
An Isoform of the Eukaryotic Translation Elongation Factor 1A (eEF1a) Acts as a Pro-Viral Factor Required for Tomato Spotted Wilt Virus Disease in Nicotiana benthamiana

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viral Strains and Plant Material
2.2. RNP Purification from TSWV-Infected N. benthamiana
2.3. S. cerevisiae Expressing a TSWV Replicon System
2.4. In-Gel Tryptic Digestion and Mass-Spectrometry (MS) Analysis
2.5. Functional Enrichment Analyses and eEF1A Phylogeny
2.6. Virus-Induced Gene Silencing and Serological Detection of TSWV
2.7. Gene Expression Analysis Using Real Time RT-PCR
3. Results
3.1. TSWV RNPs Isolated from N. benthamiana Contain Proteins Implicated in Plant Defence Responses and Solute Transport
3.2. The TSWV RNP-Resident N. benthamiana Proteins Display an Overrepresentation for GO Terms Implicated in Immune System Process, Protein Folding, and Transport
3.3. TSWV RNPs Produced in Yeast Associate with Cellular Proteins Involved in Transcription and Translation
3.4. The Yeast TSWV-like RNP Interactome Is Overrepresented for GO Terms Associated with Translation and Ribosome Biogenesis
3.5. TSWV RNPomes Purified from Yeast and N. benthamiana Contain Conserved Proteins Potentially Involved in Viral Replication
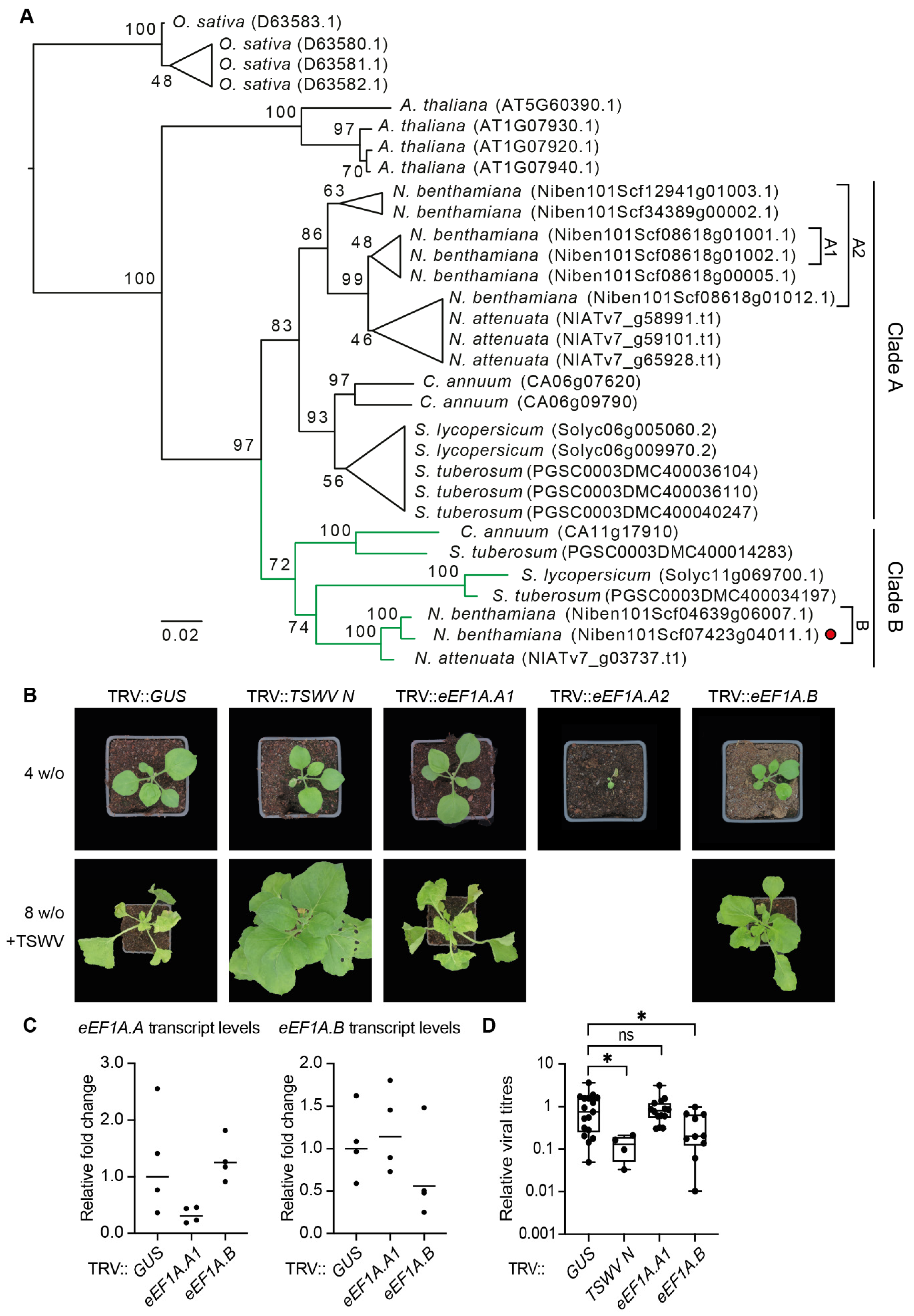
3.6. Functional Studies on the TSWV RNP Interactome Mark an Isoform of the Eukaryotic Translation Elongation Factor 1A as a Pro-Viral Factor of TSWV
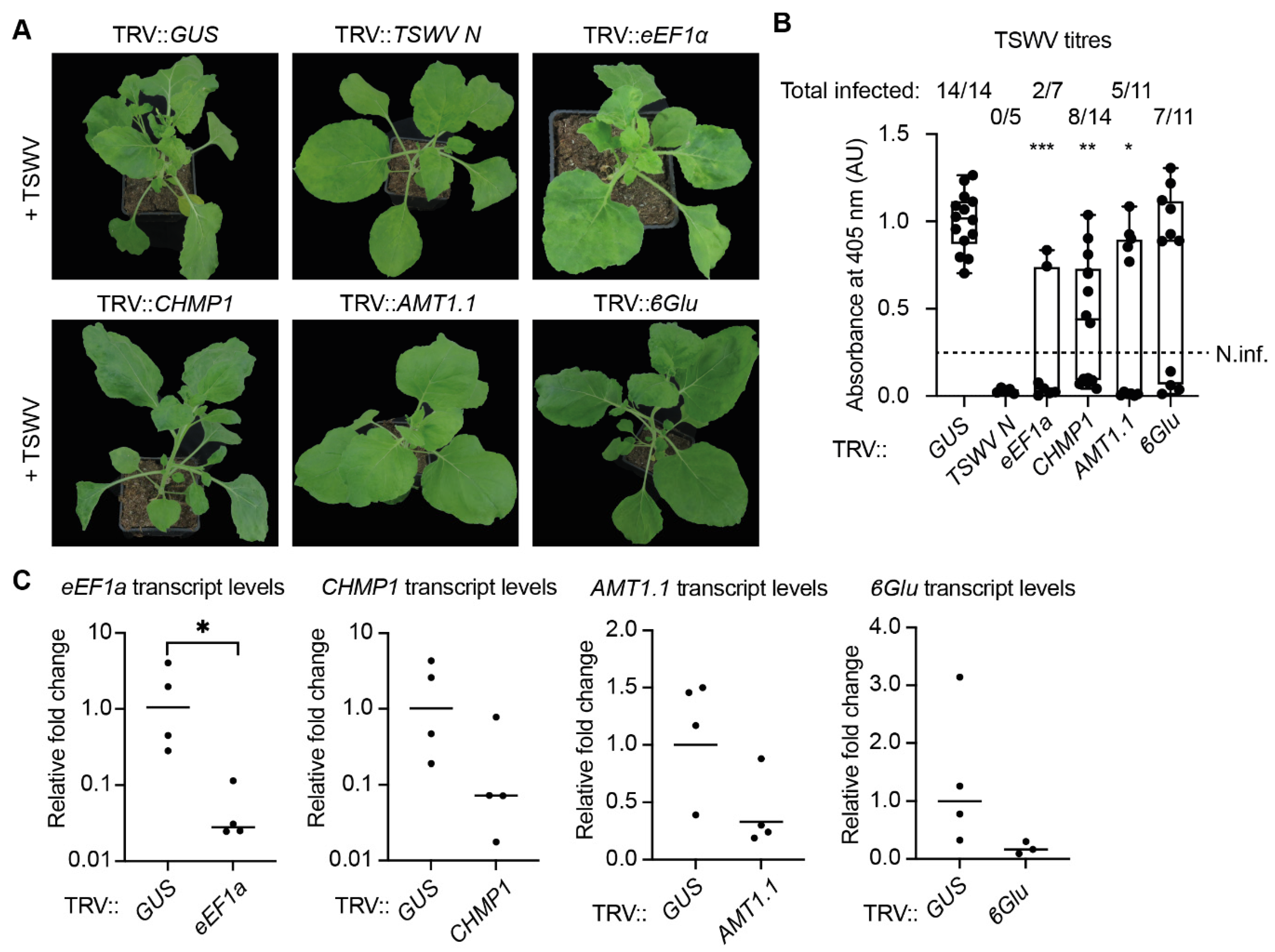
3.7. Silencing of One Specific Isoforms of the eEF1A Gene Family in N. benthamiana Is Sufficient for Loss of TSWV Susceptibility
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parrella, G.; Gognalons, P.; Gebre, S.; Xe, K.; Vovlas, C.; Marchoux, G. An update of the host range of Tomato spotted wilt virus. J. Plant Pathol. 2003, 85, 227–264. [Google Scholar]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Tospovirus-thrips interactions. Annu. Rev. Phytopathol. 2005, 43, 459–489. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J. The dependence of viral RNA replication on co-opted host factors. Nat. Rev. Microbiol. 2012, 10, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef] [PubMed]
- Van Schie, C.C.N.; Takken, F.L.W. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Hashimoto, M.; Neriya, Y.; Yamaji, Y.; Namba, S. Recessive resistance to plant viruses: Potential resistance genes beyond translation initiation factors. Front. Microbiol. 2016, 7, 1695. [Google Scholar] [CrossRef] [Green Version]
- García-Marcos, A.; Pacheco, R.; Manzano, A.; Aguilar, E.; Tenllado, F. Oxylipin biosynthesis genes positively regulate programmed cell death during compatible infections with the synergistic pair Potato virus X-Potato virus Y and Tomato spotted wilt virus. J. Virol. 2013, 87, 5769–5783. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Xue, F.; Xu, M.; Chen, X.; Zhao, W.; Garcia-Murria, M.J.; Mingarro, I.; Liu, Y.; Huang, Y.; Jiang, L.; et al. The ER-membrane transport system is critical for intercellular trafficking of the NSm movement protein and Tomato spotted wilt tospovirus. PLoS Pathog. 2016, 12, e1005443. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Xiang, Q.; Yang, T.; Ma, H.; Ding, X.; Tao, X. Molecular co-chaperone SGT1 is critical for cell-to-cell movement and systemic infection of Tomato spotted wilt virus in Nicotiana benthamiana. Viruses 2018, 10, 647. [Google Scholar] [CrossRef] [Green Version]
- Kormelink, R. The molecular biology of Tospoviruses and resistance strategies. In Bunyaviridae: Molecular and Cellular Biology, 1st ed.; Plyusnin, A., Elliott, R.M., Eds.; Caister Academic Press: New York, NY, USA, 2011; pp. 163–191. [Google Scholar]
- Mohamed, N.A. Isolation and characterization of subviral structures from Tomato spotted wilt virus. J. Gen. Virol. 1981, 53, 197–206. [Google Scholar] [CrossRef]
- Kikkert, M.; van Poelwijk, F.; Storms, M.; Kassies, W.; Bloksma, H.; van Lent, J.; Kormelink, R.; Goldbach, R. A protoplast system for studying Tomato spotted wilt virus infection. J. Gen. Virol. 1997, 78, 1755–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terret-Welter, Z.; Bonnet, G.; Moury, B.; Gallois, J.-L. Analysis of Tomato spotted wilt virus RNA-dependent RNA polymerase adaptative evolution and constrained domains using homology protein structure modelling. J. Gen. Virol. 2020, 101, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, R.; de Haan, P.; Peters, D.; Goldbach, R. Viral RNA synthesis in Tomato spotted wilt virus-infected Nicotiana rustica plants. J. Gen. Virol. 1992, 73, 687–693. [Google Scholar] [CrossRef]
- Kormelink, R.; Storms, M.; van Lent, J.; Peters, D.; Goldbach, R. Expression and subcellular location of the NSm protein of Tomato spotted wilt virus (TSWV), a putative viral movement protein. Virology 1994, 200, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Borst, J.W.; Goldbach, R.; Kormelink, R. Tomato spotted wilt virus nucleocapsid protein interacts with both viral glycoproteins GN and GC in planta. Virology 2009, 383, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, D.; Goldbach, R.; Kormelink, R. Requirements for ER-arrest and sequential exit to the golgi of Tomato spotted wilt virus glycoproteins. Traffic 2009, 10, 664–672. [Google Scholar] [CrossRef]
- Ribeiro, D.; Jung, M.; Moling, S.; Borst, J.W.; Goldbach, R.; Kormelink, R. The cytosolic nucleoprotein of the plant-infecting bunyavirus Tomato spotted wilt recruits endoplasmic reticulum-resident proteins to endoplasmic reticulum export sites. Plant Cell 2013, 25, 3602–3614. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, P.; Malet, H.; Cusack, S.; Reguera, J. Structural insights into bunyavirus replication and its regulation by the vRNA promoter. Cell 2015, 161, 1267–1279. [Google Scholar] [CrossRef] [Green Version]
- Ferron, F.; Weber, F.; de la Torre, J.C.; Reguera, J. Transcription and replication mechanisms of Bunyaviridae and Arenaviridae L proteins. Virus Res. 2017, 234, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Van Knippenberg, I.; Goldbach, R.; Kormelink, R. Purified Tomato spotted wilt virus particles support both genome replication and transcription in vitro. Virology 2002, 303, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Raju, R.; Kolakofsky, D. Translational requirement of La Crosse virus S-mRNA synthesis: In vivo studies. J. Virol. 1987, 61, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barr, J.N. Bunyavirus mRNA synthesis is coupled to translation to prevent premature transcription termination. RNA 2007, 13, 731–736. [Google Scholar] [CrossRef]
- Van Knippenberg, I.; Goldbach, R.; Kormelink, R. In vitro transcription of Tomato spotted wilt virus is independent of translation. J. Gen. Virol. 2004, 85, 1335–1338. [Google Scholar] [CrossRef]
- Komoda, K.; Ishibashi, K.; Kawamura-Nagaya, K.; Ishikawa, M. Possible involvement of eEF1A in Tomato spotted wilt virus RNA synthesis. Virology 2014, 468–470, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, P.D. Yeast as a model host to explore plant virus-host interactions. Annu. Rev. Phytopathol. 2008, 46, 217–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, K.; Matsumoto-Yokoyama, E.; Ishikawa, M.A. Tomato spotted wilt virus S RNA-based replicon system in yeast. Sci. Rep. 2017, 7, 12647. [Google Scholar] [CrossRef] [PubMed]
- Adkins, S.; Quadt, R.; Choi, T.-J.; Ahlquist, P.; German, T. An RNA-dependent RNA polymerase activity associated with virions of Tomato spotted wilt virus, a plant- and insect-infecting Bunyavirus. Virology 1995, 207, 308–311. [Google Scholar] [CrossRef] [Green Version]
- De Ávila, A.C.; de Haan, P.; Smeets, M.L.L.; Resende, R.D.O.; Kormelink, R.; Kitajima, E.W.; Goldbach, R.W.; Peters, D. Distinct levels of relationships between tospovirus isolates. Arch. Virol. 1993, 128, 211–227. [Google Scholar] [CrossRef] [PubMed]
- Inoue-Nagata, A.K.; Kormelink, R.; Nagata, T.; Kitajima, E.W.; Goldbach, R.; Peters, D. Effects of temperature and host on the generation of Tomato spotted wilt virus defective interfering RNAs. Phytopathology 1997, 87, 1168–1173. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to Tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Tas, P.W.L.; Boerjan, M.L.; Peters, D. Purification and serological analysis of Tomato spotted wilt virus. Neth. J. Plant Pathol. 1977, 83, 61–72. [Google Scholar] [CrossRef]
- Griep, R.A.; Prins, M.; van Twisk, C.; Keller, H.J.H.G.; Kerschbaumer, R.J.; Kormelink, R.; Goldbach, R.W.; Schots, A. Application of phage display in selecting Tomato spotted wilt virus-specific single-chain antibodies (scFvs) for sensitive diagnosis in ELISA. Phytopathology 2000, 90, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Grefen, C. The split-ubiquitin system for the analysis of three-component interactions. In Arabidopsis Protocols, Sanchez-Serrano, J.J., Salinas, J., Eds.; Humana Press: Totowa, NJ, USA, 2014; pp. 659–678. [Google Scholar]
- Maio, F.; Helderman, T.A.; Arroyo-Mateos, M.; van der Wolf, M.; Boeren, S.; Prins, M.; van den Burg, H.A. Identification of tomato proteins that interact with Replication initiator protein (Rep) of the Geminivirus TYLCV. Front. Plant Sci. 2020, 11, 1069. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Rappsilber, J.; Andersen, J.S.; Mann, M. Microcolumns with self-assembled particle frits for proteomics. J. Chromatogr. A 2002, 979, 233–239. [Google Scholar] [CrossRef]
- Humphrey, S.J.; Crossett, B.; Parker, B.L. NanoBlow: A simple device to limit contaminants during nanoLC-MS. J. Proteome Res. 2019, 18, 3219–3222. [Google Scholar] [CrossRef]
- Hubner, N.C.; Bird, A.W.; Cox, J.; Splettstoesser, B.; Bandilla, P.; Poser, I.; Hyman, A.; Mann, M. Quantitative proteomics combined with BAC TransgeneOmics reveals in vivo protein interactions. J. Cell Biol. 2010, 189, 739–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, K.; van Lent, J.W.M.; Boeren, S.; Fang, M.; Theilmann, D.A.; Erlandson, M.A.; Vlak, J.M.; van Oers, M.M. Characterization of novel components of the Baculovirus infectivity factor complex. J. Virol. 2012, 86, 4981–4988. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. EggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, msab293. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, 465–469. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Pozo, N.; Rosli, H.G.; Martin, G.B.; Mueller, L.A. The SGN VIGS Tool: User-friendly software to design virus-induced gene silencing (VIGS) constructs for functional genomics. Mol. Plant 2015, 8, 486–488. [Google Scholar] [CrossRef] [Green Version]
- Tameling, W.I.L.; Baulcombe, D.C. Physical association of the NB-LRR resistance protein Rx with a Ran GTPase-activating protein is required for extreme resistance to Potato virus X. Plant Cell 2007, 19, 1682–1694. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- De Ávila, A.C.; Huguenot, C.; Resende, R.D.O.; Kitajima, E.W.; Goldbach, R.W.; Peters, D. Serological differentiation of 20 isolates of Tomato spotted wilt virus. J. Gen. Virol. 1990, 71, 2801–2807. [Google Scholar] [CrossRef]
- Liu, D.; Shi, L.; Han, C.; Yu, J.; Li, D.; Zhang, Y. Validation of reference genes for gene expression studies in virus-infected Nicotiana benthamiana using quantitative real-time PCR. PLoS ONE 2012, 7, e46451. [Google Scholar] [CrossRef]
- Soellick, T.-R.; Uhrig, J.F.; Bucher, G.L.; Kellmann, J.-W.; Schreier, P.H. The movement protein NSm of Tomato spotted wilt tospovirus (TSWV): RNA binding, interaction with the TSWV N protein, and identification of interacting plant proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 2373–2378. [Google Scholar] [CrossRef] [Green Version]
- German, T.L.; Adkins, S.; Witherell, A.; Richmond, K.E.; Knaack, W.R.; Willis, D.K. Infection of Arabidopsis thaliana ecotype columbia by Tomato spotted wilt virus. Plant Mol. Biol. Report. 1995, 13, 110–117. [Google Scholar] [CrossRef]
- Padmanabhan, C.; Ma, Q.; Shekasteband, R.; Stewart, K.S.; Hutton, S.F.; Scott, J.W.; Fei, Z.; Ling, K.-S. Comprehensive transcriptome analysis and functional characterization of PR-5 for its involvement in tomato Sw-7 resistance to tomato spotted wilt tospovirus. Sci. Rep. 2019, 9, 7673. [Google Scholar] [CrossRef]
- Xu, M.; Chen, J.; Huang, Y.; Shen, D.; Sun, P.; Xu, Y.; Tao, X. Dynamic transcriptional profiles of Arabidopsis thaliana infected by Tomato spotted wilt virus. Phytopathology 2019, 110, 153–163. [Google Scholar] [CrossRef]
- Zhai, Y.; Gnanasekaran, P.; Pappu, H.R. Identification and characterization of plant-interacting targets of Tomato spotted wilt virus silencing suppressor. Pathogens 2021, 10, 27. [Google Scholar] [CrossRef]
- Olesinski, A.A.; Almon, E.; Navot, N.; Perl, A.; Galun, E.; Lucas, W.J.; Wolf, S. Tissue-specific expression of the Tobacco mosaic virus movement protein in transgenic potato plants alters plasmodesmal function and carbohydrate partitioning. Plant Physiol. 1996, 111, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Von Bargen, S.; Salchert, K.; Paape, M.; Piechulla, B.; Kellmann, J.-W. Interactions between the Tomato spotted wilt virus movement protein and plant proteins showing homologies to myosin, kinesin and DnaJ-like chaperones. Plant Physiol. Biochem. 2001, 39, 1083–1093. [Google Scholar] [CrossRef]
- Hofius, D.; Maier, A.T.; Dietrich, C.; Jungkunz, I.; Börnke, F.; Maiss, E.; Sonnewald, U. Capsid protein-mediated recruitment of host DnaJ-Like proteins is required for Potato virus Y infection in tobacco plants. J. Virol. 2007, 81, 11870–11880. [Google Scholar] [CrossRef] [Green Version]
- Marondedze, C.; Thomas, L.; Serrano, N.L.; Lilley, K.S.; Gehring, C. The RNA-binding protein repertoire of Arabidopsis thaliana. Sci. Rep. 2016, 6, 29766. [Google Scholar] [CrossRef] [Green Version]
- Beckmann, B.M.; Horos, R.; Fischer, B.; Castello, A.; Eichelbaum, K.; Alleaume, A.-M.; Schwarzl, T.; Curk, T.; Foehr, S.; Huber, W.; et al. The RNA-binding proteomes from yeast to man harbour conserved enigmRBPs. Nat. Commun. 2015, 6, 10127. [Google Scholar] [CrossRef]
- Zhong, T.; Arndt, K.T. The yeast SIS1 protein, a DnaJ homolog, is required for the initiation of translation. Cell 1993, 73, 1175–1186. [Google Scholar] [CrossRef]
- Axelos, M.; Bardet, C.; Liboz, T.; le van Thai, A.; Curie, C.; Lescure, B. The gene family encoding the Arabidopsis thaliana translation elongation factor EF-1α: Molecular cloning, characterization and expression. MGG 1989, 219, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Kidou, S.-I.; Ejiri, S.-I. Isolation, characterization and mRNA expression of four cDNAs encoding translation elongation factor 1A from rice (Oryza sativa L.). Plant Mol. Biol. 1998, 36, 137–148. [Google Scholar] [CrossRef]
- Yamaji, Y.; Sakurai, K.; Hamada, K.; Komatsu, K.; Ozeki, J.; Yoshida, A.; Yoshii, A.; Shimizu, T.; Namba, S.; Hibi, T. Significance of eukaryotic translation elongation factor 1A in Tobacco mosaic virus infection. Arch. Virol. 2010, 155, 263–268. [Google Scholar] [CrossRef]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The many roles of the eukaryotic elongation factor 1 complex. WIRES RNA 2012, 3, 543–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeenko, V.V.; Ryabova, L.A.; Spirin, A.S.; Rothnie, H.M.; Hess, D.; Browning, K.S.; Hohn, T. Eukaryotic elongation factor 1A interacts with the upstream pseudoknot domain in the 3′ untranslated region of Tobacco mosaic virus RNA. J. Virol. 2002, 76, 5678–5691. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, D.; Dreher, T.W. The tRNA-like structure of Turnip yellow mosaic virus RNA is a 3′-translational enhancer. Virology 2004, 321, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Pogany, J.; Tupman, S.; Esposito, A.M.; Kinzy, T.G.; Nagy, P.D. Translation elongation factor 1A facilitates the assembly of the Tombusvirus replicase and stimulates minus-strand synthesis. PLoS Pathog. 2010, 6, e1001175. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.; Oh, C.-S.; Kang, B.-C. Translation elongation factor 1B (eEF1B) is an essential host factor for Tobacco mosaic virus infection in plants. Virology 2013, 439, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wei, T.; Abbott, C.M.; Harrich, D. The unexpected roles of eukaryotic translation elongation factors in RNA virus replication and pathogenesis. Microbiol. Mol. Biol. Rev. 2013, 77, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Valenzuela, J.A.; Gibbon, B.C.; Hughes, P.A.; Dreher, T.W.; Larkins, B.A. eEF1A isoforms change in abundance and actin-binding activity during Maize endosperm development. Plant Physiol. 2003, 133, 1285–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, S.-M.; Chen, L.; Lambertson, D.; Anand, M.; Kinzy Terri, G.; Madura, K. Proteasome-mediated degradation of cotranslationally damaged proteins involves translation elongation factor 1A. Mol. Cell. Biol. 2005, 25, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Mansilla, F.; Dominguez, C.A.G.; Yeadon, J.E.; Corydon, T.J.; Burden, S.J.; Knudsen, C.R. Translation elongation factor eEF1A binds to a novel myosin binding protein-C-like protein. J. Cell. Biochem. 2008, 105, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Lamberti, A.; Sanges, C.; Chambery, A.; Migliaccio, N.; Rosso, F.; di Maro, A.; Papale, F.; Marra, M.; Parente, A.; Caraglia, M.; et al. Analysis of interaction partners for eukaryotic translation elongation factor 1A M-domain by functional proteomics. Biochimie 2011, 93, 1738–1746. [Google Scholar] [CrossRef] [PubMed]
- Mauch, F.; Mauch-Mani, B.; Boller, T. Antifungal hydrolases in pea tissue. Plant Physiol. 1988, 88, 936–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leah, R.; Tommerup, H.; Svendsen, I.; Mundy, J. Biochemical and molecular characterization of three barley seed proteins with antifungal properties. J. Biol. Chem. 1991, 266, 1564–1573. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Levy, A.; Erlanger, M.; Rosenthal, M.; Epel, B.L. A plasmodesmata-associated β-1,3-glucanase in Arabidopsis. Plant J. 2007, 49, 669–682. [Google Scholar] [CrossRef]
- Huang, Y.; Hong, H.; Xu, M.; Yan, J.; Dai, J.; Wu, J.; Feng, Z.; Zhu, M.; Zhang, Z.; Yuan, X.; et al. Developmentally regulated Arabidopsis thaliana susceptibility to Tomato spotted wilt virus infection. Mol. Plant Pathol. 2020, 21, 985–998. [Google Scholar] [CrossRef]
- Beffa, R.S.; Hofer, R.M.; Thomas, M.; Meins, F. Decreased susceptibility to viral disease of β-1,3-glucanase-deficient plants generated by antisense transformation. Plant Cell 1996, 8, 1001–1011. [Google Scholar] [CrossRef]
- Gao, C.; Zhuang, X.; Shen, J.; Jiang, L. Plant ESCRT complexes: Moving beyond endosomal sorting. Trends Plant Sci. 2017, 22, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, C.; Li, F.; Buono, R.; Roschzttardtz, H.; Chung, T.; Zhang, M.; Osteryoung, K.W.; Vierstra, R.D.; Otegui, M.S. The endosomal protein CHARGED MULTIVESICULAR BODY PROTEIN1 regulates the autophagic turnover of plastids in Arabidopsis. Plant Cell 2015, 27, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barajas, D.; Jiang, Y.; Nagy, P.D. A unique role for the host ESCRT proteins in replication of Tomato bushy stunt virus. PLOS Pathog. 2009, 5, e1000705. [Google Scholar] [CrossRef] [Green Version]
- Pastor, V.; Gamir, J.; Camañes, G.; Cerezo, M.; Sánchez-Bel, P.; Flors, V. Disruption of the ammonium transporter AMT1.1 alters basal defenses generating resistance against Pseudomonas syringae and Plectosphaerella cucumerina. Front. Plant Sci. 2014, 5, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Yoshii, A.; Sakurai, K.; Hamada, K.; Yamaji, Y.; Suzuki, M.; Namba, S.; Hibi, T. Identification of a novel tobacco DnaJ-like protein that interacts with the movement protein of Tobacco mosaic virus. Arch. Virol. 2009, 154, 959–967. [Google Scholar] [CrossRef]
- Cho, S.-Y.; Cho, W.K.; Sohn, S.-H.; Kim, K.-H. Interaction of the host protein NbDnaJ with Potato virus X minus-strand stem-loop 1 RNA and capsid protein affects viral replication and movement. Biochem. Biophys. Res. Commun. 2012, 417, 451–456. [Google Scholar] [CrossRef]
- Qiu, X.B.; Shao, Y.M.; Miao, S.; Wang, L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. CMLS 2006, 63, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Verchot, J. Cellular chaperones and folding enzymes are vital contributors to membrane bound replication and movement complexes during plant RNA virus infection. Front. Plant Sci. 2012, 3, 275. [Google Scholar] [CrossRef] [Green Version]
- Knox, C.; Luke, G.A.; Blatch, G.L.; Pesce, E.-R. Heat shock protein 40 (Hsp40) plays a key role in the virus life cycle. Virus Res. 2011, 160, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Kikkert, M.; van Lent, J.; Storms, M.; Bodegom, P.; Kormelink, R.; Goldbach, R. Tomato spotted wilt virus particle morphogenesis in plant cells. J. Virol. 1999, 73, 2288–2297. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
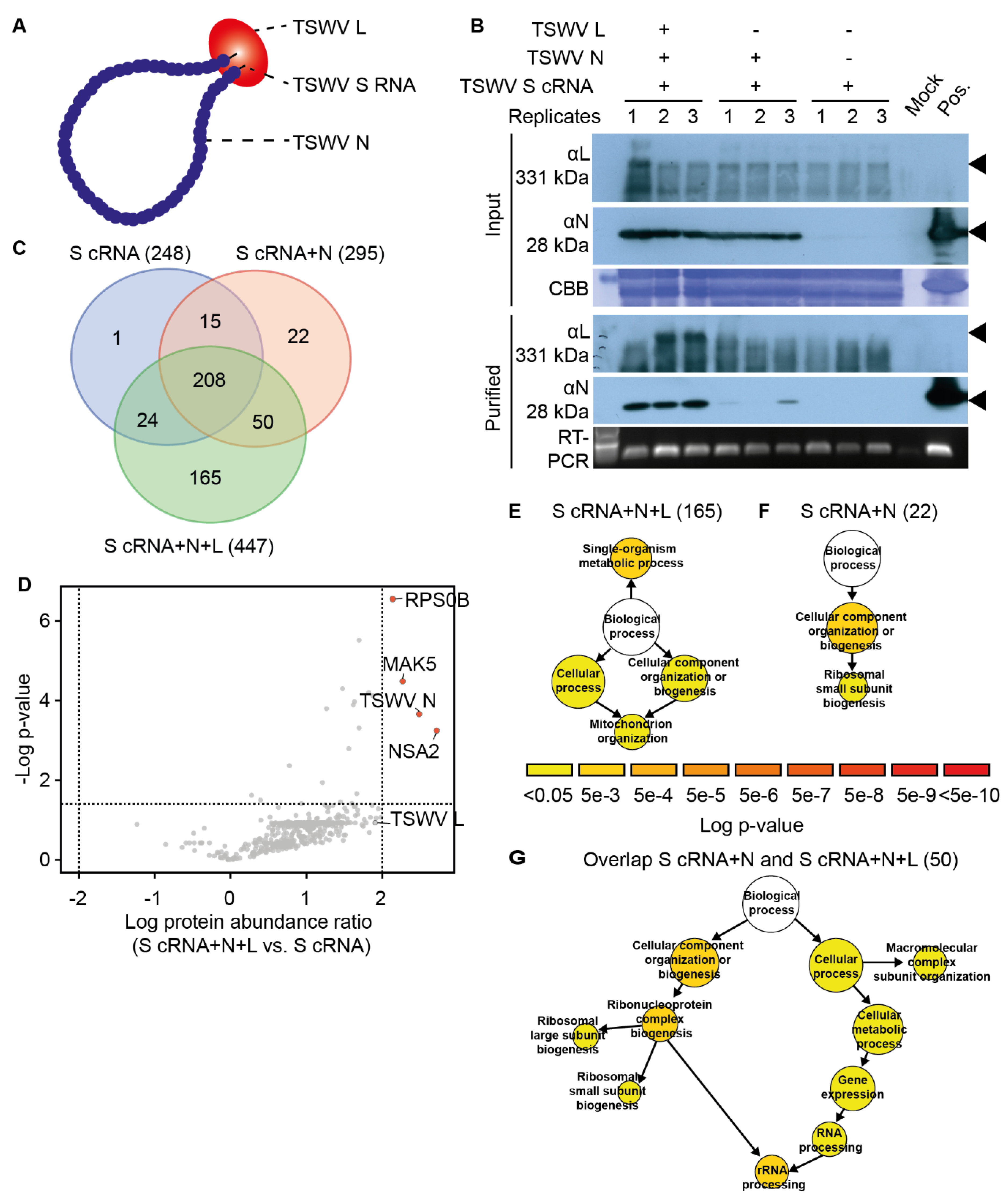{kind=link}
{kind=link}
{kind=link}
| Proteins Significant Enriched in the RNP Fraction | ||||
| Protein Annotation | Gene ID | Log Protein Abundance Ratio | −Log p-Value | GO Term Biological Processes |
| TSWV N | P25999 | 5,322364807 | 5,418489703 | Viral nucleocapsid |
| TSWV NSm | P36292 | 4,403109868 | 4,299333733 | Host–virus interaction, transport |
| Harpin inducing prot 1-like 18 | NbS00006593g0102.1 | 2,500370344 | 5,248621579 | Response to stress |
| Pleiotropic drug resistance protein 1 | NbS00010523g0001.1 | 2,279739062 | 5,176989917 | Response to stimulus, Transport |
| Probable mitochondrial chaperone BCS1-B | NbS00033465g0001.1 | 2,604620616 | 4,849324623 | Response to stress |
| Cysteine-rich receptor-like protein kinase 2 | NbS00022441g0017.1 | 2,573195457 | 4,582542858 | Cellular metabolic process |
| Cytochrome b561 | NbS00002670g0023.1 | 2,907094002 | 4,236962944 | Response to stimulus |
| Sugar transporter 13 | NbS00062251g0003.1 | 2,54924806 | 4,523826123 | Transport |
| TSWV N | Q0PTG4 | 3,811312199 | 2,677940859 | Viral nucleocapsid |
| Catalase isozyme 1 | NbS00013764g0007.1 | 2,370932102 | 3,778977343 | Response to stress |
| Pathogenesis-related protein 1 | NbS00061216g0001.1 | 2,093765418 | 3,584313513 | Defence response |
| Calcium-dependent lipid-binding (CaLB domain) family protein | NbS00018510g0001.1 | 2,740815957 | 2,934327942 | Defence response |
| Glucan endo-1,3-beta-glucosidase | NbS00010129g0001.1 | 2,055405935 | 3,606080883 | Response to stimulus, Transport |
| Inorganic phosphate transporter | NbS00006199g0011.1 | 2,083080928 | 3,522238151 | Transport |
| Hexose transporter | NbS00058697g0004.1 | 2,590578238 | 2,964279133 | Transport |
| DNA-J homolog 13 | NbS00005708g0012.1 | 2,106231213 | 3,357389199 | Protein folding |
| TSWV NSS | P26002 | 3,394036452 | 1,964366825 | Suppressor of RNA silencing |
| Clathrin light chain 2 | NbS00057576g0008.1 | 2,03093799 | 3,317251501 | Establishment of localisation |
| ABC transporter B family member 21 | NbS00020002g0005.1 | 2,291292191 | 2,847364381 | Transport |
| Embryo-specific protein ATS3B | NbS00003054g0022.1 | 2,315136909 | 2,099199888 | -- |
| Protein SSUH2 homolog isoform x2 | NbS00054171g0002.1 | 2,026340644 | 1,999542856 | Protein folding |
| TSWV L | P28976 | 2,189735095 | 1,828212439 | RNA-directed RNA polymerase |
| Inorganic phosphate transporter | NbS00023594g0004.1 | 2,184769948 | 1,807216819 | Transport |
| TSWV Gn | P36291 | 2,234362761 | 1,663396874 | Host–virus interaction |
| Proteins Significant Enriched in the Mock Fraction | ||||
| Protein Annotation | Gene ID | Log Protein Abundance Ratio | −Log p−Value | GO Term Biological Processes |
| Chloroplast photosystem I reaction centre V | NbC24101312g0001.1 | −2,362002055 | 6,199989333 | Photosynthesis |
| Photosystem I reaction centre subunit IV A | NbS00038432g0004.1 | −2,64222304 | 5,36905629 | Photosynthesis |
| Chlorophyll a-b binding protein 21 | NbS00014580g0003.1 | −2,658362707 | 4,797954045 | Photosynthesis |
| Mg protoporphyrin IX chelatase | NbS00040680g0007.1 | −2,756849289 | 4,437632643 | Chlorophyll biosynthetic process |
| Root phototropism protein 2 | NbS00008322g0011.1 | −2,00761652 | 4,429897003 | Phototropism |
| Glutamyl-tRNA reductase 1 | NbS00054987g0005.1 | −2,16445907 | 3,97925492 | Chlorophyll biosynthetic process |
| DEAD-box ATP-dependent RNA helicase 3 | NbS00021398g0012.1 | −2,179090659 | 3,935555191 | Ribosome biogenesis |
| Photosystem I reaction centre subunit IV B | NbS00019085g0006.1 | −2,744261424 | 3,31588844 | Photosynthesis |
| Photosystem I reaction centre subunit XI | NbS00028915g0014.1 | −2,117512862 | 3,870465944 | Photosynthesis |
| Suppressor of thylakoid formation 1 | NbS00012196g0002.1 | −2,017049789 | 3,25883739 | Chloroplast organisation |
| Thylakoid lumen protein 18.3 | NbS00009383g0013.1 | −2,200131734 | 2,492686209 | Chloroplast organisation |
| Chlorophyll a-b binding protein 13 | NbS00020253g0009.1 | −2,090600967 | 2,031389612 | Photosynthesis |
| Protein plastid transcriptionally active 16 | NbS00042661g0005.1 | −2,273987929 | 1,735711971 | Circadian rhythm |
| Pentatricopeptide repeat-containing protein | NbS00042677g0002.1 | −2,25110515 | 1,569628025 | Circadian rhythm |
| Oxygen-evolving enhancer protein 1 | NbS00019818g0001.1 | −2,049469153 | 1,523308383 | Photosynthesis |
| N. benthamiana ID | Yeast ID | Gene Name (Yeast) | Protein Annotation | KOG Database | Function Annotation |
|---|---|---|---|---|---|
| NbS00019858g0012.1 | P30624 | FAA1 | Long-chain-fatty-acid–CoA ligase 1 | KOG1180 | Lipid transport and metabolism |
| NbS00027624g0003.1 | P38929 | PMC1 | Calcium-transporting ATPase 2 | KOG0204 | Inorganic ion transport and metabolism |
| NbS00037522g0005.1 | Q06698 | YLR419W | Putative ATP-dependent RNA helicase | KOG0920 | Replication, recombination, and repair |
| NbS00036611g0008.1 | P32481 | GCD11 | Eukaryotic translation initiation factor 2 γ | KOG0466 | Translation, ribosomal structure, and biogenesis |
| NbS00009889g0007.1 | P35723 | YET1 | Endoplasmic reticulum transmembrane protein 1 | KOG1962 | Translation, ribosomal structure, and biogenesis |
| NbS00009856g0011.1 | P36520 | MRPL10 | 54S ribosomal protein L10 | KOG0846 | Translation, ribosomal structure, and biogenesis |
| NbS00003478g0004.1 | P46367 | ALD4 | Potassium-activated aldehyde dehydrogenase | KOG2450 | Energy production and conversion |
| NbS00048826g0001.1 | P32610 | VMA8 | V-type proton ATPase subunit D | KOG1647 | Energy production and conversion |
| NbS00039262g0004.1 | P40557 | EPS1 | ER-retained PMA1-suppressing protein 1 | KOG0191 | Energy production and conversion, post-translational modification, protein turnover, and chaperones |
| NbS00011481g0021.1 | P15108 | HSC82 | ATP-dependent molecular chaperone | KOG0019 | Post-translational modification, protein turnover, and chaperones |
| NbS00023686g0020.1 | P25694 | CDC48 | Cell division control protein 48 | KOG0730 | Post-translational modification, protein turnover, and chaperones |
| NbS00015581g0008.1 | P40358 | JEM1 | DnaJ-like chaperone | KOG0715 | Post-translational modification, protein turnover, and chaperones |
| NbS00005708g0012.1 | P25294 | SIS1 | SIt4 Suppressor1 | KOG0714 | Post-translational modification, protein turnover, and chaperones |
| NbS00006769g0021.1 | P25294 | SIS1 | SIt4 Suppressor1 | KOG0714 | Post-translational modification, protein turnover, and chaperones |
| NbS00023331g0003.1 | P10591 | SSA1 | Heat shock protein SSA1 | KOG0101 | Post-translational modification, protein turnover, and chaperones |
| NbS00040361g0003.1 | P10591 | SSA1 | Heat shock protein SSA1 | KOG0101 | Post-translational modification, protein turnover, and chaperones |
| NbS00004424g0012.1 | Q04602 | VBA4 | Vacuolar basic amino acid transporter 4 | KOG0254 | Intracellular trafficking, secretion, and vesicular transport |
| NbS00007660g0008.1 | Q04602 | VBA4 | Vacuolar basic amino acid transporter 4 | KOG0254 | Intracellular trafficking, secretion, and vesicular transport |
| NbS00023311g0013.1 | Q04602 | VBA4 | Vacuolar basic amino acid transporter 4 | KOG0254 | Intracellular trafficking, secretion, and vesicular transport |
| NbS00058697g0004.1 | Q04602 | VBA4 | Vacuolar basic amino acid transporter 4 | KOG0254 | Intracellular trafficking, secretion, and vesicular transport |
| NbS00062251g0003.1 | Q04602 | VBA4 | Vacuolar basic amino acid transporter 4 | KOG0254 | Intracellular trafficking, secretion, and vesicular transport |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helderman, T.A.; Deurhof, L.; Bertran, A.; Boeren, S.; Fokkens, L.; Kormelink, R.; Joosten, M.H.A.J.; Prins, M.; van den Burg, H.A. An Isoform of the Eukaryotic Translation Elongation Factor 1A (eEF1a) Acts as a Pro-Viral Factor Required for Tomato Spotted Wilt Virus Disease in Nicotiana benthamiana. Viruses 2021, 13, 2190. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112190
Helderman TA, Deurhof L, Bertran A, Boeren S, Fokkens L, Kormelink R, Joosten MHAJ, Prins M, van den Burg HA. An Isoform of the Eukaryotic Translation Elongation Factor 1A (eEF1a) Acts as a Pro-Viral Factor Required for Tomato Spotted Wilt Virus Disease in Nicotiana benthamiana. Viruses. 2021; 13(11):2190. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112190
Chicago/Turabian StyleHelderman, Tieme A., Laurens Deurhof, André Bertran, Sjef Boeren, Like Fokkens, Richard Kormelink, Matthieu H. A. J. Joosten, Marcel Prins, and Harrold A. van den Burg. 2021. "An Isoform of the Eukaryotic Translation Elongation Factor 1A (eEF1a) Acts as a Pro-Viral Factor Required for Tomato Spotted Wilt Virus Disease in Nicotiana benthamiana" Viruses 13, no. 11: 2190. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112190