
Lack of Evidence of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Spillover in Free-Living Neotropical Non-Human Primates, Brazil

 ,
,  , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
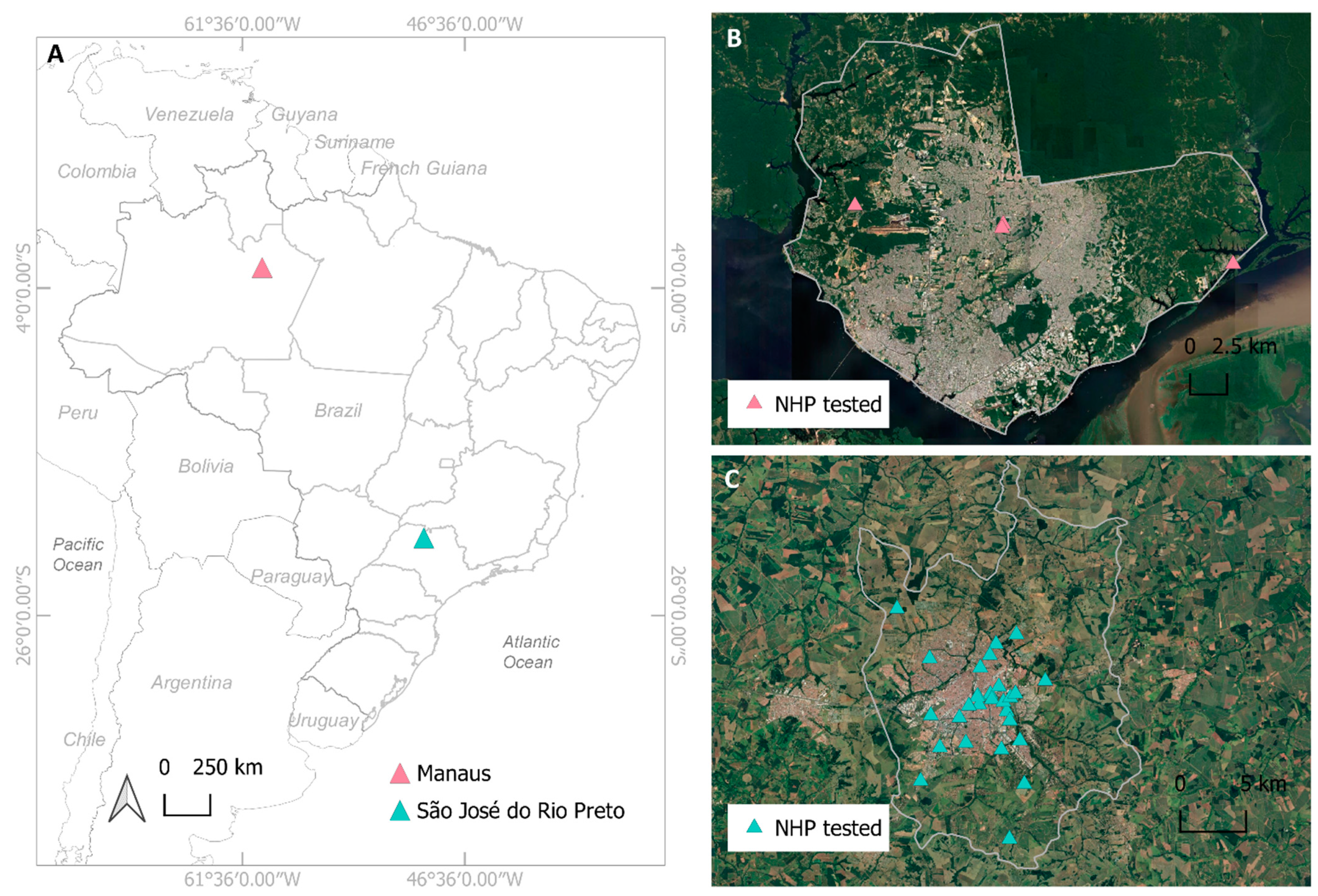
2.1. Study Areas
2.2. Ethics
2.3. Non-Human Primate Sampling
2.4. Virus RNA Extraction and Real Time RT-PCR
2.5. Plaque Reduction Neutralization Test (PRNT)
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 21 September 2021).
- Barlow, P.; van Schalkwyk, M.C.; McKee, M.; Labonté, R.; Stuckler, D. COVID-19 and the collapse of global trade: Building an effective public health response. Lancet Planet. Health 2021, 5, e102–e107. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacharapluesadee, S.; Tan, C.W.; Maneeorn, P.; Duengkae, P.; Zhu, F.; Joyjinda, Y.; Kaewpom, T.; Ni Chia, W.; Ampoot, W.; Lim, B.L.; et al. Evidence for SARS-CoV-2 related coronaviruses circulating in bats and pangolins in Southeast Asia. Nat. Commun. 2021, 12, 1–9. [Google Scholar] [CrossRef]
- Zhou, H.; Ji, J.; Chen, X.; Bi, Y.; Li, J.; Wang, Q.; Hu, T.; Song, H.; Zhao, R.; Chen, Y.; et al. Identification of novel bat coronaviruses sheds light on the evolutionary origins of SARS-CoV-2 and related viruses. Cell 2021. [Google Scholar] [CrossRef]
- Guth, S.; Hanley, K.A.; Althouse, B.M.; Boots, M. Ecological processes underlying the emergence of novel enzootic cycles: Arboviruses in the neotropics as a case study. PLoS Neglected Trop. Dis. 2020, 14, e0008338. [Google Scholar] [CrossRef] [PubMed]
- Mahdy, M.A.A.; Younis, W.; Ewaida, Z. An Overview of SARS-CoV-2 and Animal Infection. Front. Vet. Sci. 2020, 7, 596391. [Google Scholar] [CrossRef]
- USDA. Cases of SARS-CoV-2 in Animals in the Unites States. Available online: https://www.aphis.usda.gov/aphis/dashboards/tableau/sars-dashboard (accessed on 2 July 2021).
- Fischhoff, I.R.; Castellanos, A.A.; Rodrigues, J.; Varsani, A.; Han, B.A. Predicting the zoonotic capacity of mammal species for SARS-CoV-2. bioRxiv 2021. [Google Scholar] [CrossRef]
- Fenollar, F.; Mediannikov, O.; Maurin, M.; Devaux, C.; Colson, P.; Levasseur, A.; Fournier, P.-E.; Raoult, D. Mink, SARS-CoV-2, and the Human-Animal Interface. Front. Microbiol. 2021, 12, 663815. [Google Scholar] [CrossRef]
- Aguiló-Gisbert, J.; Padilla-Blanco, M.; Lizana, V.; Maiques, E.; Muñoz-Baquero, M.; Chillida-Martínez, E.; Cardells, J.; Rubio-Guerri, C. First Description of SARS-CoV-2 Infection in Two Feral American Mink (Neovison vison) Caught in the Wild. Animals 2021, 11, 1422. [Google Scholar] [CrossRef] [PubMed]
- van Aart, A.E.; Velkers, F.C.; Fischer, E.A.J.; Broens, E.M.; Egberink, H.; Zhao, S.; Engelsma, M.; Hakze-van der Honing, R.W.; Harders, F.; de Rooij, M.M.T.; et al. SARS-CoV-2 infection in cats and dogs in infected mink farms. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Hamer, S.A.; Ghai, R.R.; Zecca, I.B.; Auckland, L.D.; Roundy, C.M.; Davila, E.; Busselman, R.E.; Tang, W.; Pauvolid-Correa, A.; Killian, M.L.; et al. SARS-CoV-2 B.1.1.7 variant of concern detected in a pet dog and cat after exposure to a person with COVID-19, USA. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.S.; Abdelwhab, E.M. Evidence for SARS-CoV-2 Infection of Animal Hosts. Pathogens 2020, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.; Dantas, W.; Nascimento, J.D.; da Silva, M.; de Oliveira, R.; Pena, L. In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic. Viruses 2021, 13, 379. [Google Scholar] [CrossRef]
- Pandey, K.; Acharya, A.; Mohan, M.; Ng, C.L.; Reid, S.P.; Byrareddy, S.N. Animal models for SARS-CoV-2 research: A comprehensive literature review. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Singh, D.K.; Singh, B.; Ganatra, S.R.; Gazi, M.; Cole, J.; Thippeshappa, R.; Alfson, K.J.; Clemmons, E.; Gonzalez, O.; Escobedo, R.; et al. Responses to acute infection with SARS-CoV-2 in the lungs of rhesus macaques, baboons and marmosets. Nat. Microbiol. 2021, 6, 73–86. [Google Scholar] [CrossRef]
- Rockx, B.; Kuiken, T.; Herfst, S.; Bestebroer, T.; Lamers, M.M.; Munnink, B.B.O.; De Meulder, D.; Van Amerongen, G.; Brand, J.V.D.; Okba, N.M.A.; et al. Comparative pathogenesis of COVID-19, MERS, and SARS in a nonhuman primate model. Science 2020, 368, 1012–1015. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Zhao, Y.; Yu, W.; Yang, Y.; Gao, J.; Wang, J.; Kuang, D.; Yang, M.; Yang, J.; Ma, C.; et al. Comparison of nonhuman primates identified the suitable model for COVID-19. Signal Transduct. Target. Ther. 2020, 5, 1–9. [Google Scholar] [CrossRef]
- van Doremalen, N.; Munster, V.J. Animal models of Middle East respiratory syndrome coronavirus infection. Antivir. Res. 2015, 122, 28–38. [Google Scholar] [CrossRef]
- Greenough, T.C.; Carville, A.; Coderre, J.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Mansfield, K. Pneumonitis and Multi-Organ System Disease in Common Marmosets (Callithrix jacchus) Infected with the Severe Acute Respiratory Syndrome-Associated Coronavirus. Am. J. Pathol. 2005, 167, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Fischhoff, I.; Castellanos, A.; Rodrigues, J.; Varsani, A.; Han, B.A. Supplementary Information for Fischhoff et al. 2021 Predicting the Zoonotic Capacity of Mammal Species for SARS-CoV-2. Available online: https://0-doi-org.brum.beds.ac.uk/10.25390/caryinstitute.c.5293339.v4 (accessed on 3 July 2021).
- Sharun, K.; Dhama, K.; Pawde, A.M.; Gortázar, C.; Tiwari, R.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J.; de la Fuente, J.; Michalak, I.; Attia, Y.A. SARS-CoV-2 in animals: Potential for unknown reservoir hosts and public health implications. Vet. Q. 2021, 41, 181–201. [Google Scholar] [CrossRef] [PubMed]
- Milhim, B.H.G.A.; Estofolete, C.F.; Da Rocha, L.C.; Liso, E.; Brienze, V.M.S.; Vasilakis, N.; Terzian, A.C.B.; Nogueira, M.L. Fatal Outcome of Ilheus Virus in the Cerebrospinal Fluid of a Patient Diagnosed with Encephalitis. Viruses 2020, 12, 957. [Google Scholar] [CrossRef] [PubMed]
- Domingo, C.; Patel, P.; Yillah, J.; Weidmann, M.; Méndez, J.A.; Nakouné, E.R.; Niedrig, M. Advanced Yellow Fever Virus Genome Detection in Point-of-Care Facilities and Reference Laboratories. J. Clin. Microbiol. 2012, 50, 4054–4060. [Google Scholar] [CrossRef] [Green Version]
- Vasilakis, N.; Tesh, R.B.; Durbin, A.P.; Munoz-Jordan, J.L.; Da Rosa, A.P.A.T.; Weaver, S.C. Antigenic relationships between sylvatic and endemic dengue viruses. Am. J. Trop. Med. Hyg. 2008, 79, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.B.; Machado, R.R.G.; Amgarten, D.E.; Malta, F.D.M.; De Araujo, G.G.; Monteiro, C.O.; Candido, E.D.; Soares, C.P.; De Menezes, F.G.; Pires, A.C.C.; et al. SARS-CoV-2 isolation from the first reported patients in Brazil and establishment of a coordinated task network. Memórias do Instituto Oswaldo Cruz 2020, 115, e200342. [Google Scholar] [CrossRef]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/region/amro/country/br (accessed on 4 July 2021).
- Castro, M.C.; Kim, S.; Barberia, L.; Ribeiro, A.F.; Gurzenda, S.; Ribeiro, K.B.; Abbott, E.; Blossom, J.; Rache, B.; Singer, B.H. Spatiotemporal pattern of COVID-19 spread in Brazil. Science 2021, 372, 821–826. [Google Scholar] [CrossRef]
- Sao Paulo Governo do Estado Saude. SP Contra o Novo Coronavírus—Boletim Completo. Available online: https://www.seade.gov.br/coronavirus/ (accessed on 23 July 2021).
- Fundacao de Vigilancia em Saude do Amazonas. Painel de Monitoramento da COVID-19. Available online: https://www.fvs.am.gov.br/indicadorSalaSituacao_view/60/2 (accessed on 23 July 2021).
- Boehm, E.; Kronig, I.; Neher, R.A.; Eckerle, I.; Vetter, P.; Kaiser, L.; Geneva Centre for Emerging Viral, D. Novel SARS-CoV-2 variants: The pandemics within the pandemic. Clin. Microbiol. Infect. 2021. [Google Scholar] [CrossRef]
- Imai, M.; Halfmann, P.J.; Yamayoshi, S.; Iwatsuki-Horimoto, K.; Chiba, S.; Watanabe, T.; Nakajima, N.; Ito, M.; Kuroda, M.; Kiso, M.; et al. Characterization of a new SARS-CoV-2 variant that emerged in Brazil. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Bartlett, S.L.; Diel, D.G.; Wang, L.; Zec, S.; Laverack, M.; Martins, M.; Caserta, L.C.; Killian, M.L.; Terio, K.; Olmstead, C.; et al. SARS-CoV-2 infection and longitudinal fecal screening in malayan tigers (panthera tigris jacksoni), amur tigers (panthera tigris altaica ), and african lions (panthera leo krugeri) at the bronx zoo, new york, usa. J. Zoo Wildl. Med. 2021, 51, 733–744. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D.; et al. From People to Panthera: Natural SARS-CoV-2 Infection in Tigers and Lions at the Bronx Zoo. mBio 2020, 11, e02220-20. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Martins, M.; Falkenberg, S.; Buckley, A.; Caserta, L.C.; Mitchell, P.K.; Cassmann, E.D.; Rollins, A.; Zylich, N.C.; Renshaw, R.W.; et al. Susceptibility of white-tailed deer (Odocoileus virginianus) to SARS-CoV-2. J. Virol. 2021, 95, e00083-21. [Google Scholar] [CrossRef] [PubMed]
- Watsa, M.E. Wildlife Disease Surveillance Focus Group Rigorous wildlife disease surveillance. Science 2020, 369, 145–147. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| ID | Genus | Neighborhood | Date | SARS-CoV-2 RT-qPCR |
|---|---|---|---|---|
| NHP21/01 | Callithrix | Engenheiro Schimdt | 03/16/2020 | Neg |
| NHP21/02 | Callithrix | Vila Elvira | 03/26/2020 | Neg |
| NHP21/03 | Callithrix | Central | 04/17/2020 | Neg |
| NHP21/04 | Callithrix | Central | 06/01/2020 | Neg |
| NHP21/05 | Callithrix | Jaguaré | 06/08/2020 | Neg |
| NHP21/06 | Callithrix | Cidade Jardim | 06/16/2020 | Neg |
| NHP21/07 | Callithrix | Vila Toninho | 08/25/2020 | Neg |
| NHP21/08 | Callithrix | Americano | 08/27/2020 | Neg |
| NHP21/09 | Callithrix | Vila Toninho | 09/08/2020 | Neg |
| NHP21/10 | Callithrix | Caic | 09/24/2020 | Neg |
| NHP21/11 | Callithrix | Vila Toninho | 10/02/2020 | Neg |
| NHP21/12 | Callithrix | Solo Sagrado | 10/06/2020 | Neg |
| NHP21/13 | Callithrix | Vila Toninho | 10/21/2020 | Neg |
| NHP21/14 | Callithrix | Jaguaré | 10/21/2020 | Neg |
| NHP21/15 | Callithrix | Jaguaré | 10/23/2020 | Neg |
| NHP21/16 | Callithrix | Jaguaré | 10/26/2020 | Neg |
| NHP21/17 | Callithrix | Solo Sagrado | 10/27/2020 | Neg |
| NHP21/18 | Callithrix | São Francisco | 11/06/2020 | Neg |
| NHP21/19 | Callithrix | Anchieta | 11/09/2020 | Neg |
| NHP21/20 | Alouatta | Solo Sagrado | 11/09/2020 | Neg |
| NHP21/21 | Callicebus | São Deocleciano | 11/11/2020 | Neg |
| NHP21/22 | Callithrix | Americano | 11/13/2020 | Neg |
| NHP21/23 | Callithrix | Americano | 11/13/2020 | Neg |
| NHP21/24 | Callithrix | São Francisco | 11/17/2020 | Neg |
| NHP21/25 | Callithrix | Central | 12/10/2020 | Neg |
| NHP21/26 | Callithrix | Novo Horizonte | 09/22/2020 | Neg |
| NHP21/27 | Alouatta | Jose Bonifácio | 10/05/2020 | Neg |
| NHP21/28 | Alouatta | Tabapuã | 10/26/2020 | Neg |
| NHP21/29 | Callithrix | Parque Industrial | 02/03/2021 | Neg |
| NHP21/30 | Callithrix | Americano | 02/11/2021 | Neg |
| NHP21/31 | Callithrix | São Francisco | 02/17/2021 | Neg |
| NHP21/32 | Callithrix | Central | 03/11/2021 | Neg |
| NHP21/33 | Callithrix | Central | 03/18/2021 | Neg |
| NHP21/34 | Callithrix | Central | 03/18/2021 | Neg |
| ID | Genera/Specie | Neighborhood | Date | PRNT Titer |
|---|---|---|---|---|
| H111P-SB | Saguinus bicolor | Sumauma | 10/11/2020 | <20 |
| H114P-SB | Saguinus bicolor | Sumauma | 10/11/2020 | <20 |
| H115P-SB | Saguinus bicolor | Sumauma | 10/11/2020 | <20 |
| H116P-SB | Saguinus bicolor | Sumauma | 10/11/2020 | <20 |
| H117P-SB | Saguinus bicolor | Sumauma | 10/11/2020 | <20 |
| H128P-SB | Saguinus bicolor | Puraquequara | 21/07/2020 | <20 |
| H129P-SB | Saguinus bicolor | Puraquequara | 21/07/2020 | <20 |
| H130P-SB | Saguinus bicolor | Puraquequara | 21/07/2020 | <20 |
| H131P-SB | Saguinus bicolor | Puraquequara | 21/07/2020 | <20 |
| H132P-SB | Saguinus bicolor | Taruma | 11/08/2020 | <20 |
| H133P-SB | Saguinus bicolor | Taruma | 11/08/2020 | <20 |
| H134P-SB | Saguinus bicolor | Taruma | 11/08/2020 | <20 |
| H135P-SB | Saguinus bicolor | Taruma | 11/08/2020 | <20 |
| H136P-SB | Saguinus bicolor | Taruma | 11/08/2020 | <20 |
| H137P-SB | Saguinus bicolor | Taruma | 11/08/2020 | <20 |
| H138P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H139P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H140P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H141P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H142P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H143P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H144P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H145P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H146P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H147P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
| H148P-SB | Saguinus bicolor | Sumauma | 09/11/2020 | <20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sacchetto, L.; Chaves, B.A.; Costa, E.R.; de Menezes Medeiros, A.S.; Gordo, M.; Araújo, D.B.; Oliveira, D.B.L.; da Silva, A.P.B.; Negri, A.F.; Durigon, E.L.; et al. Lack of Evidence of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Spillover in Free-Living Neotropical Non-Human Primates, Brazil. Viruses 2021, 13, 1933. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101933
Sacchetto L, Chaves BA, Costa ER, de Menezes Medeiros AS, Gordo M, Araújo DB, Oliveira DBL, da Silva APB, Negri AF, Durigon EL, et al. Lack of Evidence of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Spillover in Free-Living Neotropical Non-Human Primates, Brazil. Viruses. 2021; 13(10):1933. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101933
Chicago/Turabian StyleSacchetto, Lívia, Bárbara Aparecida Chaves, Edson Rodrigues Costa, Aline Souza de Menezes Medeiros, Marcelo Gordo, Danielle Bastos Araújo, Danielle Bruna Leal Oliveira, Ana Paula Betaressi da Silva, Andréia Francesli Negri, Edison Luiz Durigon, and et al. 2021. "Lack of Evidence of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Spillover in Free-Living Neotropical Non-Human Primates, Brazil" Viruses 13, no. 10: 1933. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101933