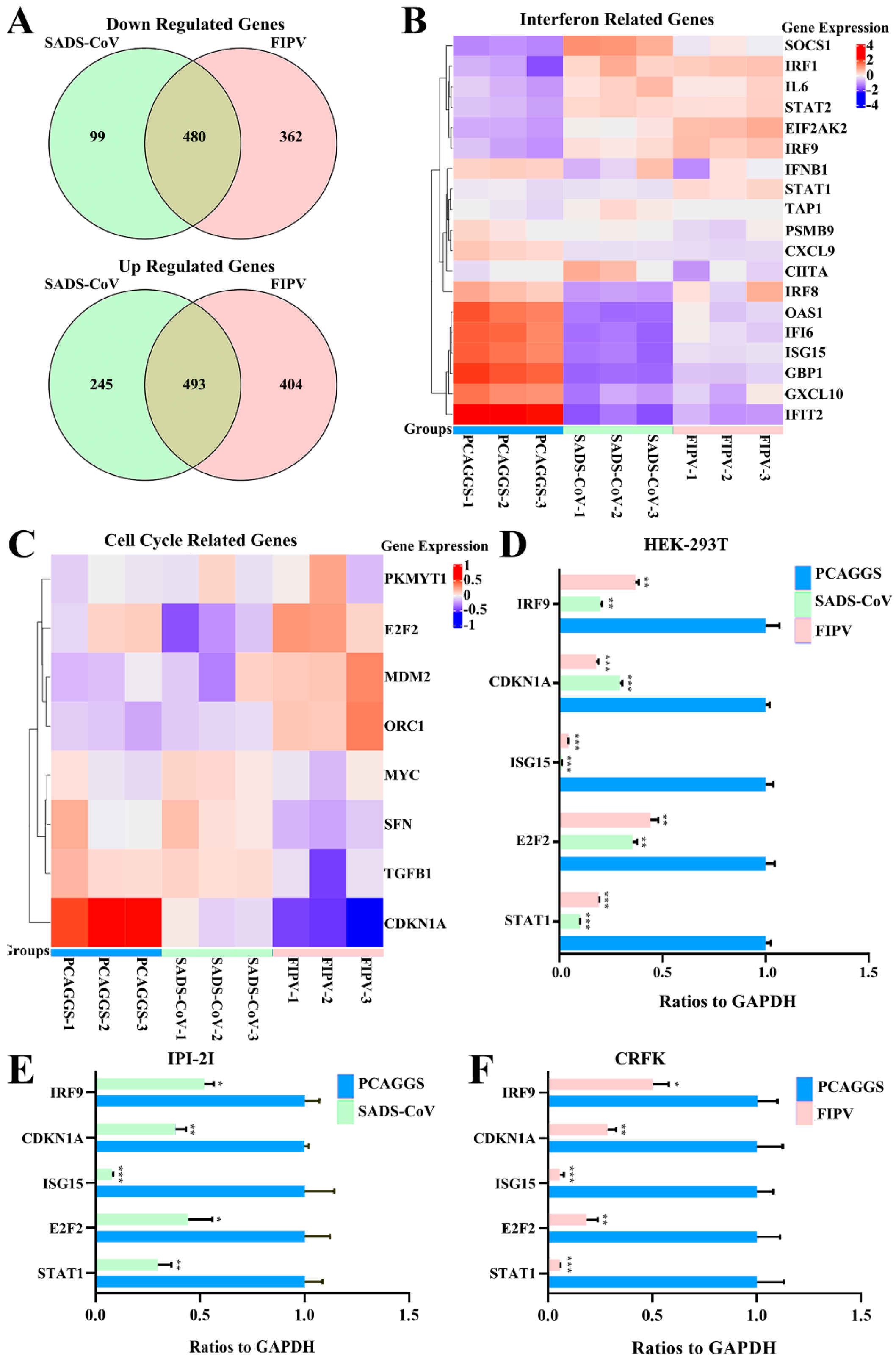
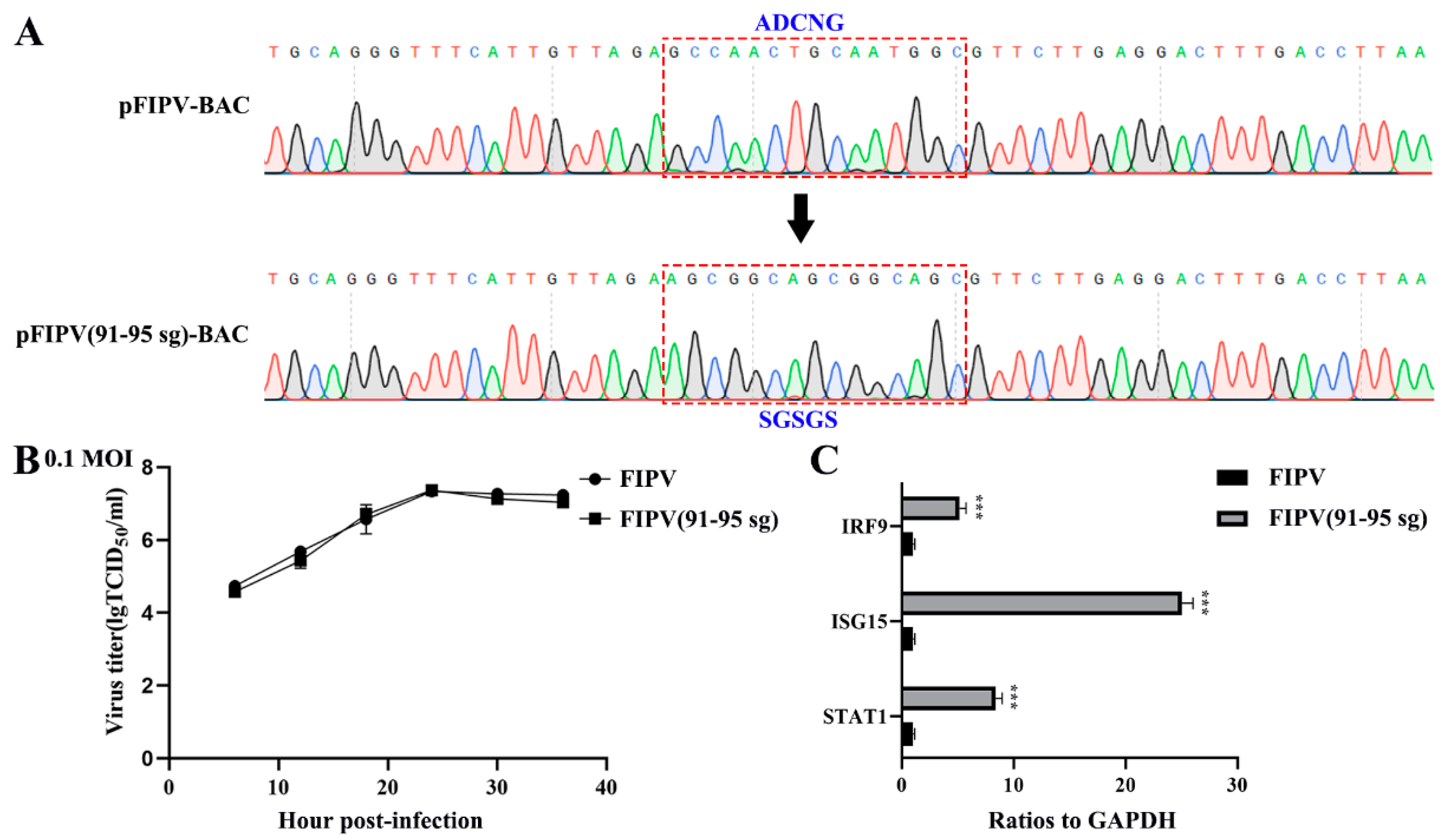
3.4. RNA Sequences in HEK-293T Cells Transfected with SADS-CoV or FIPV nsp1
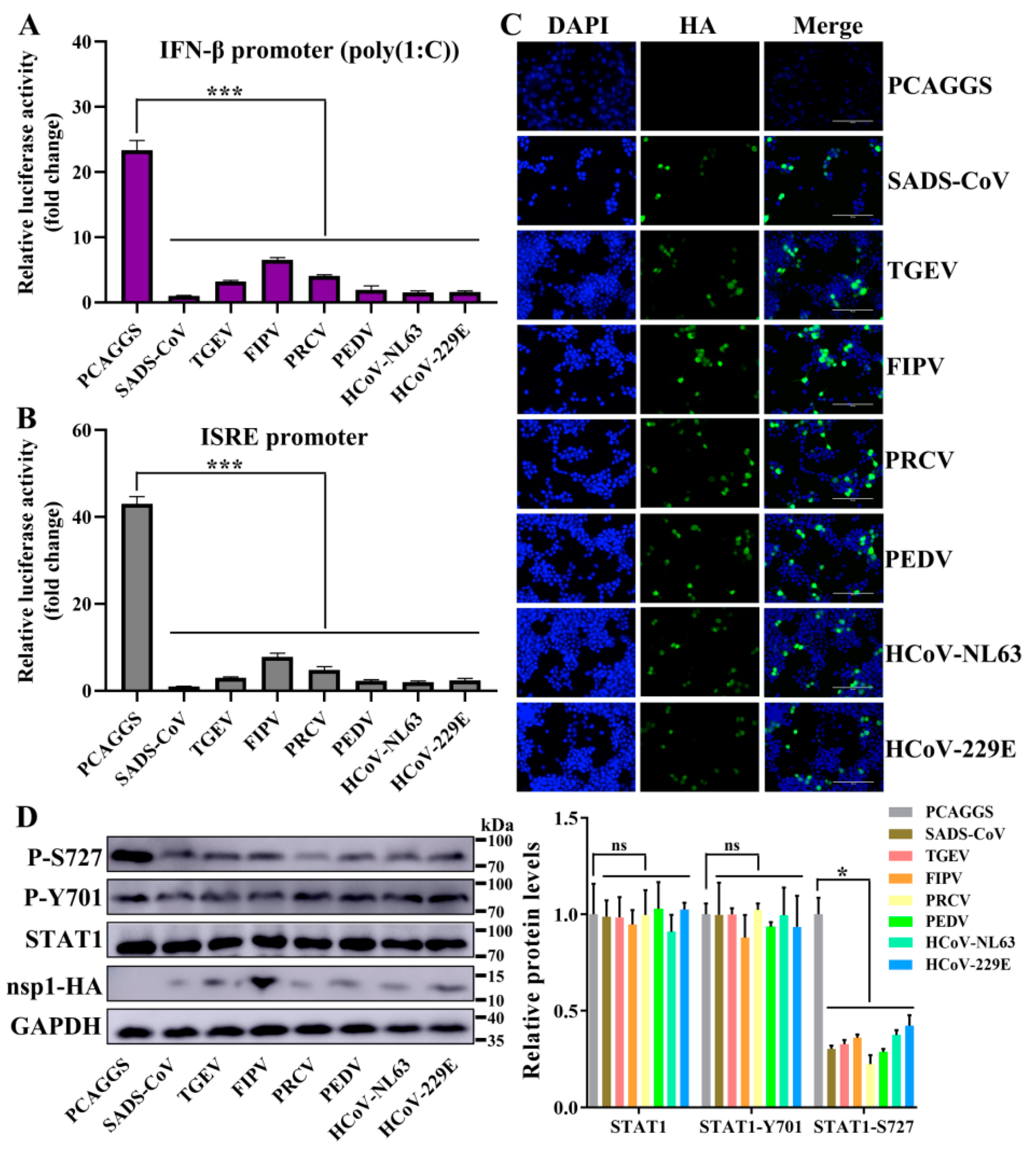
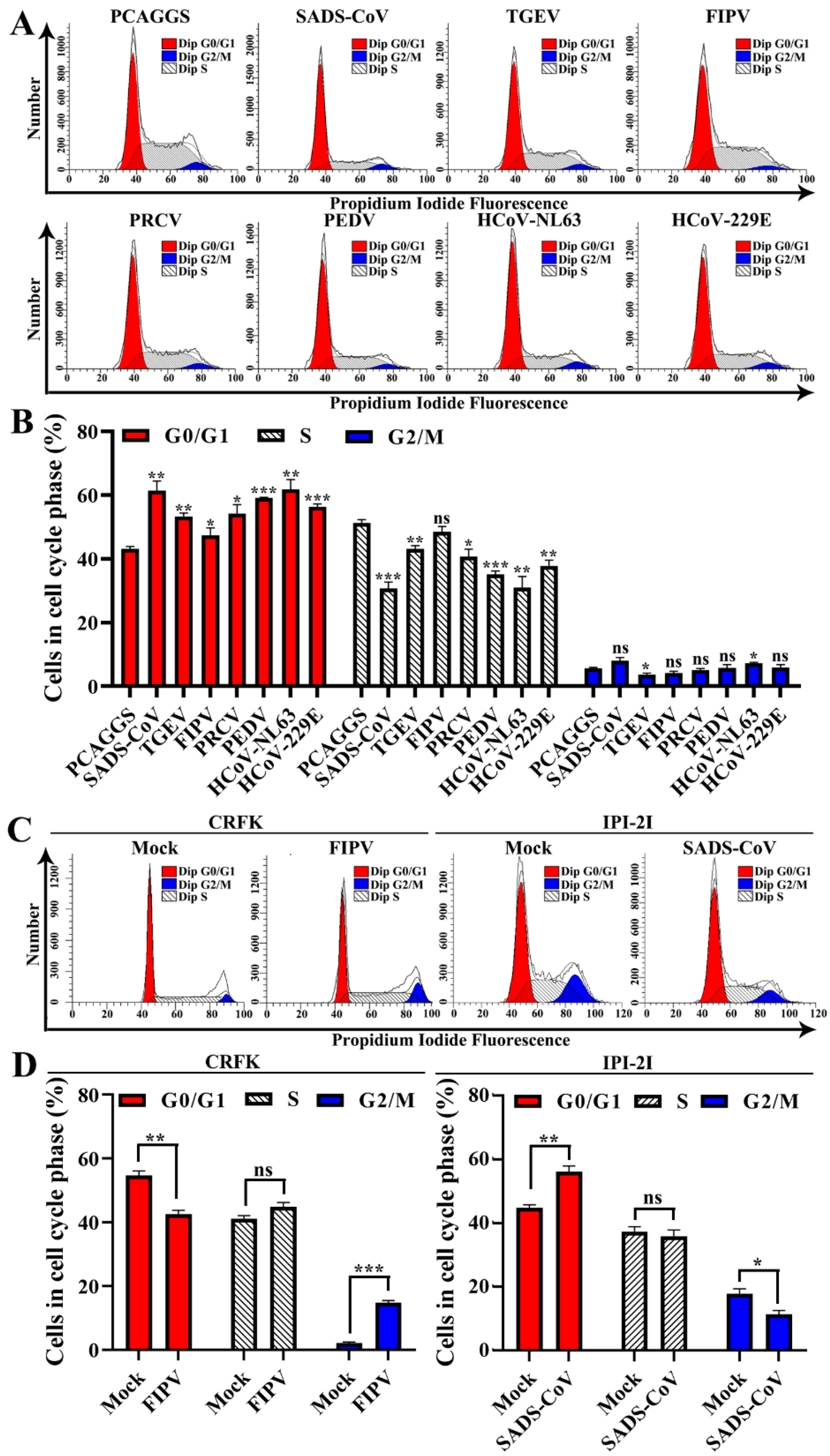
3.5. Alphacoronavirus nsp1 Can Regulate IFN-I and the Cell Cycle
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiang, S.; Du, L.; Shi, Z. An emerging coronavirus causing pneumonia outbreak in wuhan, china: Calling for developing therapeutic and prophylactic strategies. Emerg. Microbes Infect. 2020, 9, 275–277. [Google Scholar] [CrossRef]
- Weiss, S.R.; Leibowitz, J.L. Coronavirus pathogenesis. Adv. Virus Res. 2011, 81, 85–164. [Google Scholar]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [PubMed] [Green Version]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlman, S.; Netland, J. Coronaviruses post-sars: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.D.; Wunderink, R.G. Mers, sars and other coronaviruses as causes of pneumonia. Respirology 2018, 23, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, G.R.; Andre, N.M.; Millet, J.K. Improving virus taxonomy by recontextualizing sequence-based classification with biologically relevant data: The case of the alphacoronavirus 1 species. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an hku2-related coronavirus of bat origin. Nature 2018, 556, 255–258. [Google Scholar] [CrossRef]
- Li, K.; Li, H.; Bi, Z.; Song, D.; Zhang, F.; Lei, D.; Luo, S.; Li, Z.; Gong, W.; Huang, D.; et al. Significant inhibition of re-emerged and emerging swine enteric coronavirus in vitro using the multiple shrna expression vector. Antivir. Res. 2019, 166, 11–18. [Google Scholar] [CrossRef]
- Zhou, L.; Li, Q.N.; Su, J.N.; Chen, G.H.; Wu, Z.X.; Luo, Y.; Wu, R.T.; Sun, Y.; Lan, T.; Ma, J.Y. The re-emerging of sads-cov infection in pig herds in southern china. Transbound. Emerg. Dis. 2019, 66, 2180–2183. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.L.; Qin, P.; Wang, B.; Liu, Y.; Xu, G.H.; Peng, L.; Zhou, J.; Zhu, S.J.; Huang, Y.W. Broad cross-species infection of cultured cells by bat hku2-related swine acute diarrhea syndrome coronavirus and identification of its replication in murine dendritic cells in vivo highlight its potential for diverse interspecies transmission. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.C. An update on feline infectious peritonitis: Virology and immunopathogenesis. Vet. J. 2014, 201, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipar, A.; Meli, M.L. Feline infectious peritonitis: Still an enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.L.; Liang, Q.Z.; Xu, S.Y.; Mazing, E.; Xu, G.H.; Peng, L.; Qin, P.; Wang, B.; Huang, Y.W. Characterization of a novel bat-hku2-like swine enteric alphacoronavirus (seacov) infection in cultured cells and development of a seacov infectious clone. Virology 2019, 536, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Huang, Y.; Lau, S.K.; Yuen, K.Y. Coronavirus genomics and bioinformatics analysis. Viruses 2010, 2, 1804–1820. [Google Scholar] [CrossRef] [Green Version]
- Snijder, E.J.; Bredenbeek, P.J.; Dobbe, J.C.; Thiel, V.; Ziebuhr, J.; Poon, L.L.; Guan, Y.; Rozanov, M.; Spaan, W.J.; Gorbalenya, A.E. Unique and conserved features of genome and proteome of sars-coronavirus, an early split-off from the coronavirus group 2 lineage. J. Mol. Biol. 2003, 331, 991–1004. [Google Scholar] [CrossRef]
- Kamitani, W.; Huang, C.; Narayanan, K.; Lokugamage, K.G.; Makino, S. A two-pronged strategy to suppress host protein synthesis by sars coronavirus nsp1 protein. Nat. Struct. Mol. Biol. 2009, 16, 1134–1140. [Google Scholar] [CrossRef]
- Tanaka, T.; Kamitani, W.; DeDiego, M.L.; Enjuanes, L.; Matsuura, Y. Severe acute respiratory syndrome coronavirus nsp1 facilitates efficient propagation in cells through a specific translational shutoff of host mrna. J. Virol. 2012, 86, 11128–11137. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, K.; Narayanan, K.; Wada, M.; Popov, V.L.; Cajimat, M.; Baric, R.S.; Makino, S. The endonucleolytic rna cleavage function of nsp1 of middle east respiratory syndrome coronavirus promotes the production of infectious virus particles in specific human cell lines. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Lokugamage, K.G.; Narayanan, K.; Nakagawa, K.; Terasaki, K.; Ramirez, S.I.; Tseng, C.T.; Makino, S. Middle east respiratory syndrome coronavirus nsp1 inhibits host gene expression by selectively targeting mrnas transcribed in the nucleus while sparing mrnas of cytoplasmic origin. J. Virol. 2015, 89, 10970–10981. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Wang, G.; Yang, Y.; Shi, J.; Fang, L.; Li, F.; Xiao, S.; Fu, Z.F.; Peng, G. A conserved region of nonstructural protein 1 from alphacoronaviruses inhibits host gene expression and is critical for viral virulence. J. Biol. Chem. 2019, 294, 13606–13618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Ye, G.; Deng, F.; Wang, G.; Cui, M.; Fang, L.; Xiao, S.; Fu, Z.F.; Peng, G. Structural basis for the inhibition of host gene expression by porcine epidemic diarrhea virus nsp1. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.S.; Johnson, M.A.; Wuthrich, K. NMR assignment of the sars-cov protein nsp1. J. Biomol. NMR 2006, 36 (Suppl. S1), 46. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Lokugamage, K.G.; Rozovics, J.M.; Narayanan, K.; Semler, B.L.; Makino, S. Alphacoronavirus transmissible gastroenteritis virus nsp1 protein suppresses protein translation in mammalian cells and in cell-free hela cell extracts but not in rabbit reticulocyte lysate. J. Virol. 2011, 85, 638–643. [Google Scholar] [CrossRef] [Green Version]
- Zust, R.; Cervantes-Barragan, L.; Kuri, T.; Blakqori, G.; Weber, F.; Ludewig, B.; Thiel, V. Coronavirus non-structural protein 1 is a major pathogenicity factor: Implications for the rational design of coronavirus vaccines. PLoS Pathog 2007, 3, e109. [Google Scholar] [CrossRef] [Green Version]
- Wathelet, M.G.; Orr, M.; Frieman, M.B.; Baric, R.S. Severe acute respiratory syndrome coronavirus evades antiviral signaling: Role of nsp1 and rational design of an attenuated strain. J. Virol. 2007, 81, 11620–11633. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, K.; Huang, C.; Lokugamage, K.; Kamitani, W.; Ikegami, T.; Tseng, C.T.; Makino, S. Severe acute respiratory syndrome coronavirus nsp1 suppresses host gene expression, including that of type i interferon, in infected cells. J. Virol. 2008, 82, 4471–4479. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Liang, R.; Liu, Z.; Shen, Z.; Shi, J.; Shi, Y.; Deng, F.; Xiao, S.; Fu, Z.F.; Peng, G. The n-terminal domain of spike protein is not the enteric tropism determinant for transmissible gastroenteritis virus in piglets. Viruses 2019, 11, 313. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Fang, L.; Yang, H.; Liu, H.; Du, T.; Fang, P.; Wang, D.; Chen, H.; Xiao, S. Isolation, genomic characterization, and pathogenicity of a chinese porcine deltacoronavirus strain chn-hn-2014. Vet. Microbiol. 2016, 196, 98–106. [Google Scholar] [CrossRef]
- Ye, G.; Deng, F.; Shen, Z.; Luo, R.; Zhao, L.; Xiao, S.B.; Fu, Z.F.; Peng, G.Q. Structural basis for the dimerization and substrate recognition specificity of porcine epidemic diarrhea virus 3c-like protease. Virology 2016, 494, 225–235. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Grosse-Kunstleve, R.W.; Hung, L.W.; Ioerger, T.R.; McCoy, A.J.; Moriarty, N.W.; Read, R.J.; Sacchettini, J.C.; Sauter, N.K.; Terwilliger, T.C. Phenix: Building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 2002, 58, 1948–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grell, L.; Parkin, C.; Slatest, L.; Craig, P.A. Ez-viz, a tool for simplifying molecular viewing in pymol. Biochem. Mol. Biol. Educ. 2006, 34, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, G.Y.; Sun, D.J.; Zhang, J.W.; Xie, X.X.; Wu, X.Q.; Fang, W.S.; Tian, J.W.; Yan, C.H.; Wang, H.B.; Fu, F.H. Lx2-32c, a novel semi-synthetic taxane, exerts antitumor activity against prostate cancer cells in vitro and in vivo. Acta Pharm. Sin. B 2017, 7, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Cheng, V.C.; Lau, S.K.; Woo, P.C.; Yuen, K.Y. Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection. Clin. Microbiol. Rev. 2007, 20, 660–694. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.P.; Huo, C.Y.; Xiao, J.; Fan, T.; Zou, S.M.; Qi, P.; Sun, L.Q.; Wang, M.; Hu, Y.X. P-stat1 regulates the influenza a virus replication and inflammatory response in vitro and vivo. Virology 2019, 537, 110–120. [Google Scholar] [CrossRef]
- Ding, L.; Huang, Y.; Dai, M.L.; Zhao, X.M.; Du, Q.; Dong, F.; Wang, L.L.; Huo, R.C.; Zhang, W.L.; Xu, X.G.; et al. Transmissible gastroenteritis virus infection induces cell cycle arrest at s and g2/m phases via p53-dependent pathway. Virus Res. 2013, 178, 241–251. [Google Scholar] [CrossRef]
- Luo, Y.R.; Zhou, S.T.; Yang, L.; Liu, Y.P.; Jiang, S.Y.; Dawuli, Y.; Hou, Y.X.; Zhou, T.X.; Yang, Z.B. Porcine epidemic diarrhoea virus induces cell-cycle arrest through the DNA damage-signalling pathway. J. Vet. Res. 2020, 64, 25–32. [Google Scholar] [CrossRef]
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; et al. Isolation and characterization of viruses related to the sars coronavirus from animals in southern china. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in saudi arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [Green Version]
- Rabaan, A.A.; Al-Ahmed, S.H.; Haque, S.; Sah, R.; Tiwari, R.; Malik, Y.S.; Dhama, K.; Yatoo, M.I.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. Sars-cov-2, sars-cov, and mers-cov: A comparative overview. Infez. Med. 2020, 28, 174–184. [Google Scholar] [PubMed]
- Nelson, M.A.; Macino, G. Structure and expression of the overlapping nd4l and nd5 genes of neurospora crassa mitochondria. Mol. Gen. Genet. 1987, 206, 307–317. [Google Scholar] [CrossRef]
- Cardol, P.; Lapaille, M.; Minet, P.; Franck, F.; Matagne, R.F.; Remacle, C. Nd3 and nd4l subunits of mitochondrial complex i, both nucleus encoded in chlamydomonas reinhardtii, are required for activity and assembly of the enzyme. Eukaryot. Cell 2006, 5, 1460–1467. [Google Scholar] [CrossRef] [Green Version]
- Pilz, A.; Ramsauer, K.; Heidari, H.; Leitges, M.; Kovarik, P.; Decker, T. Phosphorylation of the stat1 transactivating domain is required for the response to type i interferons. EMBO Rep. 2003, 4, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Shi, K.; Yoo, D. Suppression of type i interferon production by porcine epidemic diarrhea virus and degradation of creb-binding protein by nsp1. Virology 2016, 489, 252–268. [Google Scholar] [CrossRef] [Green Version]
- Davies, C.; Brown, C.M.; Westphal, D.; Ward, J.M.; Ward, V.K. Murine norovirus replication induces g0/g1 cell cycle arrest in asynchronously growing cells. J. Virol. 2015, 89, 6057–6066. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Shan, Y.; Zhao, Z.; Chen, J.; Cong, Y. G0/g1 arrest and apoptosis induced by sars-cov 3b protein in transfected cells. Virol. J. 2005, 2, 66. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.L.; Wu, J.; Shan, Y.J.; Yao, Z.Y.; Dong, B.; Chen, B.; Zhao, Z.H.; Wang, S.Q.; Chen, J.P.; Cong, Y.W. Sars coronavirus 7a protein blocks cell cycle progression at g0/g1 phase via the cyclin d3/prb pathway. Virology 2006, 346, 74–85. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SADS-CoV nsp1 | FIPV nsp1 | |
|---|---|---|
| Data collection | ||
| Space group | P212121 | C121 |
| Cell axial lengths (Å) | 54.83, 60.37, 84.00 | 124.42, 86.30, 87.79 |
| Cell angles (°) | 90.00, 90.00, 90.00 | 90.00, 130.75, 90.00 |
| Wavelength | 0.97918 | 0.97918 |
| Resolution range (Å) | 29.19–2.10 | 29.52–1.80 |
| Completeness (%) | 100.0 (100.0) | 98.2 (97.1) |
| Rmerge (last shell) | 0.095 (0.985) | 0.126 (0.435) |
| I/σ (last shell) | 21.20 (2.0) | 15.27 (3.0) |
| Redundancy (last shell) | 12.9 (13.2) | 6.9 (6.7) |
| Refinement | ||
| Resolution (Å) | 29.19–2.10 | 29.52–1.80 |
| Rwork/Rfree | 0.235/0.253 | 0.187/0.207 |
| No. reflections | 16,677 | 63,361 |
| No. of protein atoms | 1560 | 3360 |
| No. of solvent atoms | 69 | 510 |
| No. of ions/ligand | 0 | 0 |
| RMSD | ||
| Bond length (Å) | 0.010 | 0.007 |
| Bond angle (°) | 0.865 | 0.868 |
| B factor (Å2) | 56.48 | 25.76 |
| Ramachandran plot core, allowed, disallowed | 98.45%, 1.55%, 0.00% | 98.77%, 1.23%, 0.00% |
| Gene | F Primer Sequence (5′ to 3′) | R Primer Sequence (5′ to 3′) | Concentration (μM) | Amplicon Sizes (bp) |
|---|---|---|---|---|
| H-STAT1 | CAGACCACAGACAACCTGCT | ACAGAGCCCACTATCCGAGA | 10 | 112 |
| P-STAT1 | GGGCTCTGCTAAAGGACCAG | AGTAAGGTTCGCCTCCGTTC | 10 | 120 |
| F-STAT1 | GGGAACCTTACTTCCACGCA | TCAGCCGCCATGACTTTGTA | 10 | 100 |
| H-ISG15 | GCGCAGATCACCCAGAAGAT | GTTCGTCGCATTTGTCCACC | 10 | 167 |
| P-ISG15 | CTATGAGGTCTGGCTGACGC | GGCTTGAGGTCATACTCCCC | 10 | 147 |
| F-ISG15 | GCAGATCGCCCAGAAAACTG | GACCCTTGTGGTTCCTCACC | 10 | 185 |
| H-E2F2 | TGCCCAGCTACTGCTACCTA | ATCCCCTCCAGATCCAGCTT | 10 | 154 |
| P-E2F2 | GTACCCGCAGACTATGCCTC | AATGCACTTCCCCTTGGGAG | 10 | 187 |
| F-E2F2 | GATCAGTTTCTCCCCGCCTT | AGTTAATCAGCAGGTCCCCG | 10 | 113 |
| H-CDKN1A | ATGTGGACCTGTCACTGTCT | CGTTTGGAGTGGTAGAAATCTGTC | 10 | 189 |
| P-CDKN1A | ATGACCTGGGAGGGGGC | GCACAAGGGTACAAGACAGC | 10 | 105 |
| F-CDKN1A | GGAGCGATGGAACTTCGACT | GGGAGTGAGGCATGAGAGTG | 10 | 232 |
| H-IRF9 | ACCAGGATGCTGCCTTCTTC | CCTGGTGGCAGCAACTGATA | 10 | 268 |
| P-IRF9 | CCCTGCCATCTGGAAGACTC | CCCATTGGTCTCTGCCAACT | 10 | 289 |
| F-IRF9 | TTCTTCAAGGCATGGGCGAT | TTTGTGGTACCCGCATCCTC | 10 | 278 |
| H-GAPDH | CAAATTCCATGGCACCGTCA | GACTCCACGACGTACTCAGC | 10 | 211 |
| P-GAPDH | TCGGAGTGAACGGATTTGGC | TCTCGCTCCTGGAAGATGGT | 10 | 226 |
| F-GAPDH | AAGGTCGGTGTGAACGGATT | TTTGCCGTGGGTGGAATCAT | 10 | 152 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Z.; Yang, Y.; Yang, S.; Zhang, G.; Xiao, S.; Fu, Z.F.; Peng, G. Structural and Biological Basis of Alphacoronavirus nsp1 Associated with Host Proliferation and Immune Evasion. Viruses 2020, 12, 812. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080812
Shen Z, Yang Y, Yang S, Zhang G, Xiao S, Fu ZF, Peng G. Structural and Biological Basis of Alphacoronavirus nsp1 Associated with Host Proliferation and Immune Evasion. Viruses. 2020; 12(8):812. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080812
Chicago/Turabian StyleShen, Zhou, Yiling Yang, Siqi Yang, Guangxu Zhang, Shaobo Xiao, Zhen F. Fu, and Guiqing Peng. 2020. "Structural and Biological Basis of Alphacoronavirus nsp1 Associated with Host Proliferation and Immune Evasion" Viruses 12, no. 8: 812. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080812