1. Introduction
During its replication cycle, HIV-1 forms two distinct particles, immature and mature ones. An immature HIV-1 particle is formed by Gag polyprotein precursors consisting of four structural domains—matrix (MA, p17), capsid (CA, p24), nucleocapsid (NC), and p6—and two short spacer peptide sequences linking CA with NC (SP1) and NC with p6 (SP2). The assembly of the immature HIV-1 particle is driven mainly by interactions among CA domains of Gag polyprotein triggered by the NC–viral genomic RNA scaffold. The NC domain specifically recognizes, dimerizes, and packages viral genomic RNA into a nascent immature particle. CA-mediated multimerization on the inner leaflet of the plasma membrane (PM) leads to the formation of the hexameric lattice of the immature particle that buds into the extracellular environment. The transition to a mature, fully infectious HIV-1 particle is triggered by activation of the virus-encoded protease (PR), which cleaves Gag polyproteins into individual viral proteins. CA proteins released from the Gag precursor re-assemble to form a mature viral core. The mature HIV-1 hexameric lattice, formed approximately by 250 CA hexamers and 12 pentamers [
1,
2,
3,
4], encloses and protects viral genomic RNA, complexed to NC. The insertion of CA pentamers allows the formation of a closed conical core, characteristic of HIV-1 [
2,
5].
To initiate the replication cycle, HIV-1 must deliver its genome into the host cell. After HIV-1 enters the cell, the protective mature core is removed in a stepwise process called uncoating. This disassembly process, which is not well understood [
6,
7,
8,
9,
10,
11,
12,
13,
14], is connected to both reverse transcription and transport toward the nucleus, where viral DNA integration into the cellular genome takes place [
12,
15,
16]. The nucleic acid chaperone activity of NC facilitates the process of reverse transcription, during which viral genomic RNA is reversely transcribed into double-stranded DNA. The key structural and functional roles of CA and NC make them attractive pharmacological targets [
17,
18,
19,
20,
21].
After diamond and graphite, fullerenes were discovered to be the third family of carbon allotropes [
22]. They are highly symmetric cages of different sizes (e.g., C
60) with unique structures and physical properties. To improve their lack of solubility, several fullerene derivatives containing hydrophilic groups have been reported [
23,
24]. These derivatives have useful medicinal applications, as they possess a broad spectrum of biological activities, such as antiviral, antimicrobial, antiproliferative, antioxidant, neuroprotective, and anti-cancer [
24,
25,
26,
27]. Concerning their anti-viral activities, proline-based fullerene derivatives inhibit the activity of NS3/4A protease and NS5B polymerase of the flaviviral hepatitis C virus (HCV) [
23,
28,
29]. Giant glycofullerenes were reported to function as potent inhibitors of a reporter of Ebola virus [
30,
31]. Several cationic, anionic, and zwitterion fullerene derivatives of proline type, pyrrolidinium type, pyridine/pyridinium type, and others, were shown to block the activity of HIV-1 reverse transcriptase [
23,
28,
32]. A water-soluble, (bis-(phenethylamino-succinate) C
60 fullerene derivative was the first one reported to block the activity of HIV-1 protease
in vitro. The mechanism of inhibition—i.e., fullerene binding to its ellipsoid-shaped, hydrophobic active site—was postulated based on in silico predictions [
33,
34]. Recent studies confirmed that C
60 and C
70 fullerene derivatives of pyrrolidinium type indeed strongly inhibit HIV-1 replication—without impairing viral protease activity, however [
35,
36]. Using immunoprecipitation experiments with magnetic beads functionalized with
N,
N-dimethyl[70]fulleropyrrolidinium iodide showed that the interactions of these C
70 fullerene derivatives occur with immature HIV-1 CA-SP1 protein [
36]. Based on this result, a model was proposed in which binding of C
70 fullerene derivatives to the CA domain of Gag polyprotein modifies either the conformation or the assembly of the HIV-1 polyproteins, thereby altering the specificity or activity of HIV-1 PR. This model led to the conclusion that C
70 fullerene derivatives act as the inhibitors of maturation [
36].
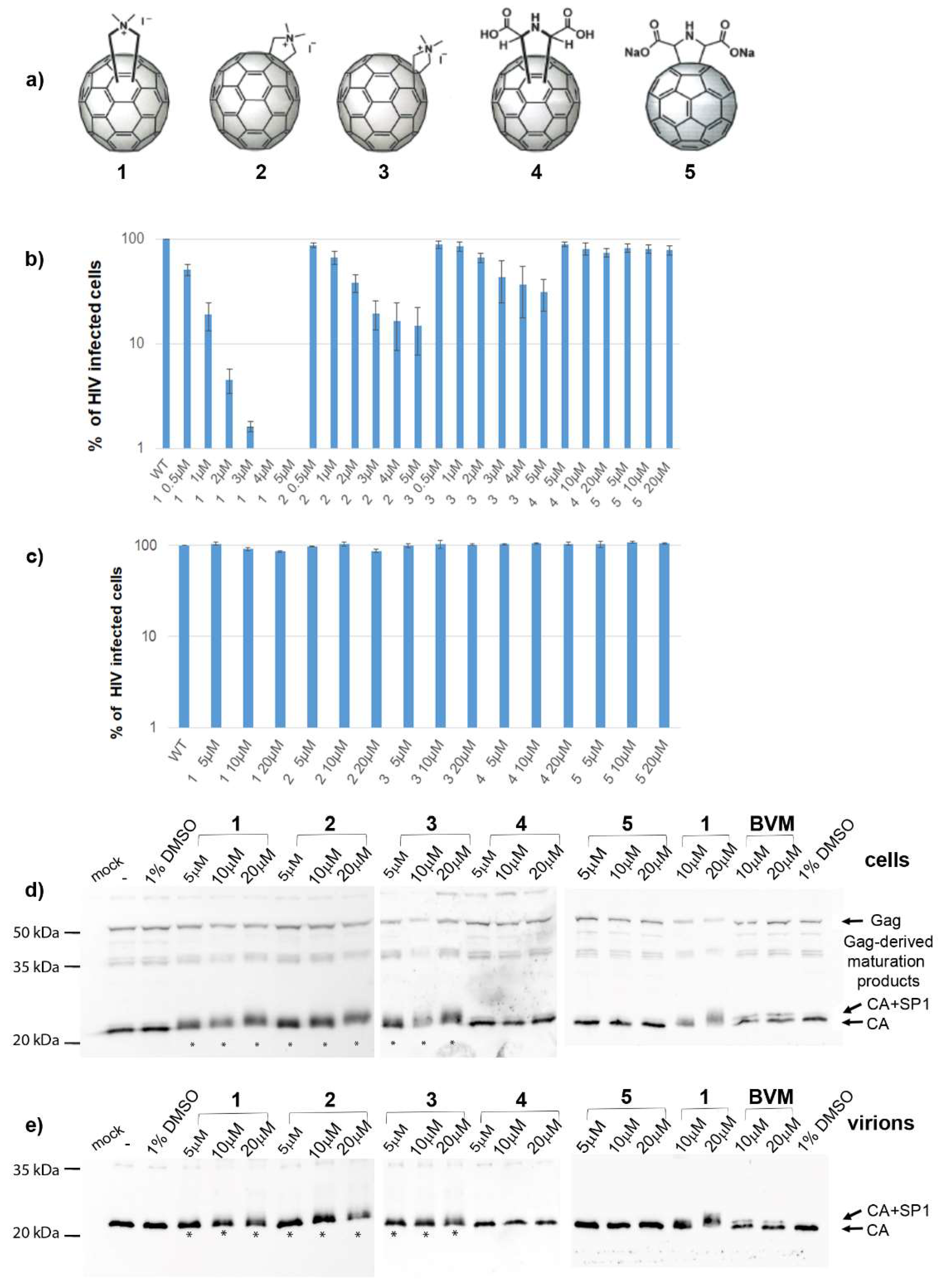
We tried to unveil the mechanism of action of
N,
N-dimethyl[70]fulleropyrrolidinium iodide fullerene derivatives (
Figure 1a). In accordance with the recently published data [
36], we confirmed that these fullerene derivatives had no effect on HIV-1 protease activity in either
in vitro or cell-based experiments. Moreover, microscale thermophoresis (MST) analysis showed a low binding affinity of fullerene
1 to HIV-1 protease. Additionally, our data showed that HIV-1 Gag maturation was not affected by fullerene
1. No effect of fullerene
1 was observed in HIV-1 RT activity either. However, in agreement with Martinez et al. [
35], we observed strong inhibition of all steps of HIV-1 cDNA synthesis during reverse transcription. We correlated this observation with the finding that virions released from the cells treated with fullerene
1 contained less viral genomic RNA (gRNA); the amount of gRNA was inversely proportional to the inhibitor concentration. MST and the electrophoretic mobility shift assay (EMSA) revealed a strong binding affinity of fullerene
1 to HIV-1 NC and nucleic acids.
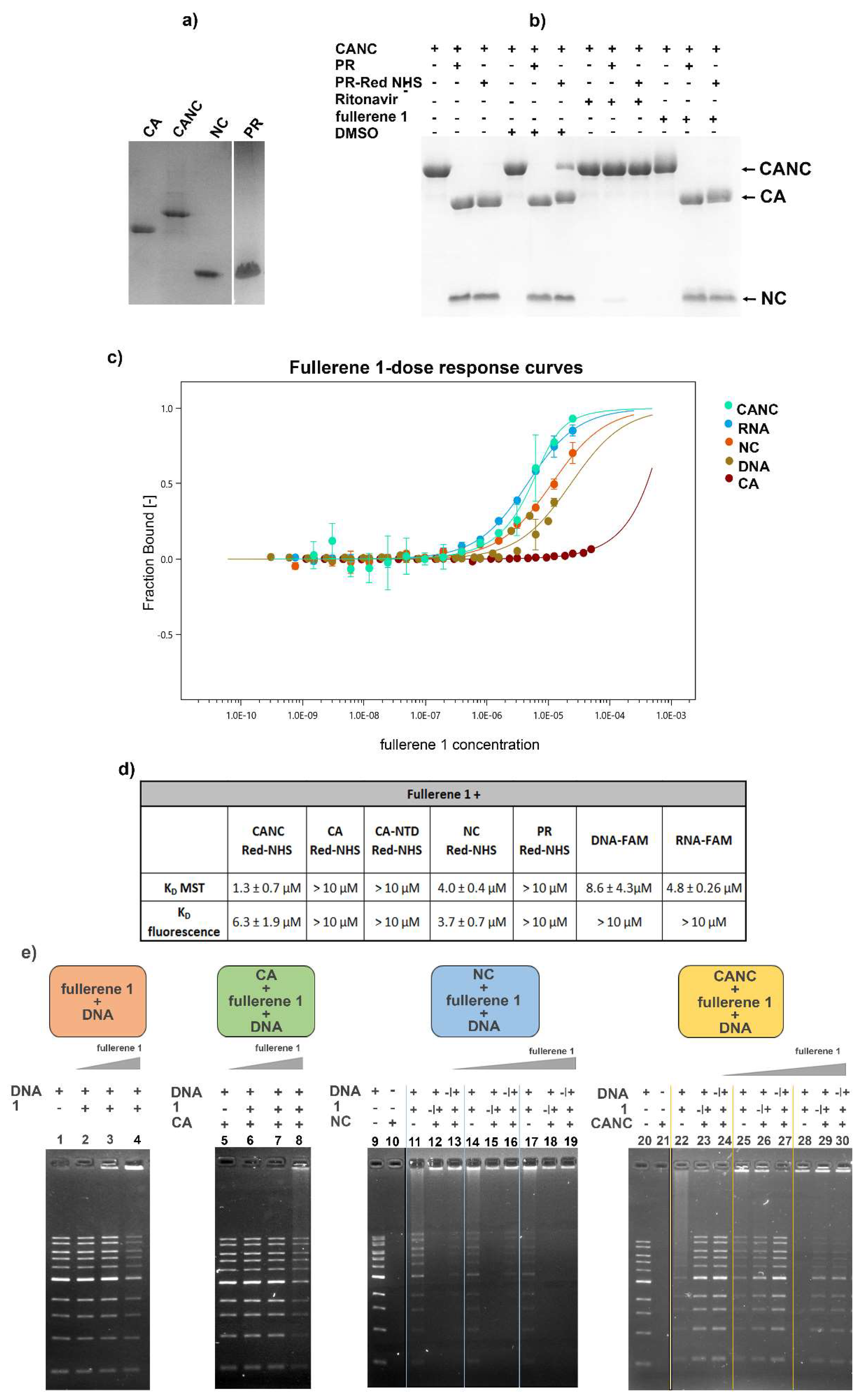
In conclusion, our data suggest that fullerene 1, due to strong binding affinity for the HIV-1 nucleocapsid domain, prevents the proper interaction of viral genomic RNA with the Gag polyprotein, thereby blocking HIV-1 infectivity. We also found out fullerene 1 possesses both oxidative and fluorescence quenching activities, strongly interfering with some analysis (e.g., ELISA). We think that this feature could be one of the reasons for inconsistency among reported anti-HIV-1 mechanisms.
2. Materials and Methods
2.1. Synthesis of Fullerenes 1–5
Fullerenes
1–
3 were synthesized by following previously reported methodologies [
36,
37]. Briefly, a mixture of C
70 (84 mg, 0.1 mmol), sarcosine (22 mg, 0.24 mmol), and paraformaldehyde (18 mg, 0.60 mmol) was refluxed in toluene (75 mL) for 1 h. The resulting brown solution was concentrated in vacuo, and the crude mixture was purified by silica-gel chromatography using toluene as eluent. Pure C
60- and C
70-fullerene derivatives were obtained by HPLC using a Buckyprep column and toluene as eluent. Fullerenes
1–
3 were synthesized from their corresponding neutral starting materials, which were dissolved in methyl iodide and stirred for 48 h at room temperature. The black precipitate was filtered and washed with CS
2, CHCl
3, toluene, MeOH, and acetone, in that order. The precursors of 4 and 5 were synthesized by following our previously reported methodology [
38]. C
60 or C
70 (0.10 mmol) was dissolved in chlorobenzene (20 mL) under sonication for 5 min; then DIB (200 mg, 0.62 mmol), glycine methyl ester hydrochloride (100 mg, 0.79 mmol) and sodium carbonate decahydrate (100 mg, 0.35 mmol) were added. The flask was wrapped with aluminum foil and sonicated for 30 min. The pure C
60 mono-adduct or the C
70 mono-regioisomeric mixture was obtained after a silica gel column purification using a toluene/ethyl acetate (9:1) mixture. The corresponding dicarboxylic methyl ester C
60/C
70 derivative (0.015 mmol) was dissolved in toluene (20 mL) and NaH (120.00 mg, ~0.120 mmol); then, 50–60% in oil was slowly added and the reaction was stirred at room temperature for 1 h. Then, ethanol (3 mL) was added followed by the addition of HCl (3 M, 2 mL). Then, the reaction was stirred at room temperature overnight. The solvent was evaporated under reduced pressure; the precipitate was washed with chloroform, methanol, and cold water; and finally, compound
4/
5 was recrystallized from methanol/acetone.
2.2. VSV-G Pseudotyped HIV-1 Particles Production
HIV-1 particles were obtained from HEK 293 cells, cotransfected by a combination of three vectors: packaging psPAX2 vector encoding HIV Gag, Pol, Tat, and Rev; reporter/transfer pWPXLd-GFP vector encoding LTR, RRE, and GFP as a reporter; and envelope pHEF-VSV-G vector, encoding vesicular stomatitis virus Env, VSV-G. The psPAX2 vector was kindly provided by Dr. Jeremy Luban; the pWPXLd-GFP and pHEF-VSV-G vectors were purchased from Addgene. HEK-293 cells were grown in Dulbecco’s modified Eagle medium (DMEM, Sigma, St. Louis, MI, USA) supplemented with 10% fetal bovine serum (Sigma, St. Louis, MI, USA) and 1% l-glutamine (Sigma, St. Louis, MI, USA) at 37 °C under 5% CO2. A day before transfection, cells were plated at 3 × 105 cells per well. The following day, cells were transfected with the appropriate vectors using polyethylenimine (PEI, 1 mg/mL) at a 2:1 PEI:DNA ratio. Four hours post transfection, the culture medium was replaced with fresh DMEM, containing various concentrations of tested compounds, solubilized in DMSO. At 48 h post-transfection, the culture media containing released virions were harvested, filtered through 0.45-µm pores membrane, and used for immunochemical quantification and characterization by Western blot using rabbit anti-HIV-1 CA antibody.
2.3. Single-Round Infectivity Assay HIV-1
The infectivity was determined similarly as described earlier [
39]. Briefly, 48 h post-transfection, the culture media from HEK 293 cells transfected with psPAX2, pWPXLd-GFP, and pHEF-VSV-G vectors at a ratio 1:1:1, in the presence of tested compounds, were collected and filtered through a 0.45 µm membrane filter. HIV-1 CA content was determined by ELISA. The freshly seeded HEK 293 cells were infected with WB-normalized amounts of VSV-G pseudotyped HIV-1 particles and incubated for 48 h. The cells were fixed with 2% paraformaldehyde and transferred to a FACS tube. Quantification of GFP-positive cells was performed using a BD FACS Aria III flow cytometer.
2.4. Flow Cytometry
The VSV-G-GFP-HIV infected cells were analyzed with a BD FACS Aria III flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA) with excitation at 488 nm and emission separated by a 530/30 band pass filter, as described earlier [
39]. The obtained data were analyzed with FACSDiva software, version 8.0.
2.5. Transmission Electron Microscopy Analysis
HEK 293 cells producing VSV-G pseudotyped HIV-1 particles were washed with PBS 48 h post-transfection and fixed with freshly prepared 3% glutaraldehyde in 0.1 M cacodylate buffer, pH 7.5. The cells were then postfixed in 1% osmium tetroxide, dehydrated in an ethanol concentration series (30%, 50%, 70%, 80%, 90%, and 100%), and embedded in Agar 100 epoxy resin. Ultrathin sections (70 nm) prepared using an RMC MT 7000 Ultramicrotome were placed onto 200-mesh copper grids and contrasted with saturated uranyl acetate and lead citrate. The samples were then analyzed with a transmission electron microscope (JEOL; JEM-1200E×, Tokio, Japan) operating at 60 kV.
2.6. Quantification of VLP Assembly Efficiency by FAITH
HIV-1 CANC protein (18 µM) or ΔMACANCSP2 (15 µM) in 96-well plate was pre-incubated with tested inhibitors in the molar ratio 1:1 and kept on ice for 1 h. To start assembly reaction dually labelled oligonucleotide (tqON) was added to proteins in the molar ration protein:tqON 10:1, and the volume of the reaction mixture was adjusted to 100 µL by assembly buffer (50 mM Tris, pH 8.0, 1 µM ZnCl2, 340 mM NaCl). Following 3 h incubation at room temperature, Exonuclease I (ExoI) with Mg2+ ions was added, and the fluorescence of the fluorophore released from degraded tqON was measured using a Tecan M200Pro plate reader. The efficiency of assembly is calculated based on the formula E = 100 × ∆F2/∆F1, in which ∆F1 corresponds to the difference between relative fluorescence of tqON and relative fluorescence of tqON in the presence of HIV-1 Gag-derived protein, and ∆F2 corresponds to the difference between relative fluorescence of tqON and relative fluorescence of tqON in the presence of HIV-1 Gag-derived protein and inhibitors.
2.7. Western Blot and Semi-Quantitative Western Blot
At 48 h post-transfection, 100 µL aliquots of virus-containing culture media were combined with 20 µL of PLB (6×), and the samples were analyzed by Western blot using rabbit anti-HIV-1 CA (p24) (in house production). Proteins were resolved by reducing SDS-PAGE and blotted onto a nitrocellulose membrane. The antigen-antibody complexes were detected by Clarity™ Western ECL Substrate (Bio-Rad) and visualized using FUSION 7S system (Vilber Lourmat, Marne-la-Vallée, France). As CA(p24)-ELISA based normalization of HIV-1 viral particles could not be used due to interference of tested fullerene 1 with TMB reaction, semi-quantitative Western blot was used as described earlier [
40]. To quantify and normalize the amount of produced HIV-1 particles, the standard curve was prepared from recombinantly expressed and purified HIV-1 CA(p24). Concentrations of CA(p24) protein from 1 to 2000 ng were separated on 15% SDS-PAGE gels and transferred onto a nitrocellulose membrane. Then, the same Western blot procedure described above was performed. The Western blot signals of CA(p24) in the calibration curve, and the viral samples, were determined using Quantum gel documentation imaging system (Vilbert Lourmat), and the amount of CA(p24) was calculated.
2.8. Pulse-Chase Experiment
This experiment was performed as described earlier [
41,
42,
43] with some modifications. Forty-eight hours after transfection with appropriate DNA vectors in the presence of increasing amounts of fullerene 1, HEK 293 cells were starved for 30 min in methionine- and cysteine-deficient DMEM, pulse-labeled for 30 min with 125 Ci/mL Tran35S-label (MGP, Zlín, Czech Republic), and chased overnight in complete DMEM. The cells from pulse and pulse-chase experiments were lysed in lysis buffer (25 mM Tris-HCl pH 8.0 containing 50 mM NaCl, 1% Triton X-100, and 1% sodium deoxycholate) and centrifuged at 14,000×
g for 2 min. The culture medium from the chased cells was filtered through a 0.45 µm membrane filter, and SDS was added to a final concentration of 0.1%. Viral proteins from both the cellular lysates and culture medium were immunoprecipitated by using an anti-HIV-1 CA antibody, followed by incubation with immobilized protein A (Invitrogen, Waltham, MA, USA). Radiolabeled proteins were separated by SDS-PAGE and detected by using a Typhoon PhosphorImager (Cytiva, Marlborough, MA, USA).
2.9. qPCR Analysis of Reverse Transcription Proceeding
Three different products of reverse transcription were analyzed using qPCR experiments performed according to previously reported protocol [
44]. VSV-G pseudotyped HIV-1 particles produced in the absence or presence of 5 µM fullerene 1 were used to infect fresh HEK 293 cells. At various times post-infection (2, 4, 6, 10, 24, 33, and 48 h), the cells were collected, pelleted, washed with PBS, and used for the isolation of total DNA with the DNeasy blood and tissue kit (Qiagen, Hilden, Germany), according to the manufacturer’s protocol. To determine the level of different reverse transcription products, 2 µL of the isolated total DNA was mixed with qPCR 2× SYTO-9 master mix (Top-Bio, Vestec, Czech Republic), ROX as the reference dye at final concentration 25 nM, and 1 µM primers: ssDNA-F (5′-TAA CTA GGG AAC CCA CTG C-3′) and ssDNA-R (5′-GCT AGA GAT TTT CCA CAC TG-3′); EGFP-F (5′- GCG CAC CAT CTT CTT CAA GG-3′) and EGFP-R (5′-GTG TCG CCC TCG AAC TTC AC-3′); 2-LTR-F (5′-TAA CTA GGG AAC CCA CTG C-3′) and 2-LTR-R (5′-CTG CGT CGA GAG AGC TCC TCT GGT T-3′). To establish a calibration curve, appropriate DNA complementary to pairs of the primers was used in the concentration range of 0.06 to 250 pg/µL. To normalize the amounts of isolated viral DNAs among the individual samples, two endogenous control genes, encoding the phospholipase A (PLA) and C-C chemokine receptor type 5 CCR5, were selected: PLA-F (5′-AAG TTC TTG ATC CCC AAT GCT T-3′) and PLA-R (5′-GTC TGA TAG GAT GTG TTG GTT GC-3′); CCR5-F (5′-CCA GAA GAG CTG AGA CAT CCG-3′) and CCR5-R (5′-GCC AAG CAG CTG AGA GGT TAC T-3′)). qPCRs were performed in 96-well plates by using a QuantStudio 5 real-time PCR system (Applied Biosystems, Waltham, MA, USA) under the following reaction conditions: 2 min at 95 °C, followed by 45 cycles of 1 min at 95 °C, 30 s at 60 °C, and 30 s at 72 °C.
2.10. qPCR Analysis of Incorporated gRNA
gRNA incorporated into the HIV-1 particles was isolated and quantified as previously described [
45]. Briefly, 48 h after transfection, the medium containing HIV-1 particles was harvested, filtered through 0.45 µm pores and centrifuged through 20% sucrose cushion at 210,000×
g for 90 min in the SW41Ti rotor (Beckman Coulter, Brea, CA, USA). The pellet was resuspended in 160 µL of PBS: 140 µL was used to isolate gRNA, and the rest was analyzed using Western blot. Total RNA was isolated by using QIAamp viral RNA minikit (Qiagen, Hilden, Germany). The isolated viral RNA was incubated with Turbo DNase for 30 min at 37 °C, and the reaction was stopped by heating the mixture at 70 °C for 10 min. To control the following processes: reverse transcription and qPCR, TATAA universal RNA spike I (TATAA Biocenter, Gothenburg, Sweden) was added to each sample before reverse transcription (RT). Before RT, the amount of gRNA was normalized to the CA (p24) amount using a semi-quantitative Western blot. gRNA was transcribed to cDNA using FeLV reverse transcriptase (prepared in house). Subsequently, cDNA was used as a template in qPCR reaction prepared by mixing of 2× SYTO-9 master mix and EGFP-specific primers: EGFP-F (5′-GCG CAC CAT CTT CTT CAA GG-3′) and EGFP-R (5′-GTG TCG CCC TCG AAC TTC AC-3′). Specific primers, designed by TATAA Biocenter, were used to quantify the control RNA spikes. qPCR was performed by using a QuantStudio 5 real-time PCR system (Applied Biosystems, Waltham, MA, USA).
2.11. Cyclosporine Washout Assay
The cyclosporine A (CsA) washout assay in Owl monkey kidney (OMK) cells was performed as described earlier [
39,
46]. VSV-G pseudotyped particles were produced in HEK 293 cells in the presence or absence of CsA and with various amounts of fullerene 1. OMK cells were seeded in a 48-well plate in Eagle’s minimum essential medium (EMEM). The next day, the OMK cells were spinoculated with a normalized amount of the HIV-1 particles in EMEM containing 2.5 µM CsA and polybrene (5 ng/µL). Optionally, PF74 (5 µM) was added to OMK cells concomitantly with the infection. At indicated times, the cultivation medium containing CsA was replaced with fresh medium without CsA. After 48 h, the cells were fixed with 2% formaldehyde and GFP-positive cells were counted using flow cytometry.
2.12. The Activity of Reverse Transcriptase
The medium containing VSV-G pseudotyped HIV-1 particles produced in the absence or presence of fullerene 1 was harvested and filtered through 0.45 µm pores. The activity of reverse transcriptase in individual samples was then determined using the reverse transcriptase Colorimetric Assay (Sigma, St. Louis, MI, USA) according to the manufacturer’s protocol, as described earlier. The absorbance of the colorimetric reaction was measured using an Infinite
® 200 PRO series spectrophotometer (Tecan M200, TECAN, Männedorf, Switzerland) at 490 nm [
45].
2.13. Thermophoresis
The HIV-1 proteins were fluorescently labeled using Protein Labeling Kit Red-NHS (Nanotemper Technologies, München, Germany). The labeling reaction was performed according to the manufacturer’s instructions. The 20 µM protein was mixed with the dye (1:3 molar) and reacted for 30 min. The unreacted dye was removed with supplied dye removal column. The labeled proteins and tubules were diluted 20×, and 5% DMSO was added. The labeled RNA and DNA oligomers were diluted to a concentration 20 nM. Fullerene 1 was dissolved in the same buffer and a series of 16 1:1 dilutions. For the measurement, each ligand dilution was mixed with one volume of labeled molecules, which led to a final concentration of fullerene ranging from 50 µM to 15 nM. The samples were loaded into Monolith NT.115 Capillaries (NanoTemper Technologies, München, Germany). MST was measured using a Monolith NT.115 instrument (NanoTemper Technologies, München, Germany) at temperature of 25 °C. Instrument parameters were adjusted to 100% LED power and medium MST power. Data of three independent measurements were analyzed using Affinity Analysis software version 2.3 (NanoTemper Technologies, München, Germany) using the signal from an MST over 1.5 s and the initial fluorescence data.
2.14. Electrophoretic Mobility Shift Assay (EMSA)
The binding affinity of
1 to HIV-1 CA-NC was verified and visualized using Electrophoretic Mobility Shift Assay (EMSA) according to an established protocol [
14,
47]. Reaction mixtures in the buffer containing 20 mM Tris-HCl, pH 7.5, 100 mM NaCl were prepared to keep the final concentrations of nucleic acid (1 kB DNA Ladder, NEB, Ipswich, MA, USA) 200 ng, and those of HIV-1 CA, NC, and CANC protein 1 μM. The final concentrations of
1 in the reactions were 1, 2, and 5 μM, corresponding to the final molar ratios of protein:fullerene
1 of 1:1, 1:2, and 1:5. To assess the binding affinity of
1 to nucleic acid, DNA was incubated with various amounts of
1 for 40 min at RT. When interactions among protein–fullerene
1–nucleic acid were analyzed, two experimental arrangements were tested. In the first one, HIV-1 NC or CANC proteins were incubated for 20 min at RT with an excessive amount of DNA, along with fullerene
1 in different amounts
. After incubation for 20 min at RT, the samples were resolved by 0.8% agarose gel electrophoresis. In the second arrangement, NC or CANC proteins were first preincubated for 20 min at RT with fullerene 1, and then DNA was added. Incubation proceeded for another 20 min at RT. To ensure that proteins were not contaminated with nucleic acids, the protein samples were incubated in the presence of DMSO (1%) and then loaded on the agarose gel. All samples were analyzed using 0.8% agarose gel electrophoresis in TAE (1×) buffer at 110 V, stained with Gel Red, and visualized with a Quantum gel documentation imaging system (Vilbert Lourmat, Collégien, France).
2.15. Docking of N,N-Dimethyl[70] Fulleropyrrolidinium Iodide
The structures of capsid (3NTE) and nucleocapsid proteins (NC) were obtained from the Brookhaven database (1F6U) by removing the stem-loop SL2 of the ψ-RNA packaging signal. The ligand structure was built from the C
70 fullerene, downloaded from the CHEBI server (
https://www.ebi.ac.uk/chebi/searchId.do?chebiId=CHEBI%3A33195, accessed on: 14 March 2021). Docking of fullerene 1 was carried out in two programs, i.e., PLANTS [
48] and AutoDock Vina, version 1.5.6rc3 [
49]. Both the ligand and protein were prepared correctly in auxiliary software delivered with the docking programs.
4. Discussion
In this work, we studied the mechanism of the anti-HIV-1 activity of N,N-dimethyl[70]fulleropyrrolidinium iodide fullerene derivative 1 by investigating various steps of the HIV-1 replication cycle: assembly, release, processing and maturation, viral genomic RNA incorporation, uncoating, and reverse transcription. We found that 1 affected the amount of viral genomic RNA in HIV-1 viral particles in a concentration-dependent manner. Logically, we also studied the impact of 1 on the processes associated with reverse transcription; however, neither reverse transcriptase nor protease was inhibited by these C70 derivatives. Using MST, we observed a strong binding affinity of fullerene 1 for HIV-1 NC. This binding was observed for both forms of NC, the Gag-mimicking immature domain (within the CA-NC precursor) and the mature NC protein. Fullerene 1 also bound nucleic acids; however, its affinity for the immature form of NC (CA-NC fusion protein) was about three times higher than that to RNA oligonucleotide. In contrast, the affinities of 1 for CA protein and HIV-1 protease were at least one order of magnitude lower. Altogether, our results suggest that fullerene 1 binds to HIV-1 NC and thus affects its ability to specifically interact with viral genomic RNA, thereby blocking HIV-1 infectivity.
Previously it was suggested that fullerene derivatives affect HIV-1 maturation [
35,
36]. One of the reasons why
1 was suggested to act as a maturation inhibitor was the presence of partially processed CA-containing Gag fragments such as CA-SP1-NC and CA-SP1 in fullerene-treated HEK293 cells producing HIV-1 Gag [
35,
36]. However, a more detailed analysis performed here via both
in vitro and cell-based HIV-1 Gag polyprotein processing analysis with a C
70 fullerene-derivative excluded this mechanism of action. We did not observe any impact on HIV-1 protease activity or Gag polypeptide processing under
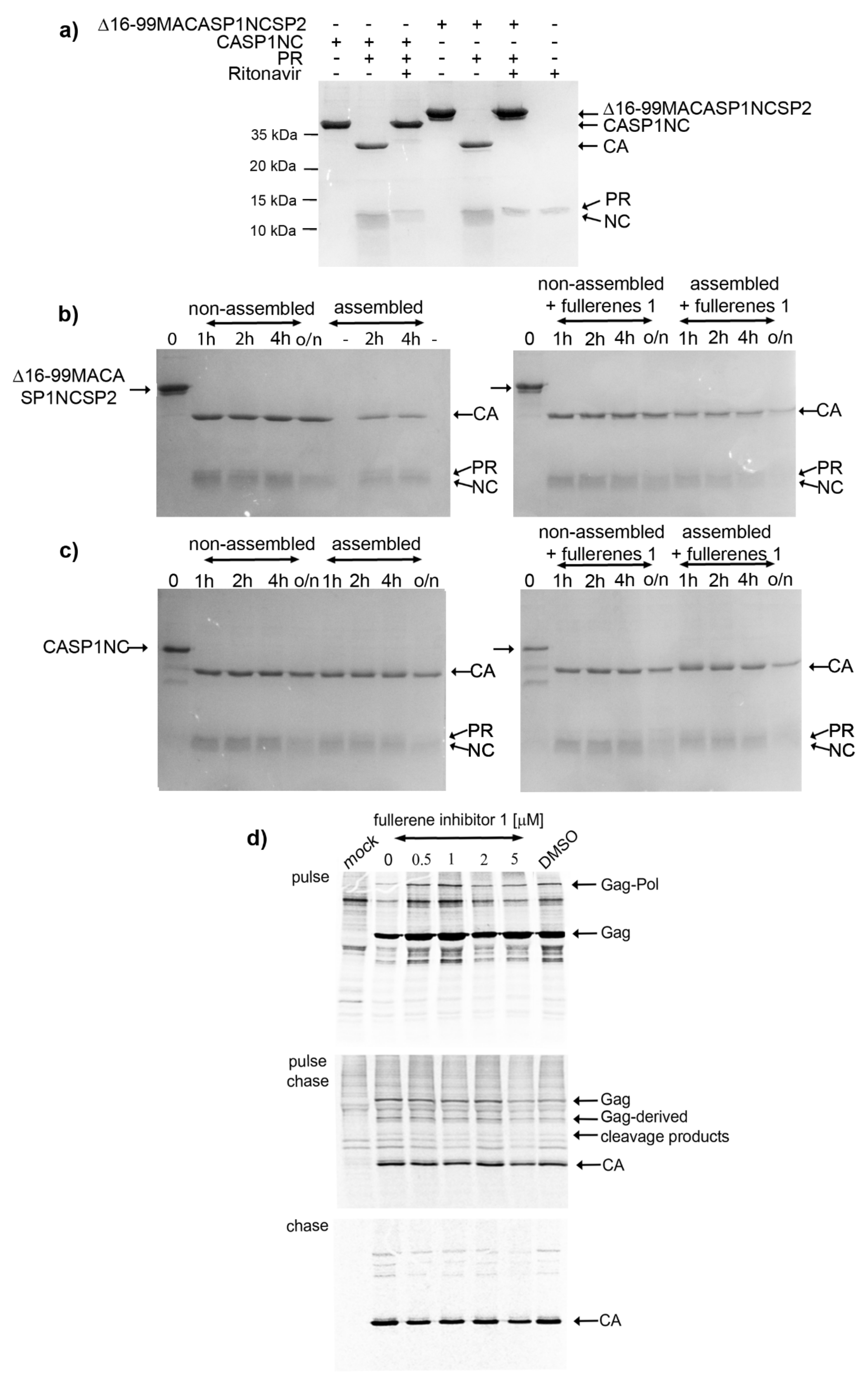
in vitro conditions, where recombinant HIV-1 protease was added to Gag-derived, purified polypeptides Δ16-99MACASP1NCSP2 and CASP1NC. There was a possibility that the fullerene-dependent effect on HIV-1 processing could be connected to conformational changes of Gag polyprotein occurring during assembly. However, we found no impact of
1 on the processing of
in vitro assembled spherical, immature-like Δ16-99MACASP1NCSP2 or tubular, mature-like CASP1NC particles. To verify this
in vitro observation, we analyzed the effect of
1 on the processing and maturation of HIV-1 Gag and Gag–Pol polyproteins authentically produced in HEK 293 cells. Similarly, viruses released from fullerene-derivatives treated HEK 293 cells showed no defects in Gag processing. At all tested concentrations (0.5–5 µM) of
1, we observed the HIV-1 polyprotein precursors (i.e., Gag Pr55 and Gag–Pol 160) and mature CAp24 of expected sizes and in ratios comparable to those of the fullerene non-treated HIV-1 producing cells.
The analysis of the cells from the chase experiment showed both fully processed CA and partially processed CA-containing Gag fragments. However, both these forms were present also in the non-treated or DMSO-only treated control samples. We assume that these CA-containing Gag fragments are generated during the initial phase of maturation when the budding viral particles still remain attached to the membrane. This assumption is supported by the pulse-chase experiment where the viruses released in the culture media contained fully processed CAp24 at all fullerene concentrations tested. The fuzzy-like appearance of the CAp24 band, which we also observed by Western blot analysis (
Figure 1d,e), was, however, not observed in the pulse-chase experiments. This may have been due to different sample treatment in the pulse-chase (i.e., immunoprecipitation) compared to the Western blot. Nevertheless, based on the above discussed results, we conclude that
1 did not affect HIV-1 Gag polyprotein maturation.
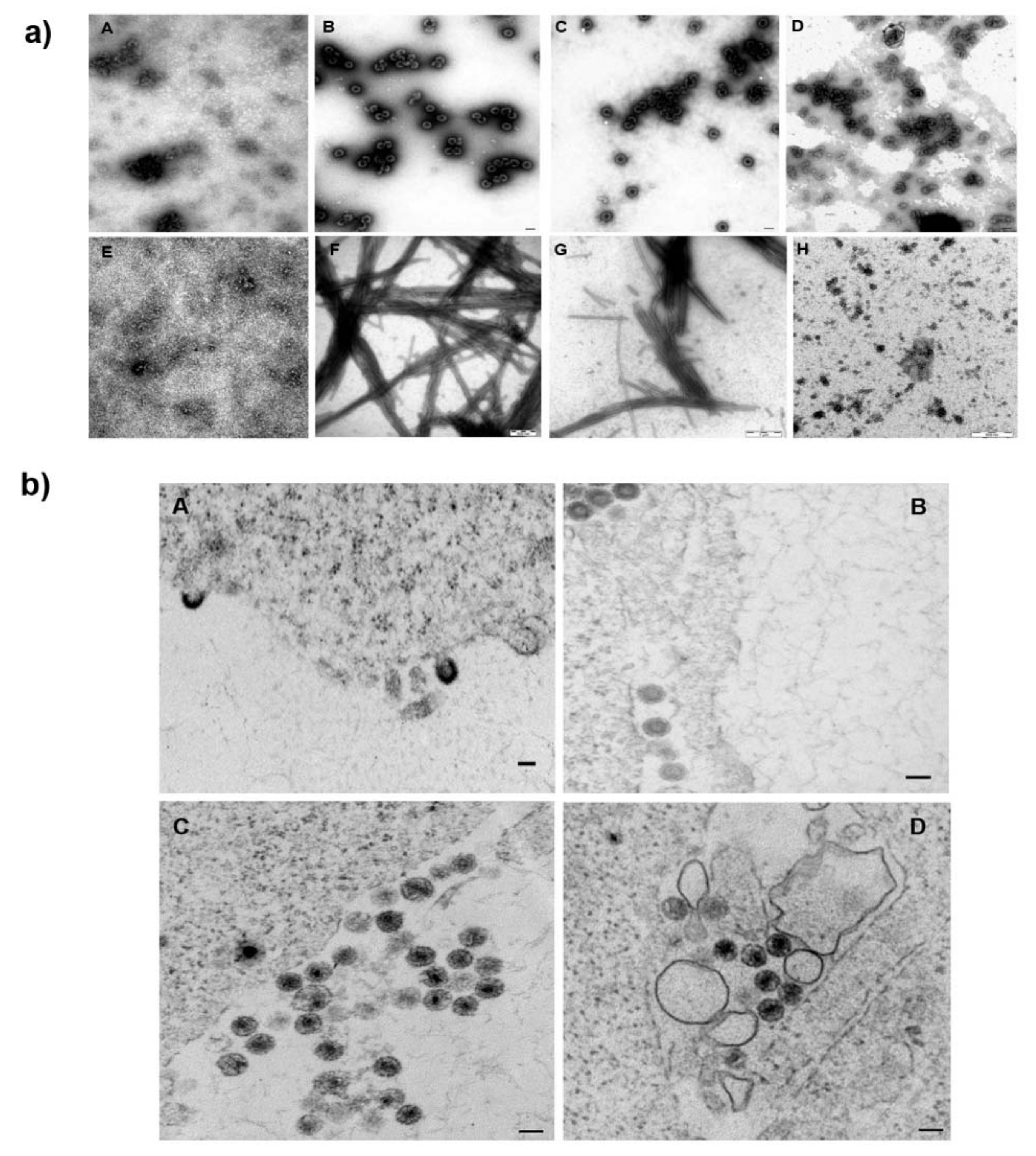
In their work, Castro et al. [
36] suggested a direct interaction between the fullerene inhibitor and HIV-1 capsid protein. As CA is the major structural protein critical for the formation of HIV-1’s immature and mature hexameric lattices and for uncoating, we next tested the possible direct binding of
1 to CA. We did not observe any effect of
1 on the HIV-1 assembly either by using an
in vitro assembly system or by TEM analysis of the thin sections of the HEK 293 cells producing HIV-1. No effect of
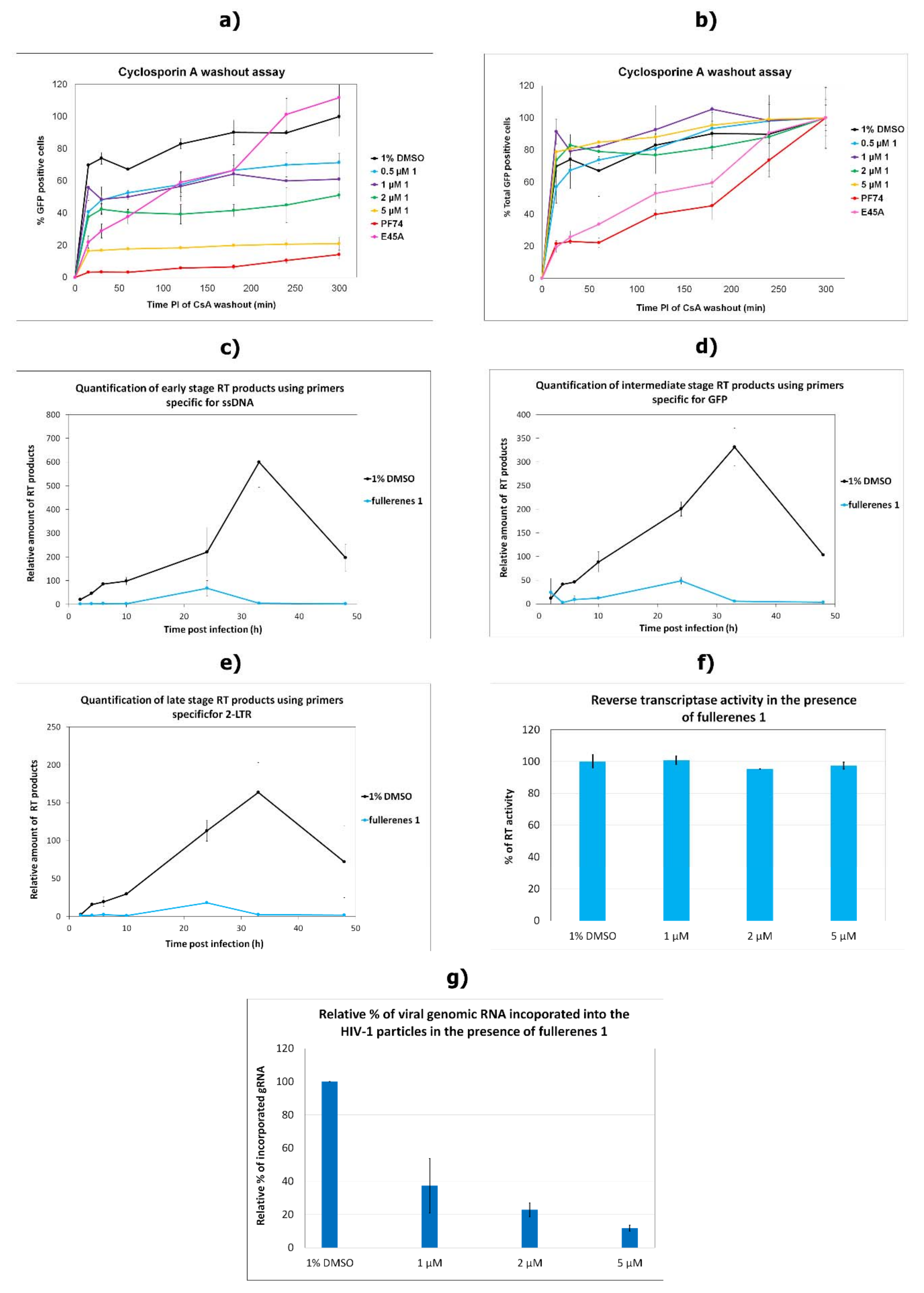
1 was detected in HIV-1 CA uncoating. The CsA washout assay, which monitors the HIV-1 capsid core uncoating, clearly showed a concentration-dependent effect of
1 on virus infectivity in OMK cells. However, the rate of uncoating, which would prove an effect of
1 on hexameric CA lattice’s core stability, was not affected by fullerene
1. In comparison to PF74 and E45A CA mutant, which were both shown to affect capsid hexameric stability [
39,
55,
57,
58,
59], fullerene
1 did not slow down the rate of uncoating. Finally, as MST analysis confirmed the low-binding affinity of
1 to HIV-1 CA, we concluded that
1 did not directly bind to HIV-1 CA.
Since the rate of uncoating seemed to be unaffected by fullerene
1, to understand its effect on reverse transcription, we studied the direct effect of fullerene
1 on RT activity. No inhibitory effect was observed, however, so we quantified the amount of viral genomic RNA incorporated into released HIV-1. Surprisingly, we found an inverse correlation between the concentration of
1 and the amount of viral genomic RNA. That was unexpected, because Martinez et al. did not observe any effect of fullerene inhibitors on RNA incorporation [
35]. In their work they normalized fullerene derivatives-treated virions by ELISA. However, we noticed that fullerene strongly interfered with the horseradish peroxidase (HRP) conversion of the 3,3′,5,5′-tetramethylbenzidine (TMB) substrate to its oxidized colored product (
Supplementary Figure S1a). Therefore,
1 might severely skew the ELISA results and subsequent experiments based on ELISA-normalized virions (such as virus-associated genomic RNA). To avoid this fullerene-associated bias, we used a semi-quantitative Western blot for virions normalization [
40]. Apart from
1’s interference with the TMB reaction, we also observed that during MST analysis,
1 quenched the fluorescence of its fluorescently labeled interacting partner (e.g., NHS-RED-labeled CANC). We saw no quenching of FAM or NHS-RED dye itself (
Supplementary Figure S1b). This observation clarified the problems connected to FAITH measurements, during which we observed fullerene
1 concentration-dependent fluorescence quenching of tqON (
Supplementary Figure S1c). This fluorescent quenching property of
1 could explain the discrepancies between our results and Martinez et al.’s results on the effect of
1 on HIV-1 RT activity [
35]. Using the assay with the chromogenic substrate (Merck), which is not affected by fullerene
1, we did not observe any impact of
1 on RT activity (
Figure 4g). In contrast, when Martinez et al. used the same assay, the formation of fluorescently labeled RNA–DNA heteroduplex was detected. As we showed by MST that
1 binds to FAM-labeled RNA or DNA and quenches their fluorescence, it is plausible that binding of
1 to fluorescently labeled nucleic acid decreased its fluorescence, which was then misinterpreted as reduced RT activity.
The logical explanation for how fullerene
1 affected viral genomic RNA incorporation could be its interference with NC–gRNA binding. We proved the affinity of
1 to CANC and NC by MST. CANC–fullerene and NC–fullerene
1 binding were also verified by EMSA, during which NC treated by
1 reduced its ability to interact with ssDNA in an inhibitor concentration-dependent manner. Although we did not observe the direct binding of HIV-1 CA to fullerene using MST or ELISA assays, in two experiments (
Figure 1d and
Figure 5b), a fuzzy-like band, supposedly corresponding to CA with bound fullerene
1, was observed. A similar fuzzy-like appearance of the HIV-1 CA band, as a result of fullerene inhibitor binding, was previously identified by Castro et al. [
36]. They performed a pull-down analysis by incubating fullerene inhibitors attached to magnetic beads with the lysate of HEK-293 cells producing HIV-1 Gag and Pol polyproteins [
36]. Based on the fuzzy-like appearance of this immunoprecipitated CA on the immunoblot, Castro et al. concluded that the molecular weight did not correspond to fully processed CAp24, but possibly to the CA-SP1p25 protein. The reason for these fuzzy bands in our Western blot (
Figure 1d) and Coomassie-blue stained gel (
Figure 5b) remains unclear. However, in contrast to bevirimat, we did not observe any effect of
1 on HIV-1 CA maturation by using immunoprecipitation of radioactively labelled HIV-1 Gag derived proteins (pulse-chase experiment). We therefore concluded that the fuzzy-like appearance is a technical artefact; nevertheless, it represented properly processed CAp24 protein. To confirm high preference of fullerene
1 in binding NC prior to CA, we performed docking experiments. We used two programs, PLANTS [
48] and AutoDock Vina, version 1.5.6rc3 [
49], to dock
1 into HIV-1 CA (3NTE) and NC (1F6U). In the case of CA, no preferential binding site was found, as both programs selected two different preferential binding sites, always in the ratio 80:20 (
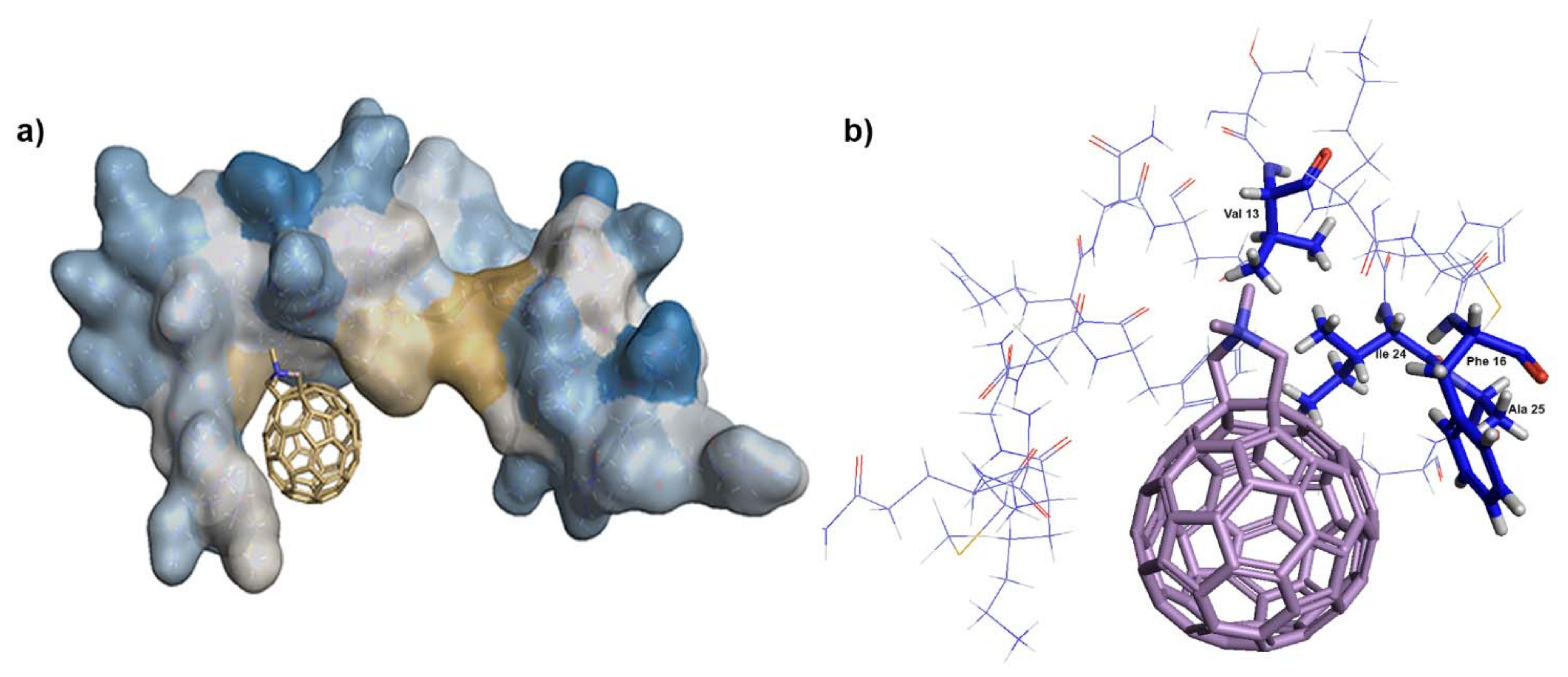
Supplementary Figure S2). In contrast, both programs placed
1 into the same position within the NC structure—into the vicinity of the hydrophobic residues Val13, Phe16, Ile24, and Ala25 of the proximal zinc finger (ZF) (
Figure 6a,b). Together with the hydrophobic residues of the distal ZF, Trp37, Gln45, and Met46, these amino acid residues form a hydrophobic plateau, which plays a key role in NC binding to nucleic acids [
18,
60,
61]. Even though we cannot completely rule out that fullerene
1 may also bind to HIV-1 CA hydrophobic pockets, our MST and EMSA data strongly argue that this interaction is insignificant and that fullerene
1 potently blocks infection by interfering in NC–gRNA interactions. The lowered amount of virus-incorporated gRNA then leads to a decrease in cDNA synthesis during reverse transcription and a significant reduction in HIV-1 infectivity.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





