
Prior Methamphetamine Use Disorder History Does Not Impair Interoceptive Processing of Soft Touch in HIV Infection

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measures
2.3. Imaging Procedures
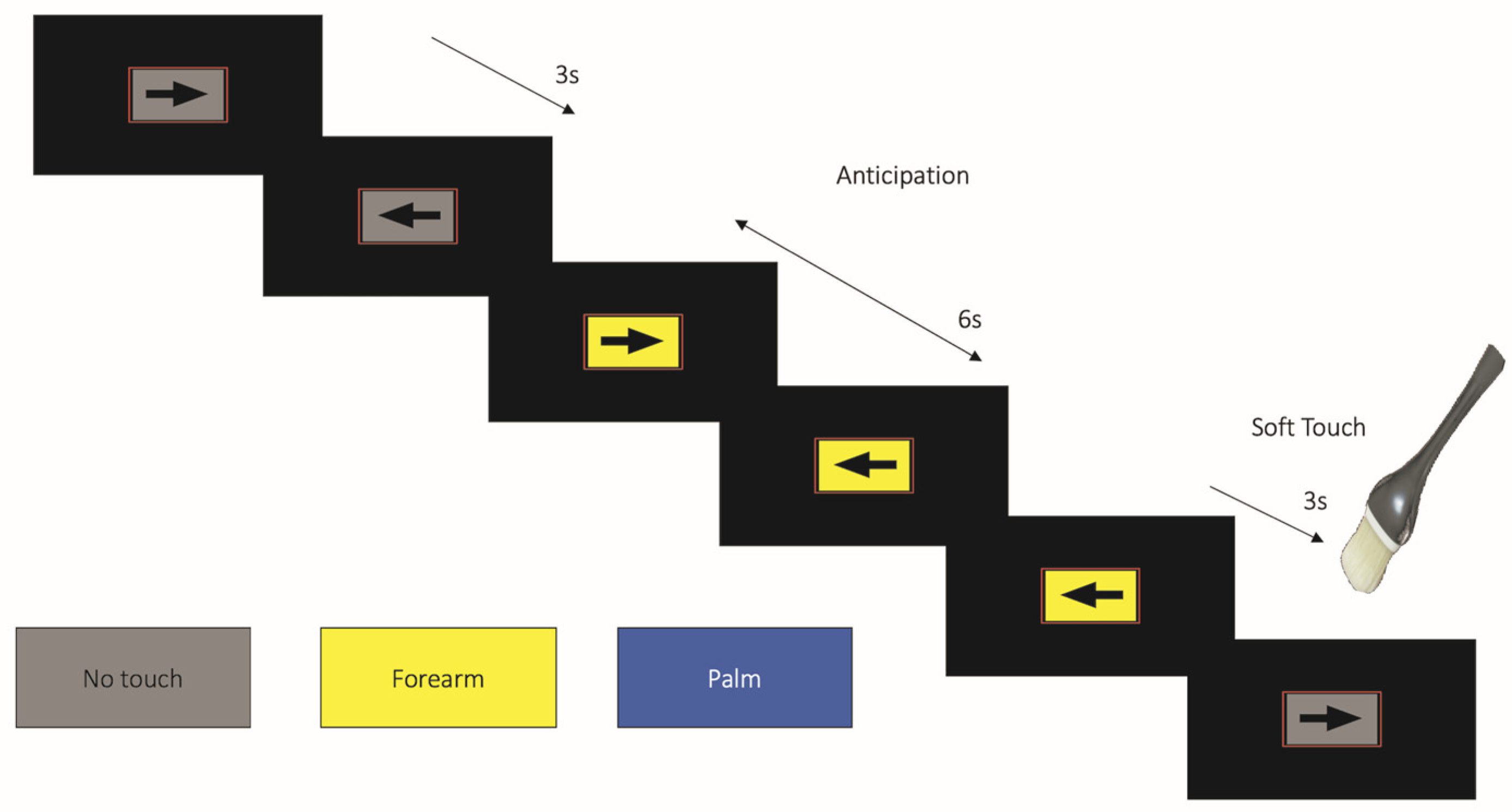
2.3.1. Soft Touch Paradigm
2.3.2. Image Acquisition
2.3.3. Image Preprocessing
2.4. Data Analysis
2.4.1. Behavioral Analysis
2.4.2. Regions of Interest
2.4.3. Neuroimaging Analysis
2.4.4. Primary Robust Regression Analyses
3. Results
3.1. Participant Characteristics
3.2. VAS Scales
3.3. Behavioral Analyses
3.4. Region of Interest Analyses
3.4.1. Main Effect of Condition
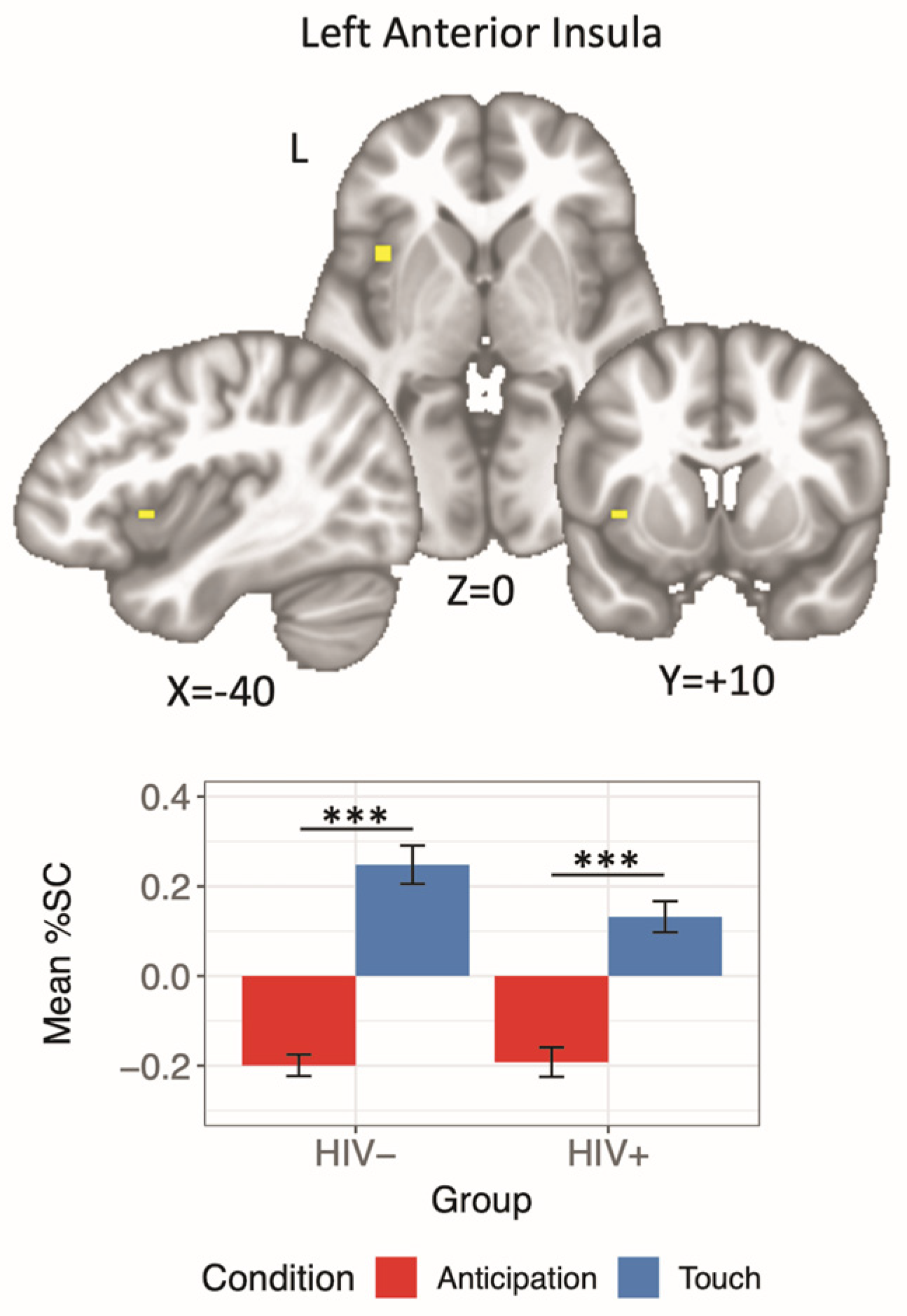
3.4.2. HIV × Condition Interaction
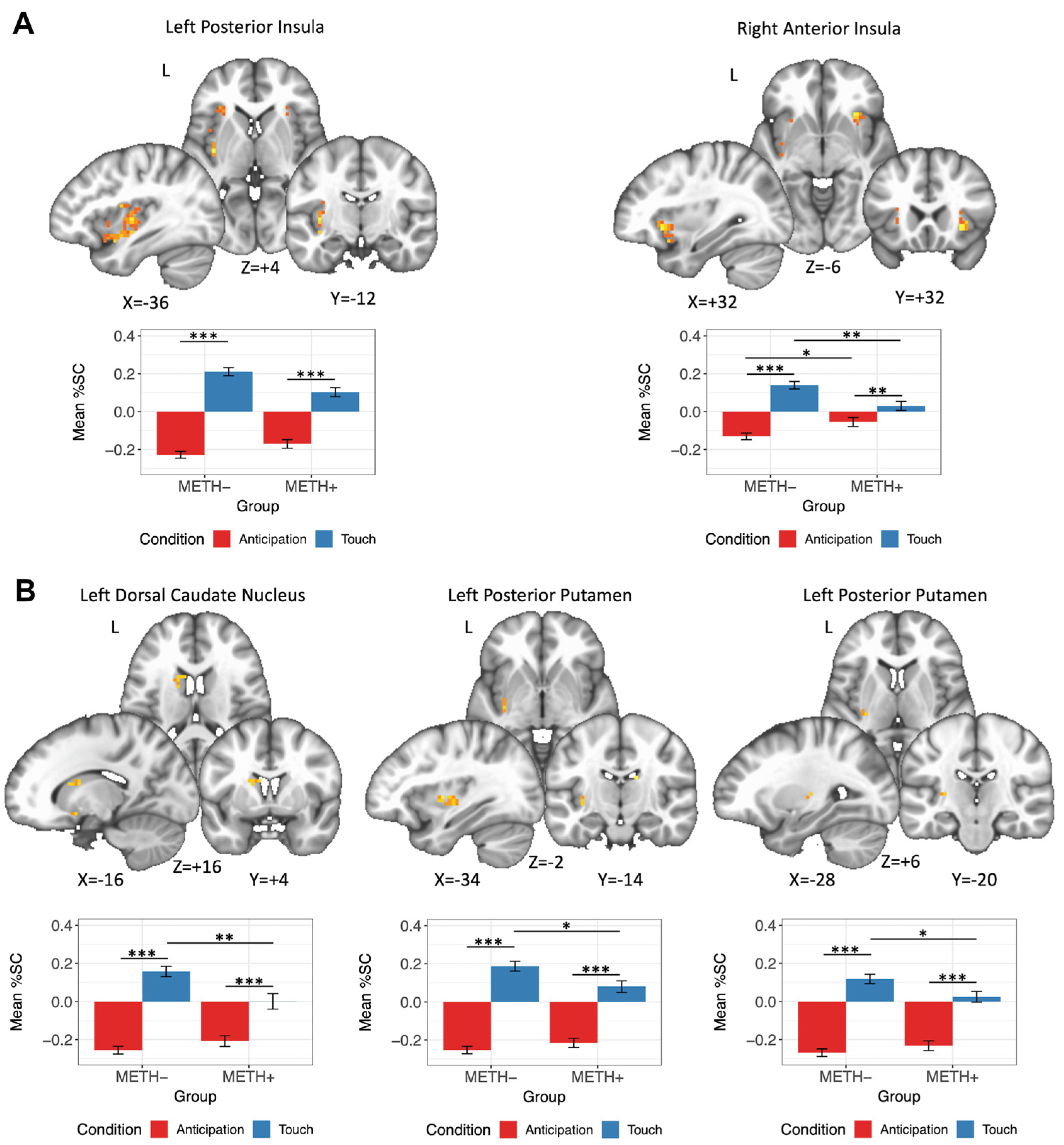
3.4.3. METH × Condition Interaction
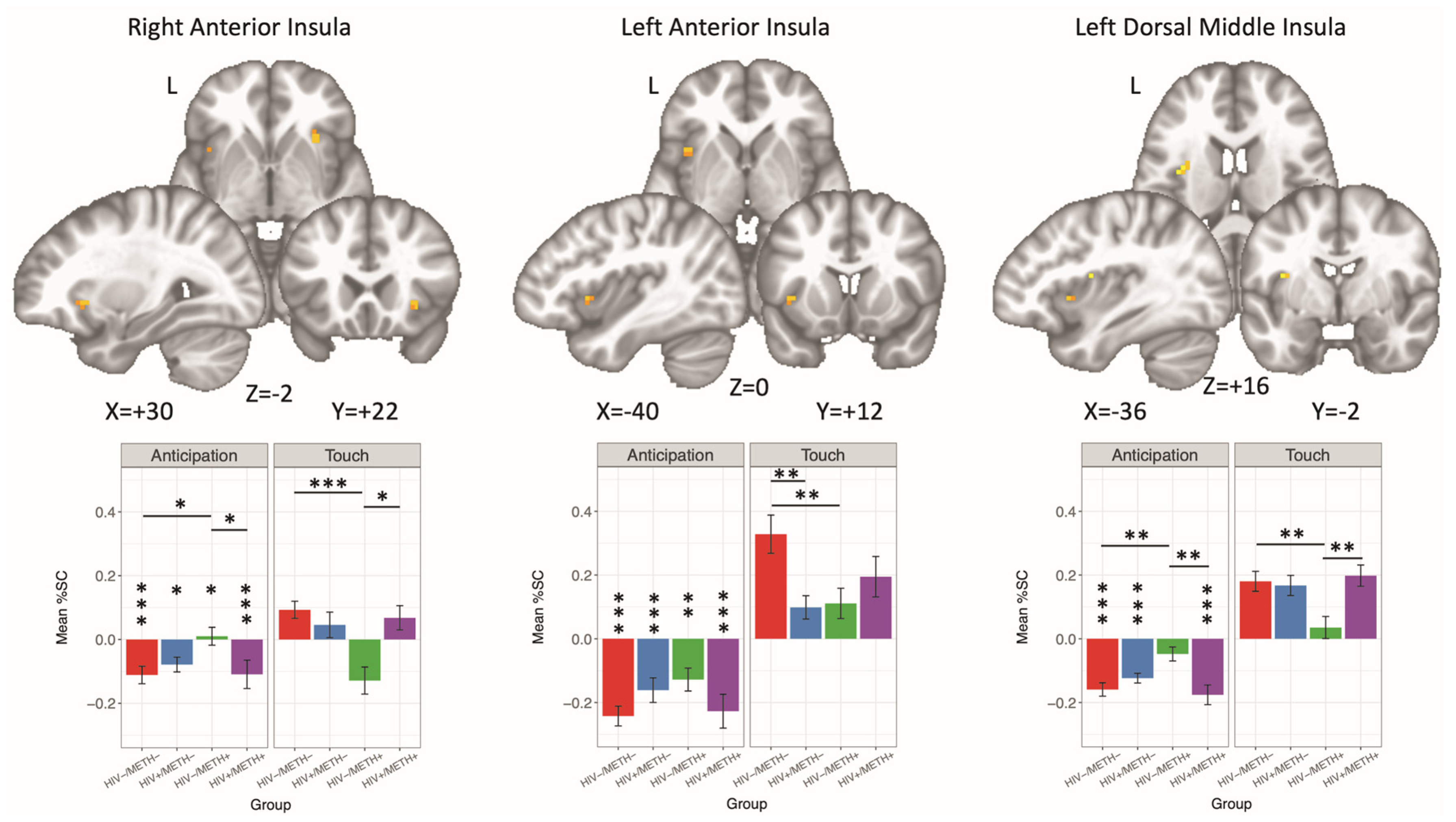
3.4.4. HIV × METH × Condition
3.5. Associations with Clinical Variables
3.5.1. PWH
3.5.2. Individuals with a History of METH Use Disorder
4. Discussion
4.1. Significance of Findings
4.2. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halkitis, P.N.; Parsons, J.T.; Stirratt, M.J. A double epidemic: Crystal methamphetamine drug use in relation to HIV transmission among gay men. J. Homosex. 2001, 41, 17–35. [Google Scholar] [CrossRef]
- Vu, N.T.; Maher, L.; Zablotska, I. Amphetamine-type stimulants and HIV infection among men who have sex with men: Implications on HIV research and prevention from a systematic review and meta-analysis. J. Int. AIDS Soc. 2015, 18, 19273. [Google Scholar] [CrossRef]
- Kuchinad, K.E.; Hutton, H.E.; Monroe, A.K.; Anderson, G.; Moore, R.D.; Chander, G. A qualitative study of barriers to and facilitators of optimal engagement in care among PLWH and substance use/misuse. BMC Res. Notes 2016, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.J.; Childers, M.E.; Cherner, M.; Lazzaretto, D.; Letendre, S.; the HIV Neurobehavioral Research Center Group. Increased human immunodeficiency virus loads in active methamphetamine users are explained by reduced effectiveness of antiretroviral therapy. J. Infect. Dis. 2003, 188, 1820–1826. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.J.; Blackstone, K.; Woods, S.P.; Ellis, R.J.; Atkinson, J.H.; Heaton, R.K.; Grant, I.; the HNRC Group; the TMARC Group. Methamphetamine use and neuropsychiatric factors are associated with antiretroviral non-adherence. AIDS Care 2012, 24, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.B.; Thomas, J.A.; Alexy, E.R.; Irvine, M.K. Crystal methamphetamine use and HIV medical outcomes among HIV-infected men who have sex with men accessing support services in New York. Drug Alcohol Depend. 2015, 147, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.K.; Lin, J.C.; Miles, S.W.; Kydd, R.R.; Russell, B.R. Striatal volume increases in active methamphetamine-dependent individuals and correlation with cognitive performance. Brain Sci. 2012, 2, 553–572. [Google Scholar] [CrossRef]
- Chang, L.; Andres, M.; Sadino, J.; Jiang, C.S.; Nakama, H.; Miller, E.; Ernst, T. Impact of apolipoprotein E ε4 and HIV on cognition and brain atrophy: Antagonistic pleiotropy and premature brain aging. Neuroimage 2011, 58, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Connolly, C.G.; Bischoff-Grethe, A.; Jordan, S.J.; Woods, S.P.; Ellis, R.J.; Paulus, M.P.; Grant, I.; the Translational Methamphetamine Aids Research Center (TMARC) Group. Altered functional response to risky choice in HIV infection. PLoS ONE 2014, 9, e111583. [Google Scholar] [CrossRef]
- Gowin, J.L.; Stewart, J.L.; May, A.C.; Ball, T.M.; Wittmann, M.; Tapert, S.F.; Paulus, M.P. Altered cingulate and insular cortex activation during risk-taking in methamphetamine dependence: Losses lose impact. Addiction 2014, 109, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Cohen, R.A.; Siegel, S.; Gullett, J.M.; Porges, E.; Woods, A.J.; Huang, H.; Zhu, Y.; Tashima, K.; Ding, M.Z. Neural response to working memory demand predicts neurocognitive deficits in HIV. J. Neurovirol. 2018, 24, 291–304. [Google Scholar] [CrossRef]
- London, E.D.; Kohno, M.; Morales, A.M.; Ballard, M.E. Chronic methamphetamine abuse and corticostriatal deficits revealed by neuroimaging. Brain Res. 2015, 1628, 174–185. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Ernst, T.; Speck, O.; Grob, C.S. Additive effects of HIV and chronic methamphetamine use on brain metabolite abnormalities. Am. J. Psychiatry 2005, 162, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ances, B.M.; Vaida, F.; Cherner, M.; Yeh, M.J.; Liang, C.L.; Gardner, C.; Grant, I.; Ellis, R.J.; Buxton, R.B.; the HIV Neurobehavioral Research Center (HNRC) Group. HIV and chronic methamphetamine dependence affect cerebral blood flow. J. Neuroimmune Pharmacol. 2011, 6, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Jernigan, T.L.; Gamst, A.C.; Archibald, S.L.; Fennema-Notestine, C.; Mindt, M.R.; Marcotte, T.D.; Heaton, R.K.; Ellis, R.J.; Grant, I. Effects of methamphetamine dependence and HIV infection on cerebral morphology. Am. J. Psychiatry 2005, 162, 1461–1472. [Google Scholar] [CrossRef] [Green Version]
- MacDuffie, K.E.; Brown, G.G.; McKenna, B.S.; Liu, T.T.; Meloy, M.J.; Tawa, B.; Archibald, S.; Fennema-Notestine, C.; Atkinson, J.H., Jr.; Ellis, R.J.; et al. Effects of HIV Infection, methamphetamine dependence and age on cortical thickness, area and volume. NeuroImage Clin. 2018, 20, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Archibald, S.L.; Jacobson, M.W.; Fennema-Notestine, C.; Ogasawara, M.; Woods, S.P.; Letendre, S.; Grant, I.; Jernigan, T.L. Functional interactions of HIV-infection and methamphetamine dependence during motor programming. Psychiatry Res. Neuroimaging 2012, 202, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Craig, A.D. Interoception: The sense of the physiological condition of the body. Curr. Opin. Neurobiol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- LaMotte, R.H.; Thalhammer, J.G.; Torebjork, H.E.; Robinson, C.J. Peripheral neural mechanisms of cutaneous hyperalgesia following mild injury by heat. J. Neurosci. 1982, 2, 765–781. [Google Scholar] [CrossRef]
- Craig, A.D.; Bushnell, M.C. The thermal grill illusion: Unmasking the burn of cold pain. Science 1994, 265, 252–255. [Google Scholar] [CrossRef]
- Olausson, H.; Lamarre, Y.; Backlund, H.; Morin, C.; Wallin, B.G.; Starck, G.; Ekholm, S.; Strigo, I.; Worsley, K.; Vallbo, A.B.; et al. Unmyelinated tactile afferents signal touch and project to insular cortex. Nat. Neurosci. 2002, 5, 900–904. [Google Scholar] [CrossRef]
- Vallbo, A.; Olausson, H.; Wessberg, J.; Norrsell, U. A system of unmyelinated afferents for innocuous mechanoreception in the human skin. Brain Res. 1993, 628, 301–304. [Google Scholar] [CrossRef]
- Craig, A.D. How do you feel? Interoception: The sense of the physiological condition of the body. Nat. Rev. Neurosci. 2002, 3, 655–666. [Google Scholar] [CrossRef]
- Chikama, M.; McFarland, N.R.; Amaral, D.G.; Haber, S.N. Insular cortical projections to functional regions of the striatum correlate with cortical cytoarchitectonic organization in the primate. J. Neurosci. 1997, 17, 9686–9705. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.M.; Zahm, D.S. Specificity in the projections of prefrontal and insular cortex to ventral striatopallidum and the extended amygdala. J. Neurosci. 2005, 25, 11757–11767. [Google Scholar] [CrossRef] [Green Version]
- Jasmin, L.; Burkey, A.R.; Granato, A.; Ohara, P.T. Rostral agranular insular cortex and pain areas of the central nervous system: A tract-tracing study in the rat. J. Comp. Neurol. 2004, 468, 425–440. [Google Scholar] [CrossRef]
- Klein, T.A.; Ullsperger, M.; Danielmeier, C. Error awareness and the insula: Links to neurological and psychiatric diseases. Front. Hum. Neurosci. 2013, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, A.D. Significance of the insula for the evolution of human awareness of feelings from the body. Ann. N. Y. Acad. Sci. 2011, 1225, 72–82. [Google Scholar] [CrossRef]
- May, A.C.; Stewart, J.L.; Migliorini, R.; Tapert, S.F.; Paulus, M.P. Methamphetamine dependent individuals show attenuated brain response to pleasant interoceptive stimuli. Drug Alcohol Depend. 2013, 131, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Paulus, M.P.; Stewart, J.L. Neurobiology, Clinical Presentation, and Treatment of Methamphetamine Use Disorder: A Review. JAMA Psychiatry 2020, 77, 959–966. [Google Scholar] [CrossRef]
- Walker, S.C.; McGlone, F.P. The social brain: Neurobiological basis of affiliative behaviours and psychological well-being. Neuropeptides 2013, 47, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Kalivas, P.W. The glutamate homeostasis hypothesis of addiction. Nat. Rev. Neurosci. 2009, 10, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; O’Neill, J.; Alger, J.R.; Shen, Z.; Johnson, M.C.; London, E.D. N-Acetyl and Glutamatergic Neurometabolites in Perisylvian Brain Regions of Methamphetamine Users. Int. J. Neuropsychopharmacol. 2019, 22, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.M.; Ownby, R.L.; Waldrop-Valverde, D.; Fernandez, B.; Kumar, M. Human immunodeficiency virus infection in the CNS and decreased dopamine availability: Relationship with neuropsychological performance. J. Neurovirology 2011, 17, 26–40. [Google Scholar] [CrossRef]
- Vázquez-Santiago, F.J.; Noel, R.J., Jr.; Porter, J.T.; Rivera-Amill, V. Glutamate metabolism and HIV-associated neurocognitive disorders. J. Neurovirology 2014, 20, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Umlauf, A.; Letendre, S.; Franklin, D.; Bush, W.S.; Atkinson, J.H.; Keltner, J.; Ellis, R.J. Catechol-O-methyltransferase polymorphism Val158Met is associated with distal neuropathic pain in HIV-associated sensory neuropathy. AIDS 2019, 33, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Chatterjea, D.; Martinov, T. Mast cells: Versatile gatekeepers of pain. Mol. Immunol. 2015, 63, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Forsythe, P.; Bienenstock, J. The mast cell-nerve functional unit: A key component of physiologic and pathophysiologic responses. Chem. Immunol. Allergy 2012, 98, 196–221. [Google Scholar] [CrossRef]
- Lowy, D.B.; Makker, P.G.S.; Moalem-Taylor, G. Cutaneous Neuroimmune Interactions in Peripheral Neuropathic Pain States. Front. Immunol. 2021, 12, 660203. [Google Scholar] [CrossRef] [PubMed]
- Aruldass, A.R.; Kitzbichler, M.G.; Morgan, S.E.; Lim, S.; Lynall, M.E.; Turner, L.; Vertes, P.; Wellcome Trust Consortium for Neuroimmunology of Mood Disorders and Alzheimer’s Disease (NIMA); Alzheimer’s, D.; Cavanagh, J.; et al. Dysconnectivity of a brain functional network was associated with blood inflammatory markers in depression. Brain Behav. Immun. 2021, 98, 299–309. [Google Scholar] [CrossRef]
- Savitz, J.; Harrison, N.A. Interoception and Inflammation in Psychiatric Disorders. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 514–524. [Google Scholar] [CrossRef]
- Ellis, R.J.; Diaz, M.; Sacktor, N.; Marra, C.; Collier, A.C.; Clifford, D.B.; Calcutt, N.; Fields, J.A.; Heaton, R.K.; Letendre, S.L.; et al. Predictors of worsening neuropathy and neuropathic pain after 12 years in people with HIV. Ann. Clin. Transl. Neurol. 2020, 7, 1166–1173. [Google Scholar] [CrossRef]
- Karpul, D.; McIntyre, S.; van Schaik, A.; Breen, P.P.; Heckmann, J.M. Vibrotactile sensitivity of patients with HIV-related sensory neuropathy: An exploratory study. Brain Behav. 2019, 9, e01184. [Google Scholar] [CrossRef]
- McGlone, F.; Wessberg, J.; Olausson, H. Discriminative and affective touch: Sensing and feeling. Neuron 2014, 82, 737–755. [Google Scholar] [CrossRef] [Green Version]
- Liljencrantz, J.; Olausson, H. Tactile C fibers and their contributions to pleasant sensations and to tactile allodynia. Front. Behav. Neurosci. 2014, 8, 37. [Google Scholar] [CrossRef] [Green Version]
- Robinson-Papp, J.; Gelman, B.B.; Grant, I.; Singer, E.; Gensler, G.; Morgello, S.; the National NeuroAIDS Tissue Consortium. Substance abuse increases the risk of neuropathy in an HIV-infected cohort. Muscle Nerve 2012, 45, 471–476. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM:VI-TR, 4th ed.; American Psychiatric Association: Washington, DC, USA, 2000. [Google Scholar]
- Kessler, R.C.; Üstün, T.B. The World Mental Health (WMH) Survey Initiative Version of the World Health Organization (WHO) Composite International Diagnostic Interview (CIDI). Int. J. Methods Psychiatr. Res. 2004, 13, 93–121. [Google Scholar] [CrossRef]
- Wilkinson, G.S.; Robertson, G.J. Wide Range Achievement Test 4 Professional Manual; Psychological Assessment Resources: Lutz, FL, USA, 2006. [Google Scholar]
- Heaton, R.K.; Clifford, D.B.; Franklin, D.R., Jr.; Woods, S.P.; Ake, C.; Vaida, F.; Ellis, R.J.; Letendre, S.L.; Marcotte, T.D.; Atkinson, J.H.; et al. HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study. Neurology 2010, 75, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Beck, A.; Steer, R.; Brown, G. Manual for the Beck Depression Inventory-II; Psychological Corporation: San Antonio, TX, USA, 1996. [Google Scholar]
- Löken, L.S.; Wessberg, J.; Morrison, I.; McGlone, F.; Olausson, H. Coding of pleasant touch by unmyelinated afferents in humans. Nat. Neurosci. 2009, 12, 547–548. [Google Scholar] [CrossRef]
- Migliorini, R.; Stewart, J.L.; May, A.C.; Tapert, S.F.; Paulus, M.P. What do you feel? Adolescent drug and alcohol users show altered brain response to pleasant interoceptive stimuli. Drug Alcohol Depend. 2013, 133, 661–668. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.L.; May, A.C.; Tapert, S.F.; Paulus, M.P. Hyperactivation to pleasant interoceptive stimuli characterizes the transition to stimulant addiction. Drug Alcohol Depend. 2015, 154, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Gountouna, V.-E.; Job, D.E.; McIntosh, A.M.; Moorhead, T.W.J.; Lymer, G.K.L.; Whalley, H.C.; Hall, J.; Waiter, G.D.; Brennan, D.; McGonigle, D.J.; et al. Functional Magnetic Resonance Imaging (fMRI) reproducibility and variance components across visits and scanning sites with a finger tapping task. Neuroimage 2010, 49, 552–560. [Google Scholar] [CrossRef]
- Cox, R.W. AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput. Biomed. Res. 1996, 29, 162–173. [Google Scholar] [CrossRef]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.; Woolrich, M.W.; Smith, S.M. Fsl. Neuroimage 2012, 62, 782–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, Z.S.; Glen, D.R.; Chen, G.; Beauchamp, M.S.; Desai, R.; Cox, R.W. A new method for improving functional-to-structural MRI alignment using local Pearson correlation. Neuroimage 2009, 44, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ségonne, F.; Dale, A.M.; Busa, E.; Glessner, M.; Salat, D.; Kahn, H.K.; Fischl, B. A hybrid approach to the skull stripping problem in MRI. Neuroimage 2004, 22, 1060–1075. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.L.R.; Jenkinson, M.; Smith, S. Non-Linear Registration, aka Spatial Normalisation; TR07JA2; FMRIB Centre: Oxford, UK, 2010. [Google Scholar]
- Jenkinson, M.; Smith, S. A global optimisation method for robust affine registration of brain images. Med Image Anal. 2001, 5, 143–156. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M.; DebRoy, S.; Sarkar, D.; Team, R.D.C. Nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1-109. 2013. Available online: https://cran.r-project.org/package=nlme (accessed on 26 April 2019).
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means. 2020. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.L.; Juavinett, A.L.; May, A.C.; Davenport, P.W.; Paulus, M.P. Do you feel alright? Attenuated neural processing of aversive interoceptive stimuli in current stimulant users. Psychophysiology 2015, 52, 249–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff-Grethe, A.; Wierenga, C.E.; Berner, L.A.; Simmons, A.N.; Bailer, U.; Paulus, M.P.; Kaye, W.H. Neural hypersensitivity to pleasant touch in women remitted from anorexia nervosa. Transl. Psychiatry 2018, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Eklund, A.; Nichols, T.E.; Knutsson, H. Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates. Proc. Natl. Acad. Sci. USA 2016, 113, 7900–7905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Yoshihara, Y.; Watanabe, D.; Fukumoto, M.; Wada, K.; Nakakura, T.; Kuriyama, K.; Shirasaka, T.; Murai, T. Neurocognitive impairment and gray matter volume reduction in HIV-infected patients. J. Neurovirol. 2020, 26, 590–601. [Google Scholar] [CrossRef]
- Keltner, J.R.; Connolly, C.G.; Vaida, F.; Jenkinson, M.; Fennema-Notestine, C.; Archibald, S.; Akkari, C.; Schlein, A.; Lee, J.; Wang, D.; et al. HIV Distal Neuropathic Pain Is Associated with Smaller Ventral Posterior Cingulate Cortex. Pain Med. 2017, 18, 428–440. [Google Scholar] [CrossRef] [Green Version]
- Keltner, J.R.; Fennema-Notestine, C.; Vaida, F.; Wang, D.; Franklin, D.R.; Dworkin, R.H.; Sanders, C.; McCutchan, J.A.; Archibald, S.L.; Miller, D.J.; et al. HIV-associated distal neuropathic pain is associated with smaller total cerebral cortical gray matter. J. Neurovirol. 2014, 20, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Keltner, J.R.; Tong, A.; Visser, E.; Jenkinson, M.; Connolly, C.G.; Dasca, A.; Sheringov, A.; Calvo, Z.; Umbao, E.; Mande, R.; et al. Evidence for a novel subcortical mechanism for posterior cingulate cortex atrophy in HIV peripheral neuropathy. J. Neurovirol. 2020, 26, 530–543. [Google Scholar] [CrossRef]
- Bak, Y.; Jun, S.; Choi, J.Y.; Lee, Y.; Lee, S.-K.; Han, S.; Shin, N.-Y. Altered intrinsic local activity and cognitive dysfunction in HIV patients: A resting-state fMRI study. PLoS ONE 2018, 13, e0207146. [Google Scholar] [CrossRef] [PubMed]
- Chaganti, J.R.; Heinecke, A.; Gates, T.M.; Moffat, K.J.; Brew, B.J. Functional Connectivity in Virally Suppressed Patients with HIV-Associated Neurocognitive Disorder: A Resting-State Analysis. AJNR Am. J. Neuroradiol. 2017, 38, 1623–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, J.B.; Brier, M.R.; Snyder, A.Z.; Vaida, F.F.; Ances, B.M. Pathways to neurodegeneration: Effects of HIV and aging on resting-state functional connectivity. Neurology 2013, 80, 1186–1193. [Google Scholar] [CrossRef] [Green Version]
- Guha, A.; Wang, L.; Tanenbaum, A.; Esmaeili-Firidouni, P.; Wendelken, L.A.; Busovaca, E.; Clifford, K.; Desai, A.; Ances, B.M.; Valcour, V. Intrinsic network connectivity abnormalities in HIV-infected individuals over age 60. J. Neurovirol. 2016, 22, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Ortega, M.; Brier, M.R.; Ances, B.M. Effects of HIV and combination antiretroviral therapy on cortico-striatal functional connectivity. AIDS 2015, 29, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strigo, I.A.; Keltner, J.; Ellis, R.; Simmons, A.N. Association of Painful Human Immunodeficiency Virus Distal Sensory Polyneuropathy with Aberrant Expectation of Pain Relief: Functional Magnetic Resonance Imaging Evidence. Brain Commun. 2021, 3, fcab260. [Google Scholar] [CrossRef] [PubMed]
- Namkung, H.; Kim, S.-H.; Sawa, A. The Insula: An Underestimated Brain Area in Clinical Neuroscience, Psychiatry, and Neurology. Trends Neurosci. 2017, 40, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff-Grethe, A.; Connolly, C.G.; Jordan, S.J.; Brown, G.G.; Paulus, M.P.; Tapert, S.F.; Heaton, R.K.; Woods, S.P.; Grant, I.; the TMARC Group. Altered reward expectancy in individuals with recent methamphetamine dependence. J. Psychopharmacol. 2017, 31, 17–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Völlm, B.A.; de Araujo, I.E.; Cowen, P.J.; Rolls, E.T.; Kringelbach, M.L.; Smith, K.A.; Jezzard, P.; Heal, R.J.; Matthews, P.M. Methamphetamine activates reward circuitry in drug naive human subjects. Neuropsychopharmacology 2004, 29, 1715–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdejo-Garcia, A.; Clark, L.; Dunn, B.D. The role of interoception in addiction: A critical review. Neurosci. Biobehav. Rev. 2012, 36, 1857–1869. [Google Scholar] [CrossRef]
- Stewart, J.L.; Khalsa, S.S.; Kuplicki, R.; Puhl, M.; Investigators, T.; Paulus, M.P. Interoceptive attention in opioid and stimulant use disorder. Addict. Biol. 2020, 25, e12831. [Google Scholar] [CrossRef]
- Nestor, L.J.; Ghahremani, D.G.; Monterosso, J.; London, E.D. Prefrontal hypoactivation during cognitive control in early abstinent methamphetamine-dependent subjects. Psychiatry Res. Neuroimaging 2011, 194, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.L.; Connolly, C.G.; May, A.C.; Tapert, S.F.; Wittmann, M.; Paulus, M.P. Striatum and insula dysfunction during reinforcement learning differentiates abstinent and relapsed methamphetamine-dependent individuals. Addiction 2014, 109, 460–471. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.J.; Schweinsburg, B.C.; Alhassoon, O.M.; Gongvatana, A.; Brown, G.G.; Young-Casey, C.; Letendre, S.L.; Grant, I. Effects of human immunodeficiency virus and methamphetamine on cerebral metabolites measured with magnetic resonance spectroscopy. J. Neurovirol. 2007, 13, 150–159. [Google Scholar] [CrossRef]
- Nagarajan, R.; Sarma, M.K.; Thomas, M.A.; Chang, L.; Natha, U.; Wright, M.; Hayes, J.; Nielsen-Saines, K.; Michalik, D.E.; Deville, J.; et al. Neuropsychological function and cerebral metabolites in HIV-infected youth. J. Neuroimmune Pharmacol. 2012, 7, 981–990. [Google Scholar] [CrossRef]
- Illenberger, J.M.; Harrod, S.B.; Mactutus, C.F.; McLaurin, K.A.; Kallianpur, A.; Booze, R.M. HIV Infection and Neurocognitive Disorders in the Context of Chronic Drug Abuse: Evidence for Divergent Findings Dependent upon Prior Drug History. J Neuroimmune Pharmacol. 2020, 15, 715–728. [Google Scholar] [CrossRef]
- Rippeth, J.D.; Heaton, R.K.; Carey, C.L.; Marcotte, T.D.; Moore, D.J.; Gonzalez, R.; Wolfson, T.; Grant, I.; the HNRC Group. Methamphetamine dependence increases risk of neuropsychological impairment in HIV infected persons. J. Int. Neuropsychol. Soc. 2004, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.E.; Ipser, J.C.; Stein, D.J.; Joska, J.A.; Naudé, P.J.W. Peripheral immune dysregulation in the ART era of HIV-associated neurocognitive impairments: A systematic review. Psychoneuroendocrinology 2020, 118, 104689. [Google Scholar] [CrossRef] [PubMed]
- Assis, M.A.; Carranza, P.G.; Ambrosio, E. A “Drug-Dependent” Immune System Can Compromise Protection against Infection: The Relationships between Psychostimulants and HIV. Viruses 2021, 13, 722. [Google Scholar] [CrossRef] [PubMed]
- Matt, S.M.; Gaskill, P.J. Where Is Dopamine and how do Immune Cells See it?: Dopamine-Mediated Immune Cell Function in Health and Disease. J. Neuroimmune Pharmacol. 2020, 15, 114–164. [Google Scholar] [CrossRef]
- Kass-Iliyya, L.; Leung, M.; Marshall, A.; Trotter, P.; Kobylecki, C.; Walker, S.; Gosal, D.; Jeziorska, M.; Malik, R.A.; McGlone, F.; et al. The perception of affective touch in Parkinson’s disease and its relation to small fibre neuropathy. Eur. J. Neurosci. 2017, 45, 232–237. [Google Scholar] [CrossRef]
- Ernst, T.; Chang, L. Adaptation of brain glutamate plus glutamine during abstinence from chronic methamphetamine use. J. Neuroimmune Pharmacol. 2008, 3, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Volkow, N.D.; Wang, G.-J.; Smith, L.; Fowler, J.S.; Telang, F.; Logan, J.; Tomasi, D. Recovery of dopamine transporters with methamphetamine detoxification is not linked to changes in dopamine release. Neuroimage 2015, 121, 20–28. [Google Scholar] [CrossRef]
- Nance, R.M.; Trejo, M.E.P.; Whitney, B.M.; Delaney, J.A.C.; Altice, F.L.; Beckwith, C.G.; Chander, G.; Chandler, R.; Christopoulous, K.; Cunningham, C.; et al. Impact of Abstinence and of Reducing Illicit Drug Use Without Abstinence on Human Immunodeficiency Virus Viral Load. Clin. Infect. Dis. 2020, 70, 867–874. [Google Scholar] [CrossRef]
- Iudicello, J.E.; Morgan, E.E.; Gongvatana, A.; Letendre, S.L.; Grant, I.; Woods, S.P.; the Translational Methamphetamine AIDS Research Center (TMARC) Group. Detrimental impact of remote methamphetamine dependence on neurocognitive and everyday functioning in older but not younger HIV+ adults: Evidence for a legacy effect? J. Neurovirology 2014, 20, 85–98. [Google Scholar] [CrossRef]
- Meade, C.S.; Cordero, D.M.; Hobkirk, A.L.; Metra, B.M.; Chen, N.-K.; Huettel, S.A. Compensatory activation in fronto-parietal cortices among HIV-infected persons during a monetary decision-making task. Hum. Brain Mapp. 2016, 37, 2455–2467. [Google Scholar] [CrossRef] [Green Version]
- Hassanzadeh-Behbahani, S.; Shattuck, K.F.; Bronshteyn, M.; Dawson, M.; Diaz, M.; Kumar, P.; Moore, D.J.; Ellis, R.J.; Jiang, X. Low CD4 nadir linked to widespread cortical thinning in adults living with HIV. NeuroImage: Clin. 2020, 25, 102155. [Google Scholar] [CrossRef] [PubMed]
- Guha, A.; Brier, M.R.; Ortega, M.; Westerhaus, E.; Nelson, B.; Ances, B.M. Topographies of Cortical and Subcortical Volume Loss in HIV and Aging in the cART Era. J. Acquir. Immune Defic. Syndr. 2016, 73, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Jernigan, T.L.; Archibald, S.L.; Fennema-Notestine, C.; Taylor, M.J.; Theilmann, R.J.; Julaton, M.D.; Notestine, R.J.; Wolfson, T.; Letendre, S.L.; Ellis, R.J.; et al. Clinical factors related to brain structure in HIV: The CHARTER study. J. Neurovirol. 2011, 17, 248–257. [Google Scholar] [CrossRef]
- Ances, B.M.; Ortega, M.; Vaida, F.; Heaps, J.; Paul, R. Independent effects of HIV, aging, and HAART on brain volumetric measures. J. Acquir. Immune. Defic. Syndr. 2012, 59, 469–477. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, E.E.; Zeffiro, T.; Lopez, O.L.; Becker, J.T.; Zeffiro, T. HIV infection and age effects on striatal structure are additive. J. Neurovirol. 2019, 25, 480–495. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.A.; Harezlak, J.; Schifitto, G.; Hana, G.; Clark, U.; Gongvatana, A.; Paul, R.; Taylor, M.; Thompson, P.; Alger, J.; et al. Effects of nadir CD4 count and duration of human immunodeficiency virus infection on brain volumes in the highly active antiretroviral therapy era. J. Neurovirol. 2010, 16, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Montoya, J.L.; Cattie, J.; Morgan, E.; Woods, S.P.; Cherner, M.; Moore, D.J.; Atkinson, J.H.; Grant, I.; the Translational Methamphetamine Aids Research Center Group. The impact of age, HIV serostatus and seroconversion on methamphetamine use. Am. J. Drug Alcohol Abus. 2016, 42, 168–177. [Google Scholar] [CrossRef]
- Johansen-Berg, H.; Christensen, V.; Woolrich, M.; Matthews, P.M. Attention to touch modulates activity in both primary and secondary somatosensory areas. Neuroreport 2000, 11, 1237–1241. [Google Scholar] [CrossRef] [Green Version]
- Macaluso, E.; Driver, J. Spatial attention and crossmodal interactions between vision and touch. Neuropsychologia 2001, 39, 1304–1316. [Google Scholar] [CrossRef]
- Triscoli, C.; Ackerley, R.; Sailer, U. Touch satiety: Differential effects of stroking velocity on liking and wanting touch over repetitions. PLoS ONE 2014, 9, e113425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellingsen, D.M.; Leknes, S.; Løseth, G.; Wessberg, J.; Olausson, H. The Neurobiology Shaping Affective Touch: Expectation, Motivation, and Meaning in the Multisensory Context. Front. Psychol. 2015, 6, 1986. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
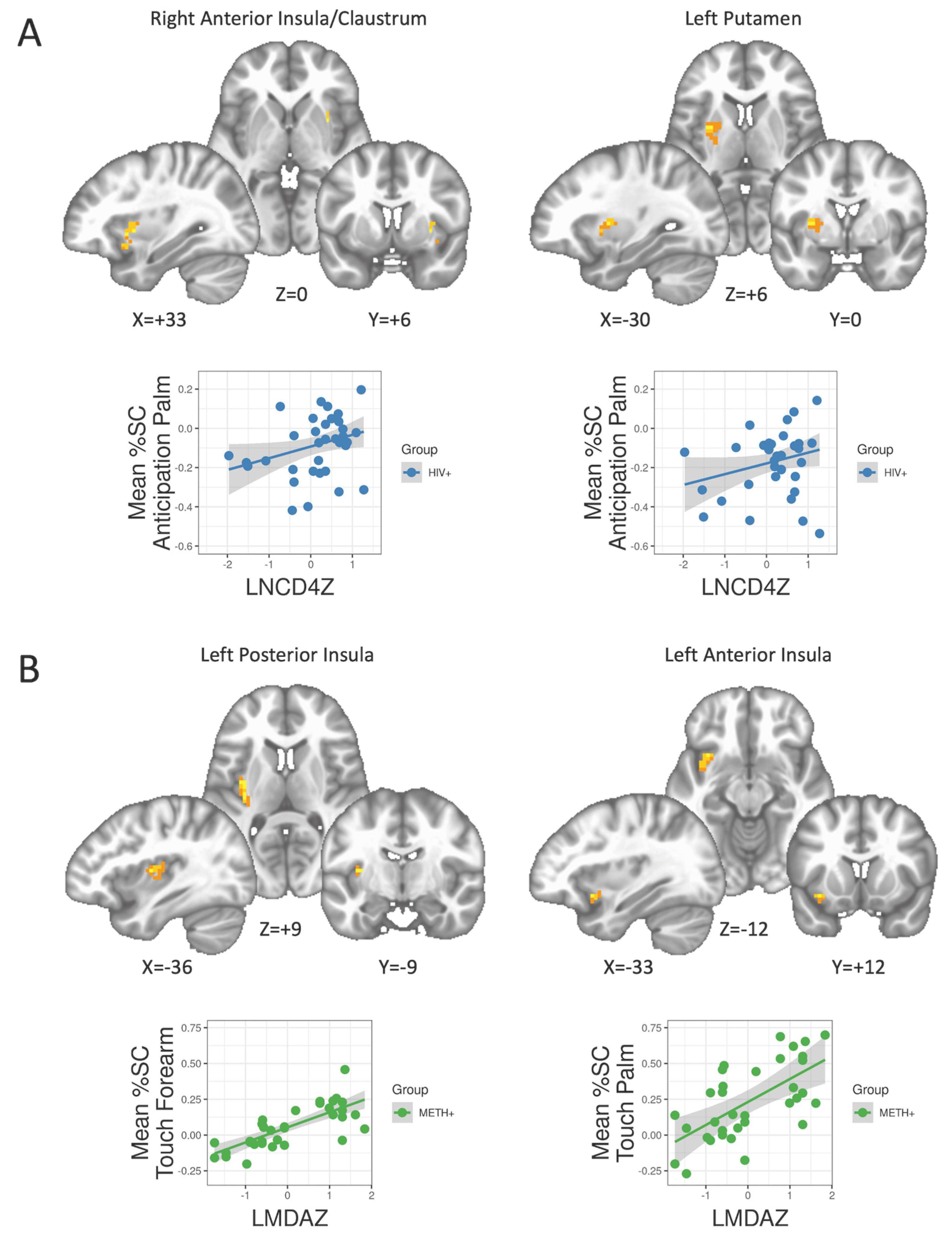{kind=link}
| Characteristic | HIV−/METH− (N = 28) | HIV+/METH− (N = 20) | HIV−/METH+ (N = 19) | HIV+/METH+ (N = 16) | p-Value |
|---|---|---|---|---|---|
| Age (years) | 38.8 (11.2) | 40.1 (11.3) | 37.2 (8.66) | 41.5 (10.1) | NS |
| Education (years) | 14.1 (2.05) | 13.8 (2.47) | 12.4 (2.31) | 13.4 (1.78) | 0.02 |
| Male/Female ☨ | 24/4 | 18/2 | 18/1 | 16/0 | NS |
| Handedness Right/Left ☨ | 27/1 | 17/3 | 18/1 | 16/0 | NS |
| Ethnicity (% Caucasian) ☨ | 64 | 50 | 53 | 63 | NS |
| Wide Range Achievement Test-4 Standard Score | 53.79 (9.12) | 49.55 (6.90) | 49.16 (7.53) | 53.88 (8.06) | 0.015 |
| Distal Symmetric Polyneuropathy (n) ☨ | 2 | 6 | 2 | 7 | 0.01 |
| Loss of Sensation (n) ☨ | 0 | 3 | 0 | 4 | 0.004 |
| Paresthesia (n) ☨ | 0 | 4 | 1 | 5 | 0.003 |
| Dysesthesia (n) ☨ | 1 | 1 | 1 | 4 | NS |
| Neuropathic pain (n) ☨ | 1 | 1 | 1 | 4 | NS |
| Beck Depression Inventory II a | 2.36 (3.23) | 7.70 (8.57) | 13.2 (9.28) | 12.4 (12.6) | <0.001 |
| GE Signa/GE MR750 System ☨ | 10/18 | 3/17 | 11/8 | 6/10 | 0.05 |
| Methamphetamine Characteristics | |||||
| Age of First Use | 24.7 (8.36) | 25.1 (7.80) | NS | ||
| Total Days Used | 1934 (1511) | 1720 (2269) | NS | ||
| Days Since Last Use a | 182 (160) | 257 (190) | NS | ||
| Total Quantity (g) | 2733 (3290) | 1335 (2563) | NS | ||
| Use Density | 1.28 (1.34) | 0.75 (0.75) | NS | ||
| Primary Route of Use ☨ | Smoking | Smoking | NS | ||
| HIV Characteristics | |||||
| Duration of Infection (months) b | 113.2 (100.6) | 139.3 (106.5) | NS | ||
| Current antiretroviral use ☨ | 14 | 12 | NS | ||
| Nadir CD4 Count | 250.0 (170.5) | 196.5 (171.2) | NS | ||
| Current CD4 Count c | 447.5 (283.2) | 543.0 (259.5) | NS |
| Post Hoc Comparisons | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Structure | Volume (μL) | X | Y | Z | F(peak) | Contrast | t Ratio | p-Value | ES |
| MAIN EFFECT OF CONDITION | |||||||||
| Left Insula Lobe | 14769 | −36 | −12 | 4 | 133.42 | Touch > Condition | 11.67 | <0.001 | 0.76 |
| Right Insula Lobe | 13716 | 36 | 16 | −14 | 70.63 | Touch > Condition | 9.71 | <0.001 | 0.56 |
| Left Striatum | 15849 | −28 | −8 | −8 | 124.88 | Touch > Condition | 10.16 | <0.001 | 0.71 |
| Right Striatum | 11880 | 12 | −6 | 24 | 73.95 | Touch > Condition | 8.76 | <0.001 | 0.51 |
| HIV × CONDITION | |||||||||
| Left Insula | 108 | −40 | 10 | 0 | 13.52 | HIV−: Touch > Anticipation | 9.25 | <0.001 | 0.66 |
| HIV+: Touch > Anticipation | 6.60 | <0.001 | 0.53 | ||||||
| METH × CONDITION | |||||||||
| Left Posterior Insula | 2322 | −36 | −12 | 4 | 22.57 | METH−: Touch > Anticipation | 15.44 | <0.001 | 0.99 |
| METH+: Touch > Anticipation | 8.75 | <0.001 | 0.65 | ||||||
| Touch: METH− > METH+ | 3.20 | 0.003 | 0.22 | ||||||
| Right Anterior Insula | 1350 | 32 | 22 | −6 | 26.53 | METH−: Touch > Anticipation | 9.55 | <0.001 | 0.60 |
| METH+: Touch > Anticipation | 3.01 | 0.004 | 0.22 | ||||||
| Anticipation: METH+ > METH− | 2.26 | 0.03 | 0.15 | ||||||
| Touch: METH− > METH+ | 3.32 | 0.003 | 0.23 | ||||||
| Left Anterior Insula | 486 | −28 | 24 | 12 | 18.10 | METH−: Touch > Anticipation | 6.76 | <0.001 | 0.70 |
| METH+: Touch > Anticipation | 2.49 | 0.03 | 0.48 | ||||||
| Left Dorsal Insula | 270 | −34 | 0 | 16 | 21.74 | METH−: Touch > Anticipation | 12.66 | <0.001 | 0.57 |
| METH+: Touch > Anticipation | 7.38 | <0.001 | 0.34 | ||||||
| Left Ventral Anterior Insula | 243 | −30 | 16 | −8 | 17.89 | METH−: Touch > Anticipation | 10.97 | <0.001 | 0.74 |
| METH+: Touch > Anticipation | 5.64 | <0.001 | 0.55 | ||||||
| Left Anterior Insula | 189 | −40 | 10 | 0 | 14.67 | METH−: Touch > Anticipation | 10.72 | <0.001 | 0.66 |
| METH+: Touch > Anticipation | 6.91 | <0.001 | 0.35 | ||||||
| Left Dorsal Caudate Nucleus | 486 | −16 | 4 | 16 | 15.92 | METH−: Touch > Anticipation | 12.21 | <0.001 | 0.92 |
| METH+: Touch > Anticipation | 5.59 | <0.001 | 0.64 | ||||||
| Touch: METH− > METH+ | 3.36 | 0.002 | 0.21 | ||||||
| Left Posterior Putamen/Claustrum | 243 | −34 | −14 | −2 | 14.32 | METH−: Touch > Anticipation | 13.29 | <0.001 | 0.69 |
| METH+: Touch > Anticipation | 8.01 | <0.001 | 0.48 | ||||||
| Touch: METH− > METH+ | 2.82 | 0.01 | 0.15 | ||||||
| Left Posterior Putamen | 162 | −28 | −20 | 6 | 14.40 | METH−: Touch > Anticipation | 12.18 | <0.001 | 0.65 |
| METH+: Touch > Anticipation | 7.34 | <0.001 | 0.41 | ||||||
| Touch: METH− > METH+ | 2.28 | 0.03 | 0.11 | ||||||
| Left Ventral Striatum | 135 | −16 | 6 | −12 | 14.25 | METH−: Touch > Anticipation | 9.29 | <0.001 | 0.65 |
| METH+: Touch > Anticipation | 5.00 | <0.001 | 0.41 | ||||||
| Left Posterior Putamen | 135 | −34 | −6 | 4 | 18.44 | METH−: Touch > Anticipation | 12.15 | <0.001 | 0.84 |
| METH+: Touch > Anticipation | 7.42 | <0.001 | 0.59 | ||||||
| HIV × METH × CONDITION | |||||||||
| Right Anterior Insula | 189 | 30 | 22 | −2 | 15.09 | Anticipation, HIV−: METH+ > METH− | 2.67 | 0.02 | 0.24 |
| Touch, HIV−: METH− > METH+ | 4.88 | <0.001 | 0.44 | ||||||
| HIV−/METH−: Touch > Anticipation | 5.08 | <0.001 | 0.41 | ||||||
| HIV+/METH−: Touch > Anticipation | 2.61 | 0.02 | 0.25 | ||||||
| HIV−/METH+: Anticipation > Touch | 2.85 | 0.01 | 0.28 | ||||||
| HIV+/METH+: Touch > Anticipation | 3.33 | <0.001 | 0.35 | ||||||
| Anticipation, METH+: HIV− > HIV+ | 2.30 | 0.04 | 0.24 | ||||||
| Touch, METH+: HIV+ > HIV− | 3.79 | 0.001 | 0.39 | ||||||
| Left Anterior Insula | 135 | −40 | 12 | 0 | 13.10 | Touch, HIV−: METH− > METH+ | 3.28 | 0.003 | 0.34 |
| HIV−/METH−: Touch > Anticipation | 10.21 | <0.001 | 0.90 | ||||||
| HIV+/METH−: Touch > Anticipation | 3.93 | <0.001 | 0.41 | ||||||
| HIV−/METH+: Touch > Anticipation | 3.52 | 0.002 | 0.38 | ||||||
| HIV+/METH+: Touch > Anticipation | 5.71 | <0.001 | 0.66 | ||||||
| Touch, METH−: HIV− > HIV+ | 3.52 | 0.002 | 0.36 | ||||||
| Left Dorsal Middle Insula | 135 | −36 | −2 | 16 | 17.37 | Anticipation, HIV−: METH+ > METH− | 2.86 | 0.01 | 0.23 |
| Touch, HIV−: METH− > METH+ | 3.71 | 0.001 | 0.30 | ||||||
| HIV−/METH−: Touch > Anticipation | 10.08 | <0.001 | 0.71 | ||||||
| HIV+/METH−: Touch > Anticipation | 7.30 | <0.001 | 0.61 | ||||||
| HIV+/METH+: Touch > Anticipation | 8.40 | <0.001 | 0.78 | ||||||
| Anticipation, METH+: HIV− > HIV+ | 2.88 | 0.01 | 0.27 | ||||||
| Touch, METH+: HIV+ > HIV− | 3.65 | 0.001 | 0.34 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bischoff-Grethe, A.; Ellis, R.J.; Tapert, S.F.; Paulus, M.P.; Grant, I.; Translational Methamphetamine AIDS Research Center (TMARC). Prior Methamphetamine Use Disorder History Does Not Impair Interoceptive Processing of Soft Touch in HIV Infection. Viruses 2021, 13, 2476. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122476
Bischoff-Grethe A, Ellis RJ, Tapert SF, Paulus MP, Grant I, Translational Methamphetamine AIDS Research Center (TMARC). Prior Methamphetamine Use Disorder History Does Not Impair Interoceptive Processing of Soft Touch in HIV Infection. Viruses. 2021; 13(12):2476. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122476
Chicago/Turabian StyleBischoff-Grethe, Amanda, Ronald J. Ellis, Susan F. Tapert, Martin P. Paulus, Igor Grant, and Translational Methamphetamine AIDS Research Center (TMARC). 2021. "Prior Methamphetamine Use Disorder History Does Not Impair Interoceptive Processing of Soft Touch in HIV Infection" Viruses 13, no. 12: 2476. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122476