
Effect of Inactivation Methods on SARS-CoV-2 Virion Protein and Structure

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Cells
2.2. Heat Inactivation
2.3. UV Inactivation
2.4. RNA Quantification
2.5. In Vitro Transcribed RNA
2.6. Sucrose Cushion Purification
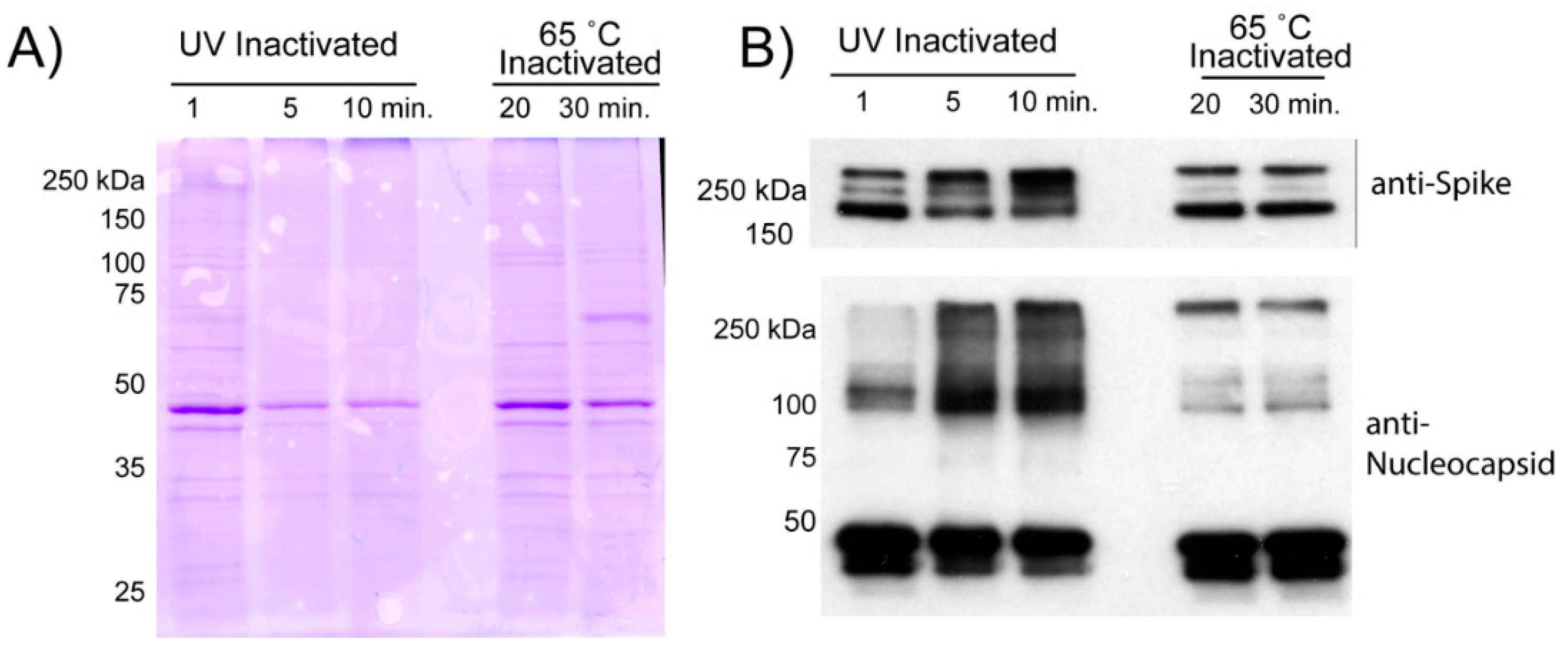
2.7. Gel Electrophoresis and Western Blotting
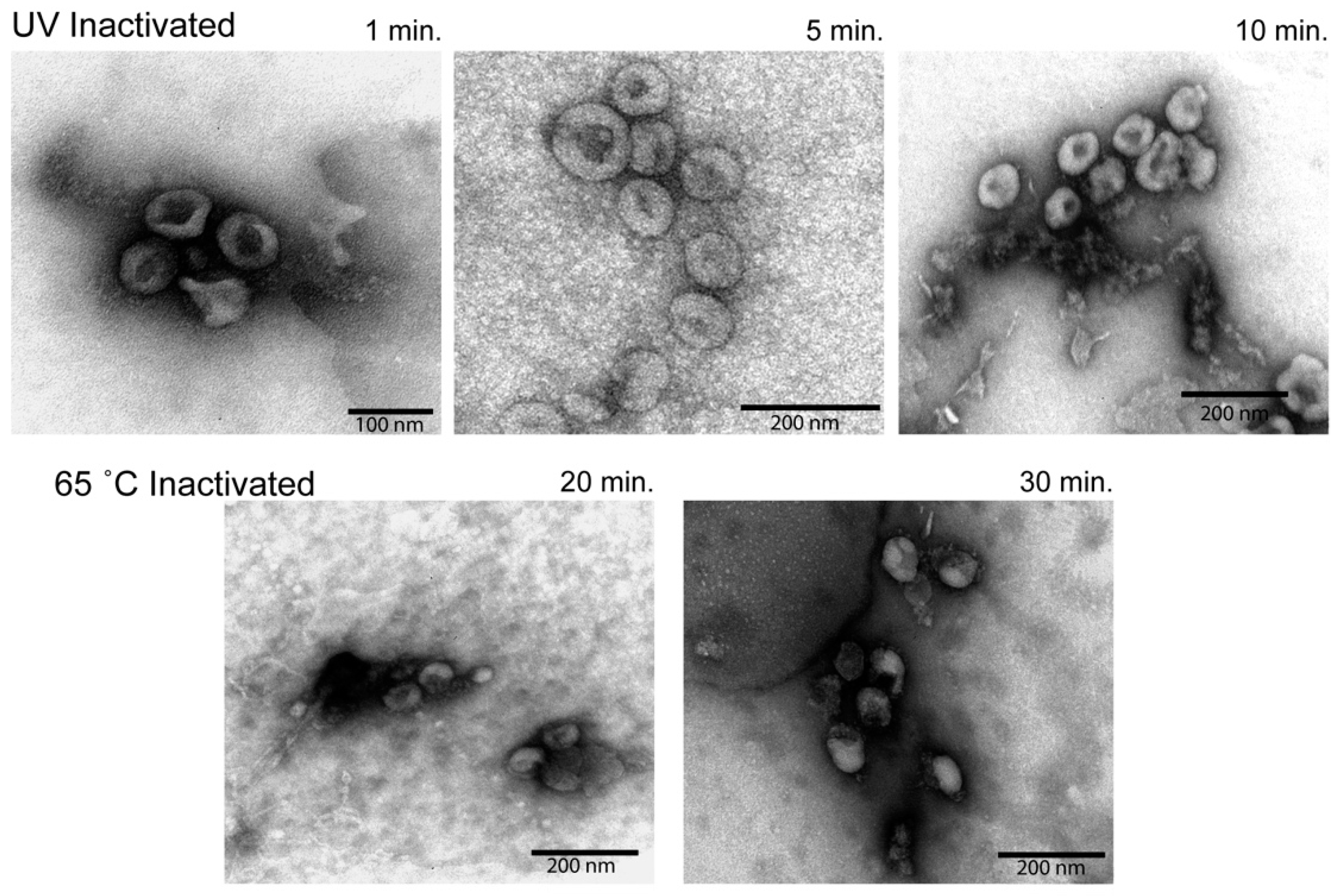
2.8. Electron Microscopy
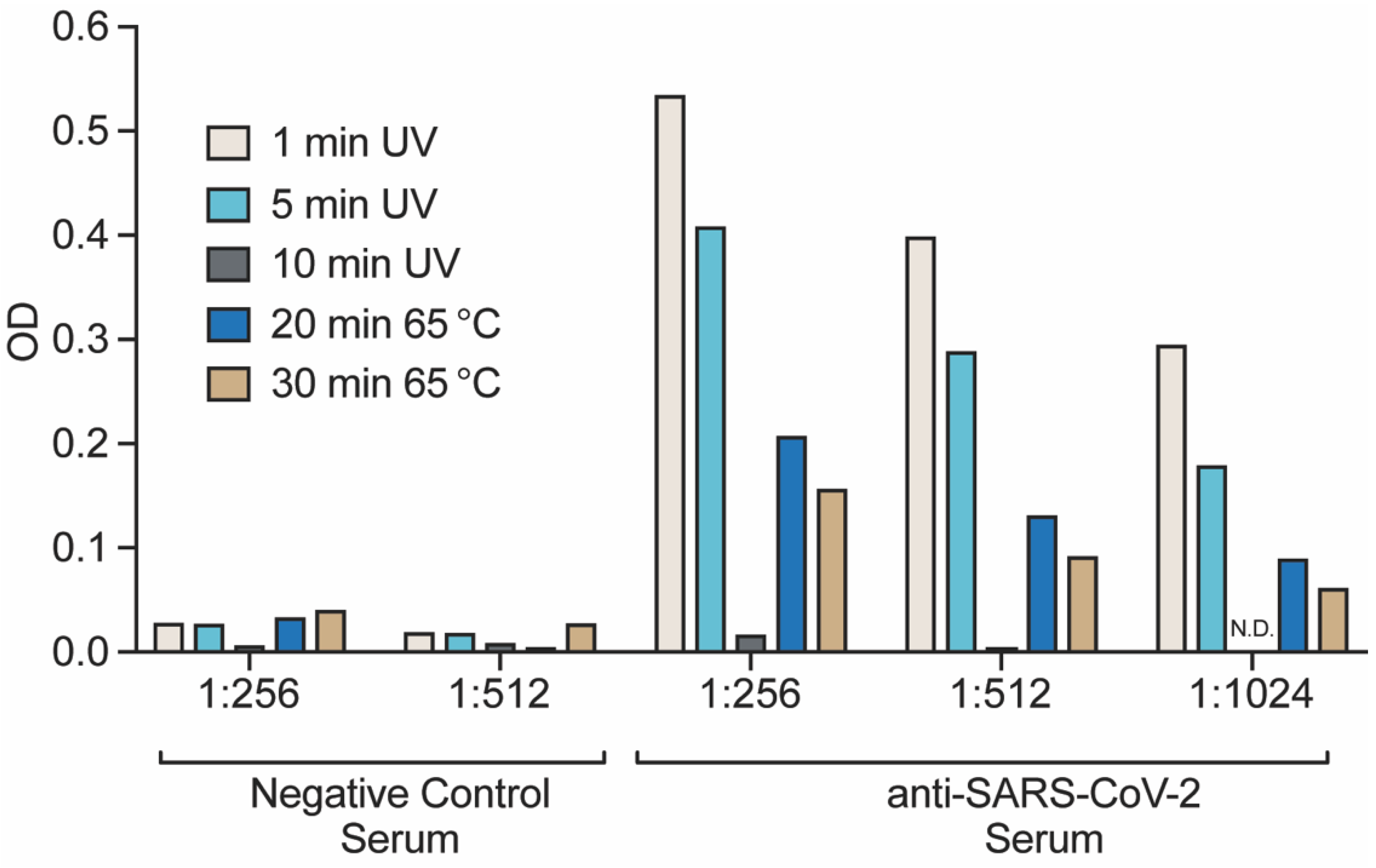
2.9. Detection of Viral Antigens by ELISA
3. Results
3.1. Inactivation of SARS-CoV-2 by Exposure to Elevated Temperature
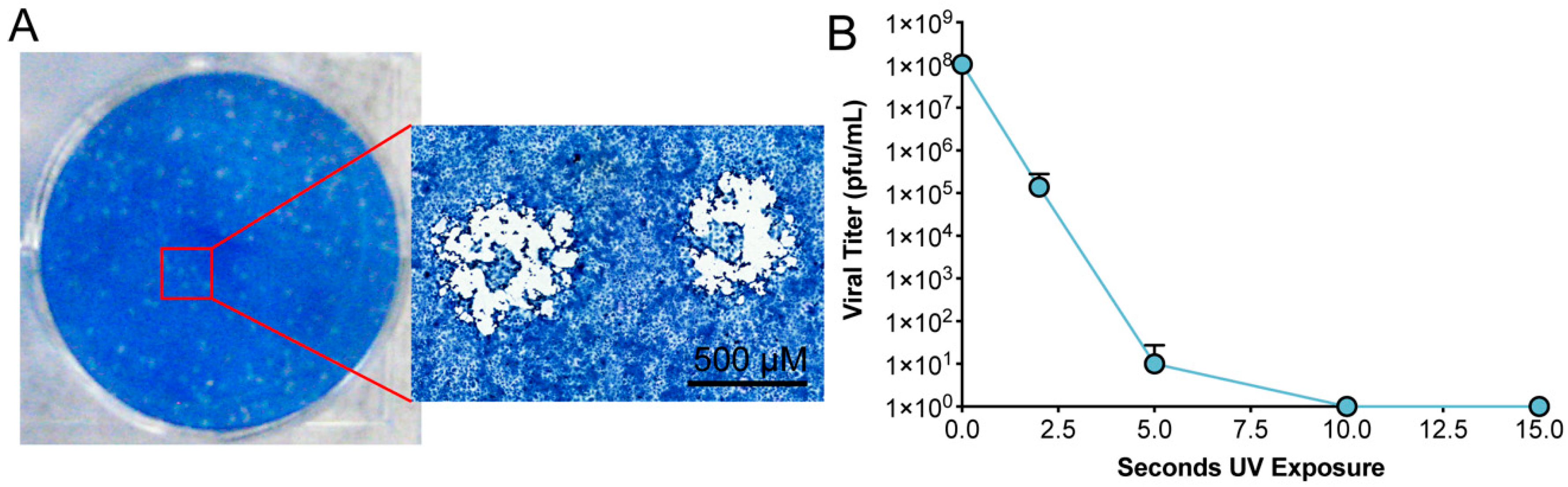
3.2. Inactivation of SARS-CoV-2 by Exposure to UV-C Irradiation
Calculating Sufficient Levels of UV-C Irradiation
3.3. Detection of RNA from Inactivated Supernatants
3.4. Purification of Virions from Inactivated Supernatant
3.5. Electron Microscopy Analysis of Inactivated Virions
3.6. Detection of Inactivated and Purified Virions by ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jureka, A.S.; Silvas, J.A.; Basler, C.F. Propagation, Inactivation, and Safety Testing of SARS-CoV-2. Viruses 2020, 12, 622. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, B.; Touret, F.; Gilles, M.; de Lamballerie, X.; Charrel, R.N. Heat Inactivation of Different Types of SARS-CoV-2 Samples: What Protocols for Biosafety, Molecular Detection and Serological Diagnostics? Viruses 2020, 12, 735. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.P.; Plourde, B.D.; Cheng, L. Using heat to kill SARS-CoV-2. Rev. Med. Virol. 2020, 177, 71. [Google Scholar] [CrossRef] [PubMed]
- Heilingloh, C.S.; Aufderhorst, U.W.; Schipper, L.; Dittmer, U.; Witzke, O.; Yang, D.; Zheng, X.; Sutter, K.; Trilling, M.; Alt, M.; et al. Susceptibility of SARS-CoV-2 to UV Irradiation. Am. J. Infect. Control 2020, 48, 1273–1275. [Google Scholar] [CrossRef] [PubMed]
- Hotchin, J.E. Use of Methyl Cellulose Gel as a Substitute for Agar in Tissue-Culture Overlays. Nature 1955, 175, 352. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of Novel Coronavirus (2019-nCoV) by Real-Time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar]
- Hofmann, M.; Wyler, R. Enzyme-linked immunosorbent assay for the detection of porcine epidemic diarrhea coronavirus antibodies in swine sera. Vet. Microbiol. 1990, 21, 263–273. [Google Scholar] [CrossRef]
- Wu, Z.-G.; Zheng, H.-Y.; Gu, J.; Li, F.; Lv, R.-L.; Deng, Y.-Y.; Xu, W.-Z.; Tong, Y.-Q. Effects of Different Temperature and Time Durations of Virus Inactivation on Results of Real-time Fluorescence PCR Testing of COVID-19 Viruses. Curr. Med. Sci. 2020, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Darnell, M.E.R.; Subbarao, K.; Feinstone, S.M.; Taylor, D.R. Inactivation of the coronavirus that induces severe acute respiratory syndrome, SARS-CoV. J. Virol. Methods 2004, 121, 85–91. [Google Scholar] [CrossRef] [PubMed]
- von Woedtke, A.K.T. The limits of sterility assurance. GMS Krankenh. Interdiszip. 2008, 3, 789–793. [Google Scholar]
- Wang, Y.; Song, W.; Zhao, Z.; Chen, P.; Liu, J.; Li, C. The impacts of viral inactivating methods on quantitative RT-PCR for COVID-19. Virus Res. 2020, 285, 197988. [Google Scholar] [CrossRef] [PubMed]
- Neuman, B.W.; Adair, B.D.; Yoshioka, C.; Quispe, J.D.; Orca, G.; Kuhn, P.; Milligan, R.A.; Yeager, M.; Buchmeier, M.J. Supramolecular Architecture of Severe Acute Respiratory Syndrome Coronavirus Revealed by Electron Cryomicroscopy. J. Virol. 2006, 80, 7918–7928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsunetsugu-Yokota, Y. Large-Scale Preparation of UV-Inactivated SARS Coronavirus Virions for Vaccine Antigen. In SARS and Other Coronaviruses; Laboratory Protocols; Humana Press: Totowa, NJ, USA, 2008; Volume 454, pp. 119–126. [Google Scholar]
- Cerutti, H.; Ricci, V.; Tesi, G.; Soldatini, C.; Castria, M.; Vaccaro, M.N.; Tornesi, S.; Toppi, S.; Verdiani, S.; Brogi, A. Large scale production and characterization of SARS-CoV-2 whole antigen for serological test development. J. Clin. Lab. Anal. 2021, e23735. [Google Scholar] [CrossRef]
- Burton, J.; Love, H.; Richards, K.; Burton, C.; Summers, S.; Pitman, J.; Easterbrook, L.; Davies, K.; Spencer, P.; Killip, M.; et al. The effect of heat-treatment on SARS-CoV-2 viability and detection. J. Virol. Methods 2021, 290, 114087. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Dilution Plated | Plaques Counted (Each Replicate) | Calculated Titer (pfu/mL) |
|---|---|---|---|
| 0 min | 10−5 10−6 | 215 14 | 1.04 × 108 |
| 15 min | undiluted | 0, 0, 0 | 0 |
| 20 min | undiluted | 0, 0, 0 | 0 |
| 25 min | undiluted | 0, 0, 0 | 0 |
| 30 min | undiluted | 0, 0, 0 | 0 |
| Time | Cumulative UV Dose µJ/cm2 | Dilution Plated | Plaques Counted (Each Replicate) | Calculated Titer (pfu/mL) |
|---|---|---|---|---|
| 0 s | 0 | 10−5 10−6 | 215 14 | 1.04 × 108 |
| 2 s | 1.82 × 104 | 10−2 10−3 | TMTC 1, TMTC, 14 23, 57, 0 | 1.39 × 105 |
| 5 s | 3.61 × 104 | undiluted | 6, 0, 0 | 10 |
| 10 s | 6.69 × 104 | undiluted | 0, 0, 0 | 0 |
| 15 s | 9.78 × 104 | undiluted | 0, 0, 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loveday, E.K.; Hain, K.S.; Kochetkova, I.; Hedges, J.F.; Robison, A.; Snyder, D.T.; Brumfield, S.K.; Young, M.J.; Jutila, M.A.; Chang, C.B.; et al. Effect of Inactivation Methods on SARS-CoV-2 Virion Protein and Structure. Viruses 2021, 13, 562. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040562
Loveday EK, Hain KS, Kochetkova I, Hedges JF, Robison A, Snyder DT, Brumfield SK, Young MJ, Jutila MA, Chang CB, et al. Effect of Inactivation Methods on SARS-CoV-2 Virion Protein and Structure. Viruses. 2021; 13(4):562. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040562
Chicago/Turabian StyleLoveday, Emma K., Kyle S. Hain, Irina Kochetkova, Jodi F. Hedges, Amanda Robison, Deann T. Snyder, Susan K. Brumfield, Mark J. Young, Mark A. Jutila, Connie B. Chang, and et al. 2021. "Effect of Inactivation Methods on SARS-CoV-2 Virion Protein and Structure" Viruses 13, no. 4: 562. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040562