
SARS-CoV-2 Variant of Concern 202012/01 Has about Twofold Replicative Advantage and Acquires Concerning Mutations

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Genome Sequence Analysis
2.3. Monte Carlo Estimation of the Confidence Interval for the Growth Rate of the Ratio of Strains
2.4. Estimation of the Relative Replication Advantage of Viral Strains
3. Results
3.1. Evolution of SARS-CoV-2 in England
3.2. Replicative Advantage of the VOC Strain over the 20A.EU1 Strain in England
3.3. Worldwide Spread of the VOC Strain
3.4. Replicative Advantage of the 501Y.V2 Strain in South Africa
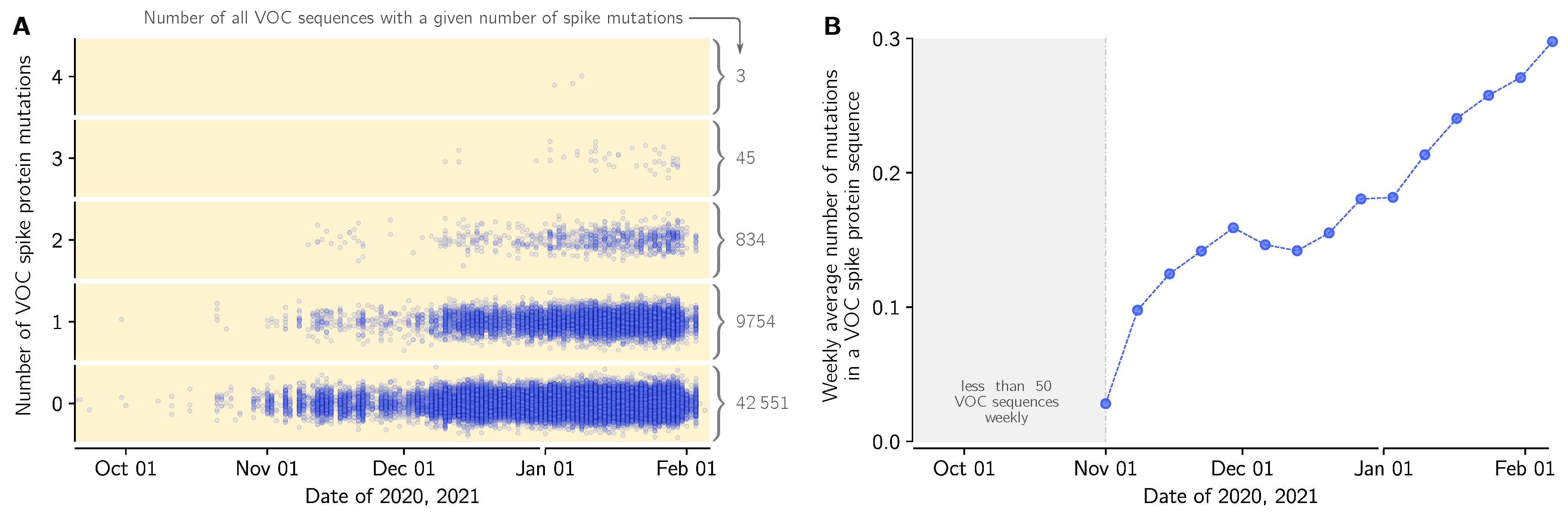
3.5. Emergence of Mutations in VOC Genomes in England
3.6. Growth of the L18F Substrain
3.7. VOC Strain Mutations in Spike Receptor-Binding Domain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rambaut, A.; Loman, N.; Pybus, O. Preliminary Genomic Characterisation of an Emergent SARS-CoV-2 Lineage in the UK Defined by a Novel Set of Spike Mutations. Available online: https://virological.org/t/preliminary-genomic-characterisation-of-an-emergent-sars-cov-2-lineage-in-the-uk-defined-by-a-novel-set-of-spike-mutations/563 (accessed on 9 January 2021).
- Shu, Y.; McCauley, J. GISAID: Global initiative on sharing all influenza data—From vision to reality. Eurosurveillance 2017, 22, 30494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Public Health England. Investigation of Novel SARS-CoV-2 Variant: 202012/01. Technical Briefing 2. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/949639/Technical_Briefing_VOC202012-2_Briefing_2_FINAL.pdf (accessed on 9 December 2020).
- Greater London Authority. Coronavirus (COVID-19) Cases. Available online: https://data.london.gov.uk/dataset/coronavirus--covid-19--cases (accessed on 9 January 2021).
- Hodcroft, E.B.; Zuber, M.; Nadeau, S.; Crawford, K.H.D.; Bloom, J.D.; Veesler, D.; Vaughan, T.G.; Comas, I.; Candelas, F.G.; Stadler, T.; et al. Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020. MedRxiv 2020. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; Wang, X.; Pascal, K.E.; Tomkins-Tinch, C.; Nyalile, T.P.; Wang, Y.; Baum, A.; Diehl, W.E.; Dauphin, A.; Carbone, C.; et al. Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 2020, 183, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Emergence and rapid spread of a new severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) lineage with multiple spike mutations in South Africa. medRxiv 2020. [Google Scholar] [CrossRef]
- Sabino, E.C.; Buss, L.F.; Carvalho, M.P.S.; Crispim, M.A.E.; Fraiji, N.A.; Pereira, R.H.M.; Parag, K.V.; da Silva Peixoto, P.; Kraemer, M.U.G.; Oikawa, M.K.; et al. Resurgence of COVID-19 in Manaus, Brazil, despite high seroprevalence. Lancet 2021, 397, 452–455. [Google Scholar] [CrossRef]
- Public Health England. Investigation of Novel SARS-CoV-2 Variant: 202012/01. Technical Briefing 1. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/947048/Technical_Briefing_VOC_SH_NJL2_SH2.pdf (accessed on 9 December 2020).
- Leung, K.; Shum, M.H.; Leung, G.M.; Lam, T.T.; Wu, J.T. Early transmissibility assessment of the N501Y mutant strains of SARS-CoV-2 in the United Kingdom, October to November 2020. Eurosurveillance 2021, 26, 2002106. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and severity of novel SARS-CoV-2 Variant of Concern 202012/01 in England. medRxiv 2021. [Google Scholar] [CrossRef]
- COVID-19 Genomics UK Consortium. Available online: https://www.cogconsortium.uk (accessed on 12 February 2021).
- Grabowski, F.; Preibisch, G.; Kochańczyk, M.; Lipniacki, T. SARS-CoV-2 Variant Under Investigation 202012/01 has more than twofold replicative advantage. MedRxiv 2021. [Google Scholar] [CrossRef]
- Wibmer, C.K.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Lambson, B.E.; Vermeulen, M.; van den Berg, K.; Rossouw, T.; Boswell, M.; et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. BioRxiv 2021. [Google Scholar] [CrossRef]
- McCallum, M.; Marco, A.D.; Lempp, F.; Tortorici, M.A.; Pinto, D.; Walls, A.C.; Beltramello, M.; Chen, A.; Liu, Z.; Zatta, F.; et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. BioRxiv 2021. [Google Scholar] [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Kochańczyk, M.; Grabowski, F.; Lipniacki, T. Super-spreading events initiated the exponential growth phase of COVID-19 with R0 higher than initially estimated. R. Soc. Open Sci. 2020, 7, 200786. [Google Scholar] [CrossRef]
- De Maio, N.; Borges, R.; Walker, C.; Weilguny, L.; Slodkowicz, G.; Goldman, N. Issues with SARS-CoV-2 Sequencing Data. Available online: https://virological.org/t/issues-with-sars-cov-2-sequencing-data (accessed on 12 February 2021).
- Qiao, B.; Olvera de la Cruz, M. Enhanced Binding of SARS-CoV-2 Spike Protein to Receptor by Distal Polybasic Cleavage Sites. ACS Nano 2020, 14, 10616–10623. [Google Scholar] [CrossRef]
- Buss, L.F.; Prete, C.A.J.; Abrahim, C.M.M.; Mendrone, A.J.; Salomon, T.; de Almeida-Neto, C.; França, R.F.O.; Belotti, M.C.; Carvalho, M.P.S.S.; Costa, A.G.; et al. Three-quarters attack rate of SARS-CoV-2 in the Brazilian Amazon during a largely unmitigated epidemic. Science 2021, 371, 288–292. [Google Scholar] [CrossRef]
- Chakraborty, S. Evolutionary and structural analysis elucidates mutations on SARS-CoV2 spike protein with altered human ACE2 binding affinity. Biochem. Biophys. Res. Commun. 2021, 534, 374–380. [Google Scholar] [CrossRef]
- Koenig, P.A.; Das, H.; Liu, H.; Kümmerer, B.M.; Gohr, F.N.; Jenster, L.M.; Schiffelers, L.D.J.; Tesfamariam, Y.M.; Uchima, M.; Wuerth, J.D.; et al. Structure-guided multivalent nanobodies block SARS-CoV-2 infection and suppress mutational escape. Science 2021, 371, eabe6230. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Flegg, J.A.; White, L.J.; Aguas, R. Levels of SARS-CoV-2 population exposure are considerably higher than suggested by seroprevalence surveys. MedRxiv 2021. [Google Scholar] [CrossRef]
- Fratev, F. The N501Y and K417N mutations in the spike protein of SARS-CoV-2 alter the interactions with both hACE2 and human derived antibody: A Free energy of perturbation study. BioRxiv 2020. [Google Scholar] [CrossRef]
- Collier, D.A.; Marco, A.D.; Ferreira, I.A.; Meng, B.; Datir, R.; Walls, A.C.; Bassi, J.; Pinto, D.; Fregni, C.S.; Bianchi, S.; et al. SARS-CoV-2 B.1.1.7 escape from mRNA vaccine-elicited neutralizing antibodies. MedRxiv 2021. [Google Scholar] [CrossRef]
- Kemp, S.A.; Collier, D.; Datir, R.; Ferreira, I.; Gayed, S.; Jahun, A.; Hosmillo, M.; Rees-Spear, C.; Mlcochova, P.; Lumb, I.U.; et al. Neutralising antibodies in Spike mediated SARS-CoV-2 adaptation. MedRxiv 2020. [Google Scholar] [CrossRef]
- Voss, W.N.; Hou, Y.J.; Johnson, N.V.; Kim, J.E.; Delidakis, G.; Horton, A.P.; Bartzoka, F.; Paresi, C.J.; Tanno, Y.; Abbasi, S.A.; et al. Prevalent, protective, and convergent IgG recognition of SARS-CoV-2 non-RBD spike epitopes in COVID-19 convalescent plasma. BioRxiv 2020. [Google Scholar] [CrossRef]
- Cele, S.; Gazy, I.; Jackson, L.; Hwa, S.H.; Tegally, H.; Lustig, G.; Giandhari, J.; Pillay, S.; Wilkinson, E.; Naidoo, Y.; et al. Escape of SARS-CoV-2 501Y.V2 variants from neutralization by convalescent plasma. MedRxiv 2021. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Proportion of VOC Genomes | 95% CrI | |
|---|---|---|---|
| England | 86.1% | (43,260/50,240) | 85.8–86.4% |
| Scotland | 71.3% | (1445/2028) | 69.3–73.2% |
| Ghana | 68.9% | (62/90) | 58.9–77.8% |
| Turkey | 68.6% | (155/226) | 62.4–74.3% |
| Slovakia | 67.7% | (42/62) | 56.5–79.0% |
| Ireland | 62.0% | (364/587) | 58.1–65.9% |
| Italy | 61.0% | (169/277) | 55.2–66.8% |
| Nigeria | 60.3% | (70/116) | 51.7–69.0% |
| N. Ireland | 57.9% | (183/316) | 52.5–63.3% |
| Wales | 53.5% | (2125/3971) | 52.0–55.1% |
| Romania | 52.0% | (26/50) | 38.0–66.0% |
| Spain | 49.1% | (576/1174) | 46.2–52.0% |
| France | 44.7% | (691/1545) | 42.3–47.2% |
| Israel | 36.9% | (113/306) | 31.7–42.5% |
| Australia | 34.4% | (66/192) | 27.6–41.2% |
| Austria | 34.2% | (178/520) | 30.2–38.3% |
| Belgium | 32.6% | (538/1651) | 30.4–34.9% |
| Czechia | 32.1% | (17/53) | 20.8–45.3% |
| Finland | 31.8% | (67/211) | 25.6–37.9% |
| Sweden | 27.7% | (109/394) | 23.4–32.2% |
| Netherlands | 21.0% | (328/1561) | 19.0–23.1% |
| Singapore | 20.6% | (30/146) | 14.4–27.4% |
| Portugal | 20.0% | (127/636) | 17.0–23.1% |
| Germany | 16.9% | (68/403) | 13.4–20.6% |
| Sri Lanka | 14.8% | (13/88) | 8.0–22.7% |
| Norway | 14.3% | (27/189) | 9.5–19.6% |
| S. Korea | 13.3% | (12/90) | 6.7–21.1% |
| India | 12.9% | (11/85) | 5.9–20.0% |
| Luxembourg | 10.7% | (27/253) | 7.1–14.6% |
| Denmark | 10.5% | (1270/12,137) | 9.9–11.0% |
| Brazil | 9.5% | (13/137) | 5.1–14.6% |
| N. Macedonia | 9.3% | (5/54) | 1.9–18.5% |
| Switzerland | 8.2% | (409/4985) | 7.4–9.0% |
| Iceland | 7.7% | (5/65) | 1.5–15.4% |
| Canada | 7.0% | (36/514) | 4.9–9.3% |
| Poland | 6.7% | (11/165) | 3.0–10.9% |
| Mexico | 6.5% | (5/77) | 1.3–13.0% |
| Latvia | 5.7% | (5/87) | 1.2–11.5% |
| USA | 3.7% | (608/16,439) | 3.4–4.0% |
| S. Africa | 0.9% | (1/112) | 0.0–2.7% |
| Colombia | 0.00% | (0/64) | |
| Mayotte | 0.00% | (0/224) | |
| Lithuania | 0.00% | (0/381) | |
| Egypt | 0.00% | (0/84) | |
| Kenya | 0.00% | (0/75) | |
| French Guiana | 0.00% | (0/98) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grabowski, F.; Preibisch, G.; Giziński, S.; Kochańczyk, M.; Lipniacki, T. SARS-CoV-2 Variant of Concern 202012/01 Has about Twofold Replicative Advantage and Acquires Concerning Mutations. Viruses 2021, 13, 392. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030392
Grabowski F, Preibisch G, Giziński S, Kochańczyk M, Lipniacki T. SARS-CoV-2 Variant of Concern 202012/01 Has about Twofold Replicative Advantage and Acquires Concerning Mutations. Viruses. 2021; 13(3):392. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030392
Chicago/Turabian StyleGrabowski, Frederic, Grzegorz Preibisch, Stanisław Giziński, Marek Kochańczyk, and Tomasz Lipniacki. 2021. "SARS-CoV-2 Variant of Concern 202012/01 Has about Twofold Replicative Advantage and Acquires Concerning Mutations" Viruses 13, no. 3: 392. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030392