
Epidemiology and Genomic Analysis of Equine Encephalosis Virus Detected in Horses with Clinical Signs in South Africa, 2010–2017

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. RNA Extraction and Real-Time Reverse Transcription-PCR (rRT-PCR)
2.3. Virus Isolation and Sequencing
2.4. Statistical Analyses
3. Results
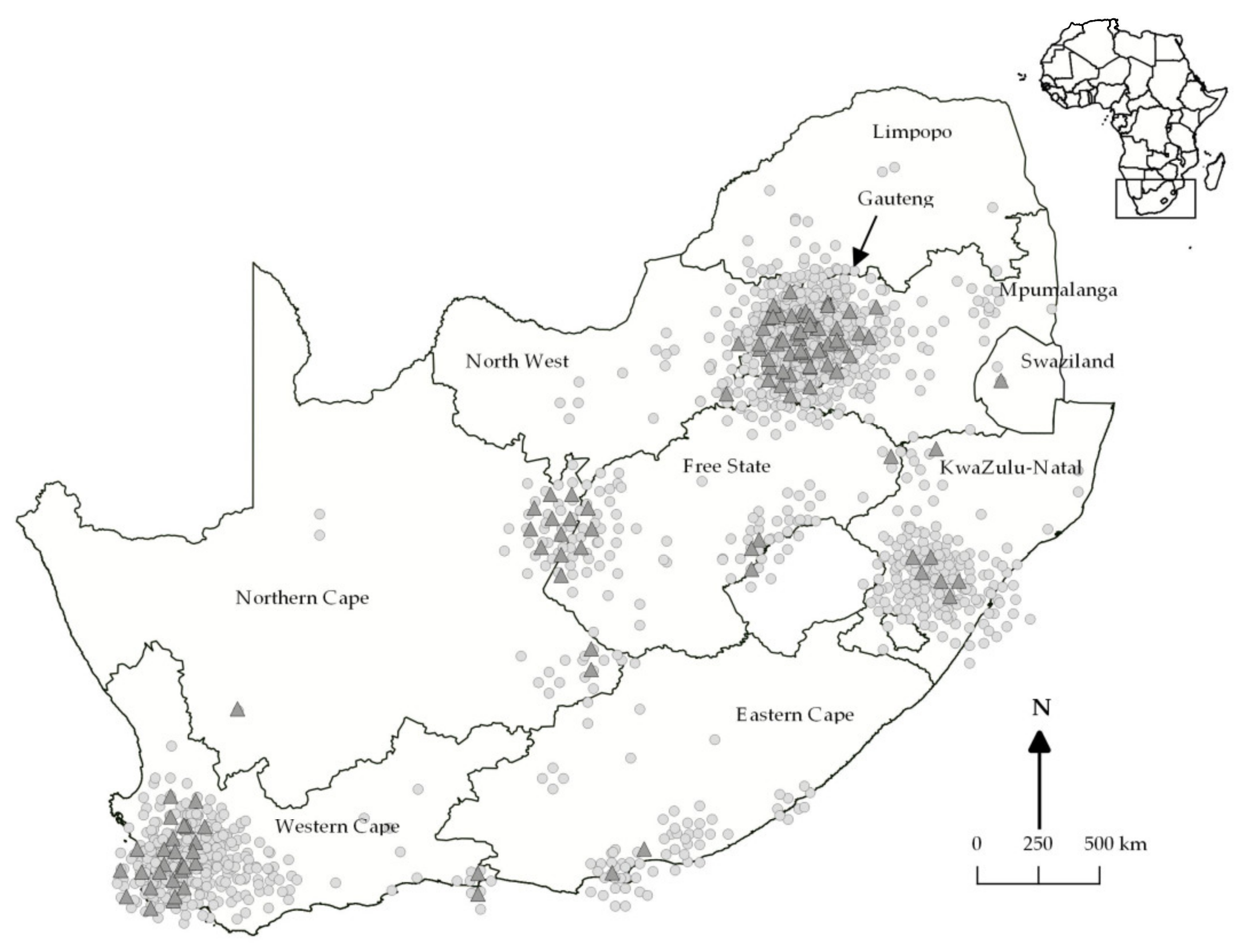
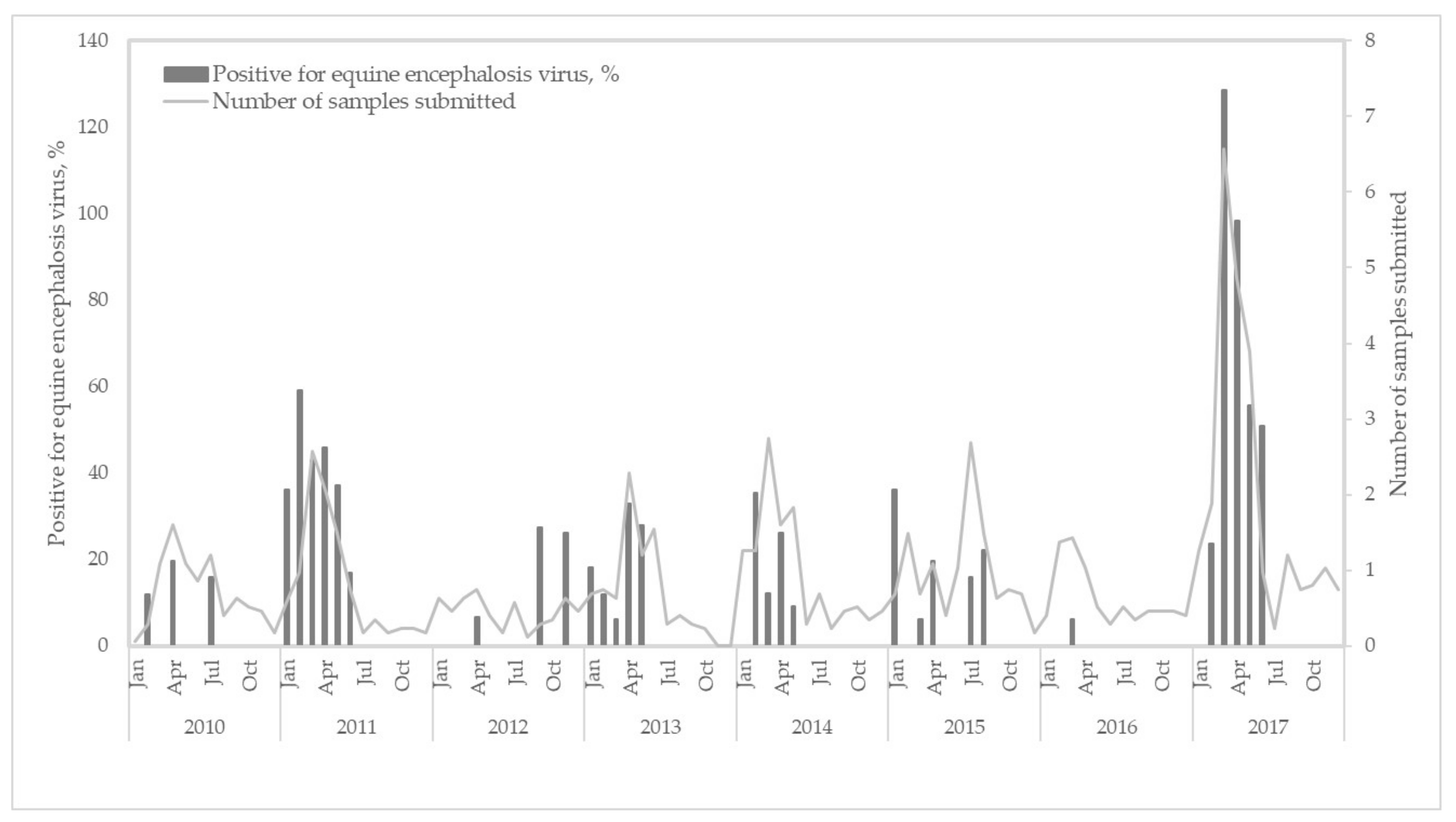
3.1. Epidemiological Analyses of Clinical Cases
3.2. Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erasmus, B.J.; Adelaar, T.F.; Smit, J.D.; Lecatsas, G.; Toms, T. The isolation and characterization of equine encephalosis virus. Bull. Off. Int. Epizoot. 1970, 74, 781–789. [Google Scholar]
- Mildenberg, Z.; Westcott, D.; Bellaiche, M.; Dastjerdi, A.; Steinbach, F.; Drew, T. Equine Encephalosis Virus in Israel. Transbound. Emerg. Dis. 2009, 56, 291. [Google Scholar] [CrossRef] [PubMed]
- Aharonson-Raz, K.; Steinman, A.; Bumbarov, V.; Maan, S.; Maan, N.; Nomikou, K.; Batten, C.; Potgieter, C.; Gottlieb, Y.; Mertens, P.; et al. Isolation and Phylogenetic Grouping of Equine Encephalosis Virus in Israel. Emerg. Infect. Dis. 2011, 17, 1883–1886. [Google Scholar] [CrossRef]
- Yadav, P.D.; Albariño, C.D.; Nyayanit, D.A.; Guerrero, L.; Jenks, M.H.; Sarkale, P.; Nichol, S.T.; Mourya, D.T. Equine Encephalosis Virus in India, 2008. Emerg. Infect. Dis. 2018, 24, 898–901. [Google Scholar] [CrossRef]
- Howell, P.G.; Guthrie, A.J.; Coetzer, J.A.W. Equine encephalosis. In Infectious Diseases of Livestock; Coetzer, J.A.W., Tustin, R.C., Eds.; Oxford University Press: Cape Town, South Africa, 2004; pp. 1247–1251. [Google Scholar]
- Viljoen, G.J.; Huismans, H. The Characterization of Equine Encephalosis Virus and the Development of Genomic Probes. J. Gen. Virol. 1989, 70, 2007–2015. [Google Scholar] [CrossRef]
- Thompson, G.M.; Jess, S.; Murchie, A.K. A review of African horse sickness and its implications for Ireland. Ir. Vet. J. 2012, 65, 9. [Google Scholar] [CrossRef] [Green Version]
- Roy, P. Orbiviruses and their replication. In Fields’ Virology. 5; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA; New York, NY, USA, 2007; pp. 1975–1997. [Google Scholar]
- Zientara, S.; Lecollinet, S. African horse sickness. Rev. Sci. Tech. 2015, 34, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Saegerman, C.; Berkvens, D.; Mellor, P.S. Bluetongue Epidemiology in the European Union. Emerg. Infect. Dis. 2008, 14, 539–544. [Google Scholar] [CrossRef]
- Carpenter, S.; Mellor, P.S.; Fall, A.G.; Garros, C.; Venter, G.J. African Horse Sickness Virus: History, Transmission, and Current Status. Annu. Rev. Èntomol. 2017, 62, 343–358. [Google Scholar] [CrossRef]
- Lu, G.; Pan, J.; Ou, J.; Shao, R.; Hu, X.; Wang, C.; Li, S. African horse sickness: Its emergence in Thailand and potential threat to other Asian countries. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Theodoridis, A.; Nevill, E.M.; Els, H.J.; Boshoff, S.T. Viruses isolated from Culicoides midges in South Africa during unsuc-cessful attempts to isolate bovine ephemeral fever virus. Onderstepoort J. Vet. Res. 1979, 46, 191–198. [Google Scholar]
- Venter, G.J.; Groenewald, D.M.; Paweska, J.T.; Venter, E.H.; Howell, P.G. Vector competence of selected South African Culi-coides species for the Bryanston serotype of equine encephalosis virus. Med. Vet. Entomol. 1999, 13, 393–400. [Google Scholar] [CrossRef]
- Paweska, J.T.; Venter, G.J. Vector competence of Culicoides species and the seroprevalence of homologous neutralizing anti-body in horses for six serotypes of equine encephalosis virus (EEV) in South Africa. Med. Vet. Entomol. 2004, 18, 398–407. [Google Scholar] [CrossRef]
- Nomikou, K.; Hughes, J.; Wash, R.; Kellam, P.; Breard, E.; Zientara, S.; Palmarini, M.; Biek, R.; Mertens, P.P.C. Widespread Reassortment Shapes the Evolution and Epidemiology of Bluetongue Virus following European Invasion. PLoS Pathog. 2015, 11, e1005056. [Google Scholar] [CrossRef]
- Weyer, C.T.; Grewar, J.D.; Burger, P.; Rossouw, E.; Lourens, C.W.; Joone, C.; Le Grange, M.; Coetzee, P.; Venter, E.H.; Martin, D.P.; et al. African Horse Sickness Caused by Genome Reassortment and Reversion to Virulence of Live, Attenuated Vaccine Viruses, South Africa, 2004–2014. Emerg. Infect. Dis. 2016, 22, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Bremer, C.W.; Huismans, H.; Van Dijk, A.A. Characterization and cloning of the African horse sickness virus genome. J. Gen. Virol. 1990, 71, 793–799. [Google Scholar] [CrossRef]
- Belhouchet, M.; Jaafar, F.M.; Firth, A.E.; Grimes, J.M.; Mertens, P.P.C.; Attoui, H. Detection of a Fourth Orbivirus Non-Structural Protein. PLoS ONE 2011, 6, e25697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verwoerd, D.W.; Els, H.J.; De Villiers, E.-M.; Huismans, H. Structure of the Bluetongue Virus Capsid. J. Virol. 1972, 10, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Roy, P. Bluetongue virus: Dissection of the polymerase complex. J. Gen. Virol. 2008, 89 Pt 8, 1789–1804. [Google Scholar] [CrossRef]
- Dhama, K.; Pawaiya, R.V.S.; Karthik, K.; Chakraborty, S.; Tiwari, R.; Verma, A. Equine encephalosis virus (EEV): A Review. Asian J. Anim. Vet. Adv. 2014, 9, 123–133. [Google Scholar] [CrossRef]
- Nason, E.L.; Rothagel, R.; Mukherjee, S.K.; Kar, A.K.; Forzan, M.; Prasad, B.V.V.; Roy, P. Interactions between the Inner and Outer Capsids of Bluetongue Virus. J. Virol. 2004, 78, 8059–8067. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, A.D.; Zhao, Y.; Roy, P. Release of Bluetongue Virus-like Particles from Insect Cells is Mediated by BTV Nonstructural Protein NS3/NS3A. Virology 1993, 193, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Howell, P.G.; Groenewald, D.; Visage, C.W.; Bosman, A.M.; Coetzer, J.A.W.; Guthrie, A.J. The classification of seven sero-types of equine encephalosis virus and the prevalence of homologous antibody in horses in South Africa. Onderstepoort J. Vet. Res. 2002, 69, 179–193. [Google Scholar]
- Barnard, B.J.H.; Paweska, J.T. Prevalence of antibodies against some equine viruses in zebra (Zebra burchellt) in the Kruger National Park, 1991–1992. Onderstepoort J. Vet. Res. 1993, 60, 175–179. [Google Scholar] [PubMed]
- Nevill, E.M.; Erasmus, B.J.; Venter, G.J. A six-year survey of viruses associated with Culicoides biting midges through-out South Africa (Diptera: Ceratopogonidae). In Bluetongue, African Horse Sickness and Related Orbiviruses, Proceedings of the Second International Symposium, Miami, FL, USA, 15–19 March 1992; Walton, T.E., Osburn, B.I., Eds.; CRC Press: Boca Raton, FL, USA, 1992; pp. 314–319. [Google Scholar]
- Howell, P.G.; Nurton, J.P.; Nel, D.; Lourens, C.W.; Guthrie, A.J. Prevalence of serotype specific antibody to equine encepha-losis virus in Thoroughbred yearlings in South Africa (1999–2004). Onderstepoort J. Vet. Res. 2008, 75, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Paweska, J. A Fatal Case of Equine Encephalosis in the Port Elizabeth District, South Africa. AnimalNet 1999. [Google Scholar]
- Grewar, J.D.; Thompson, P.N.; Lourens, C.W.; Guthrie, A.J. Equine encephalosis in Thoroughbred foals on a South African stud farm. Onderstepoort J. Vet. Res. 2015, 82, 966. [Google Scholar] [CrossRef] [PubMed]
- Wescott, D.G.; Mildenberg, Z.; Bellaiche, M.; McGowan, S.L.; Grierson, S.S.; Choudhury, B.; Steinbach, F. Evidence for the circulation of equine encephalosis virus in Israel since 2001. PLoS ONE 2013, 8, e70532. [Google Scholar] [CrossRef] [Green Version]
- Van Niekerk, M.; Freeman, M.; Paweska, J.T.; Howell, P.G.; Guthrie, A.J.; Potgieter, A.C.; Van Staden, V.; Huismans, H. Variation in the NS3 gene and protein in South African isolates of bluetongue and equine encephalosis viruses. J. Gen. Virol. 2003, 84, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Potgieter, A.C.; Page, N.A.; Liebenberg, J.; Wright, I.M.; Landt, O.; van Dijk, A.A. Improved strategies for se-quence-independent amplification and sequencing of viral double-stranded RNA genomes. J. Gen. Virol. 2009, 90, 1423–1432. [Google Scholar] [CrossRef]
- Sakamoto, K.; Mizukoshi, N.; Apiwatnakorn, B.; Iwata, A.; Tsuchiya, T.; Ueda, S.; Imagawa, H.; Sugiura, T.; Kamada, M.; Fukusho, A. The complete sequences of African horse sickness virus serotype 4 (vaccine strain) RNA segment 2 and 6 which encode outer capsid protein. J. Vet. Med. Sci. 1994, 56, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Vreede, F.T.; Cloete, M.; Napier, G.B.; van Dijk, A.A.; Viljoen, G.J. Sequence-independent amplification and cloning of large dsRNA virus genome segments by poly(dA)-oligonucleotide ligation. J. Virol. Methods 1998, 72, 243–247. [Google Scholar] [CrossRef]
- Van Vuren, P.J.; Wiley, M.; Palacios, G.; Storm, N.; McCulloch, S.D.; Markotter, W.; Birkhead, M.; Kemp, A.; Paweska, J.T. Isolation of a Novel Fusogenic Orthoreovirus from Eucampsipoda africana Bat Flies in South Africa. Viruses 2016, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djikeng, A.; Halpin, R.; Kuzmickas, R.; DePasse, J.; Feldblyum, J.; Sengamalay, N.; Afonso, C.; Zhang, X.; Anderson, N.G.; Ghedin, E.; et al. Viral genome sequencing by random priming methods. BMC Genom. 2008, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Bremer, C.W.; Viljoen, G.J. Detection of African horse sickness virus and discrimination between two equine orbivirus serogroups by reverse transcription polymerase chain reaction. Onderstepoort J. Vet. Res. 1998, 65, 1–8. [Google Scholar]
- Williams, R.; Du Plessis, D.H.; Van Wyngaardt, W. Group-reactive ELISAs for detecting antibodies to African horse sickness and equine encephalosis viruses in horse, donkey, and zebra sera. J. Vet. Diagn. Investig. 1993, 5, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzies, F.D.; McCullough, S.J.; McKeown, I.M.; Forster, J.L.; Jess, S.; Batten, C.; Murchie, A.K.; Gloster, J.; Fallows, J.G.; Pelgrim, W.; et al. Evidence for transplacental and con-tact transmission of bluetongue virus in cattle. Vet. Rec. 2008, 163, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Darpel, K.E.; Batten, C.A.; Veronesi, E.; Williamson, S.; Anderson, P.; Dennison, M.; Clifford, S.; Smith, C.; Philips, L.; Bidewell, C.; et al. Transplacental Transmission of Bluetongue Virus 8 in Cattle, UK. Emerg. Infect. Dis. 2009, 15, 2025–2028. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.; Mellor, P.S.; Szmaragd, C.; Mertens, P.P.C. Adaptive strategies of African horse sickness virus to facilitate vector transmission. Vet. Res. 2008, 40, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLachlan, N.J.; Guthrie, A.J. Re-emergence of bluetongue, African horse sickness, and other Orbivirus diseases. Vet. Res. 2010, 41, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laegrid, W.W. Virus infections of equines. In Equine Encephalosis; Studdert, M.J., Ed.; Elsevier: New York, NY, USA, 1996; pp. 1–125. [Google Scholar]
- Barnard, B.J. Antibodies against some viruses of domestic animals in southern African wild animals. Onderstepoort J. Vet. Res. 1997, 64, 95–110. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Sign | EEV Positive (n = 106; %) | EEV Negative (n = 1360; %) | OR [95.0% Confidence Interval (CI)] | p-Value |
|---|---|---|---|---|
| Pyrexia | 77 (72.6%) | 664 (48.8%) | 2.8 [1.8–4.3] | 0.00 |
| Icterus | 20 (18.8%) | 132 (9.7%) | 2.2 [1.3–3.6] | 0.00 |
| Dyspnea | 12 (11.3%) | 30 (2.2%) | 5.7 [2.8–11.4] | 0.00 |
| Neurological | 51 (48.1%) | 1093 (80.4%) | 0.2 [0.2–0.3] | 0.00 |
| Ataxia | 27 (25.5%) | 535 (39.3.%) | 0.5 [0.3–0.8] | 0.00 |
| Recumbency | 9 (8.5%) | 211 (15.5%) | 0.5 [0.3–1.0] | 0.06 |
| Paralysis | 6 (5.7%) | 127 (9.3%) | 0.6 [0.3–1.4] | 0.30 |
| Paresis | 15 (14.1%) | 189 (13.9%) | 1.0 [0.6–1.8] | 0.90 |
| Depression | 7 (6.6%) | 72 (5.3%) | 1.3 [0.6–2.8] | 0.51 |
| Supra orbital swelling | 2 (1.9%) | 29 (2.1%) | 0.9 [0.2–3.8] | 1.00 |
| Swollen limbs | 1 (0.9%) | 21 (1.5%) | 0.6 [0.1–4.6] | 1.00 |
| Paddling | 0 | 20 (1.5%) | Undefined | 0.39 |
| Outcome | N = 111 | N =1412 | OR [95.0% CI] | P-value |
| SUD | 0 | 9 (0.6%) | Undefined | 1.0 |
| Abortion | 1 (0.9%) | 10 (0.7%) | 1.3 [0.2–10.0] | 0.57 |
| Fatal | 11 (9.9%) | 358 (25.2%) | 0.3 [0.2–0.6] | 0.00 |
| No clinical history | 4 (3.6%) | 33 (2.3%) | 1.5 [0.5–4.3] | 0.4 |
| Sample Code | Month Year | Main Sign Upon Submission | Province | Specimen Isolate Obtained From | Isolate (Vero) Passage Number | VP2 SeroType |
|---|---|---|---|---|---|---|
| SAE055/11 | Mar-11 | Dyspnea | Northern Cape | EDTA | 5 (93) | 4 |
| ZRU088/13/2 | Apr-13 | Fever | Western Cape | EDTA | 5 (95) | 1 |
| ZRU122/13/1 | May-13 | Neurological | Western Cape | EDTA | 5 (95) | 1 |
| ZRU060/15 | Mar-15 | Neurological | Western Cape | EDTA | 5 (95) | 1 |
| ZRU083/15 | Apr-15 | Neurological | Free State | EDTA | 5 (95) | 1 |
| ZRU148/17 | Mar-17 | Abortion | North West | Fetal Placenta | 4 (95) | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snyman, J.; Koekemoer, O.; van Schalkwyk, A.; Jansen van Vuren, P.; Snyman, L.; Williams, J.; Venter, M. Epidemiology and Genomic Analysis of Equine Encephalosis Virus Detected in Horses with Clinical Signs in South Africa, 2010–2017. Viruses 2021, 13, 398. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030398
Snyman J, Koekemoer O, van Schalkwyk A, Jansen van Vuren P, Snyman L, Williams J, Venter M. Epidemiology and Genomic Analysis of Equine Encephalosis Virus Detected in Horses with Clinical Signs in South Africa, 2010–2017. Viruses. 2021; 13(3):398. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030398
Chicago/Turabian StyleSnyman, Jumari, Otto Koekemoer, Antoinette van Schalkwyk, Petrus Jansen van Vuren, Louwtjie Snyman, June Williams, and Marietjie Venter. 2021. "Epidemiology and Genomic Analysis of Equine Encephalosis Virus Detected in Horses with Clinical Signs in South Africa, 2010–2017" Viruses 13, no. 3: 398. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030398