
Role of Envelope Glycoprotein Complexes in Cell-Associated Spread of Human Cytomegalovirus

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Viruses
2.3. Generation of Mutant Viruses
2.4. Determination of Viral Growth
2.5. Knockdown of Gene Expression with siRNA
2.6. Detection of Viral Immediate Early Proteins by Indirect Immunofluorescence
2.7. Immunoblotting
2.8. Statistical Analyses
3. Results
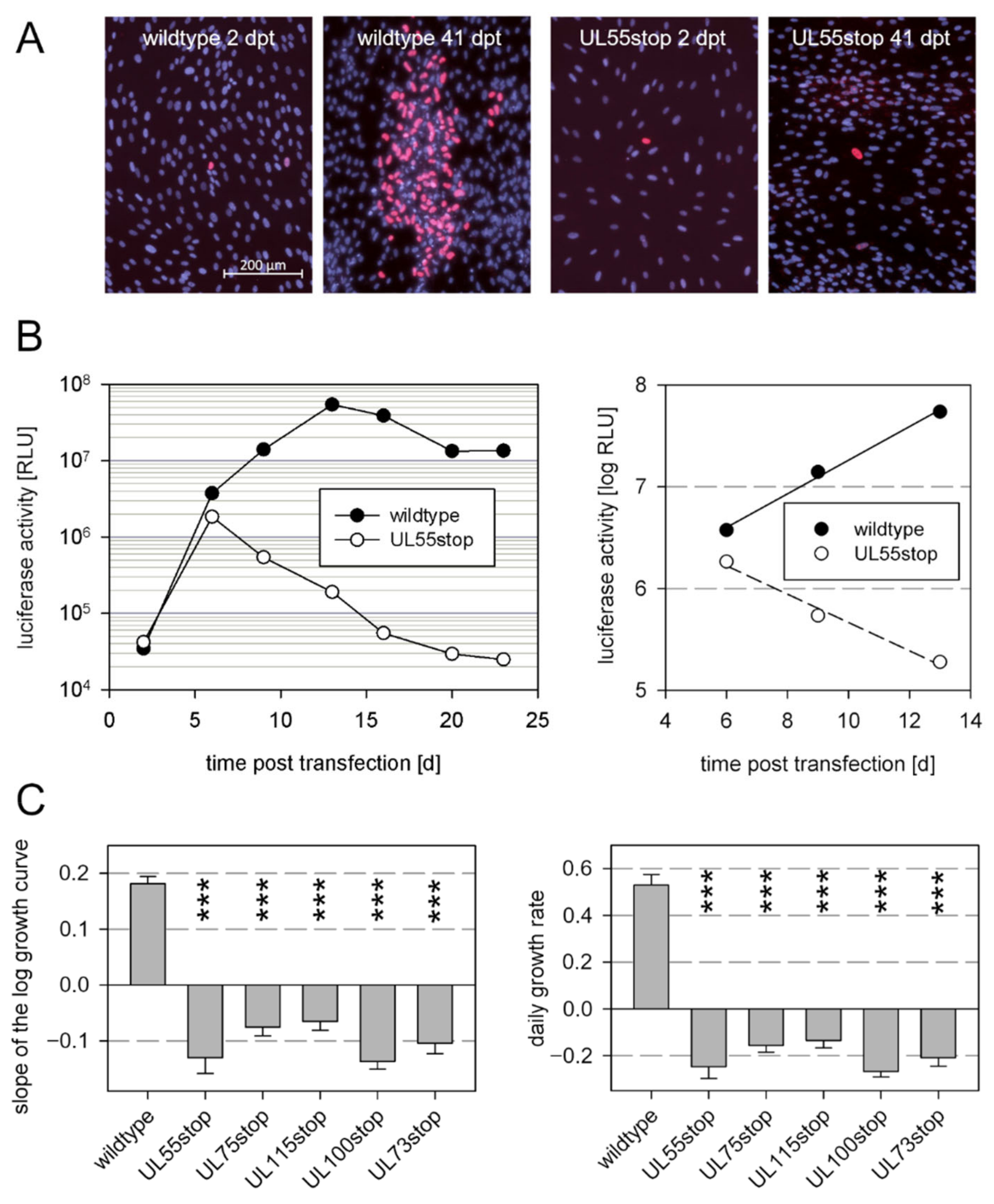
3.1. Envelope Glycoproteins That Are Essential for Cell-Free Virus Spread Are Also Essential for Cell-Associated Growth
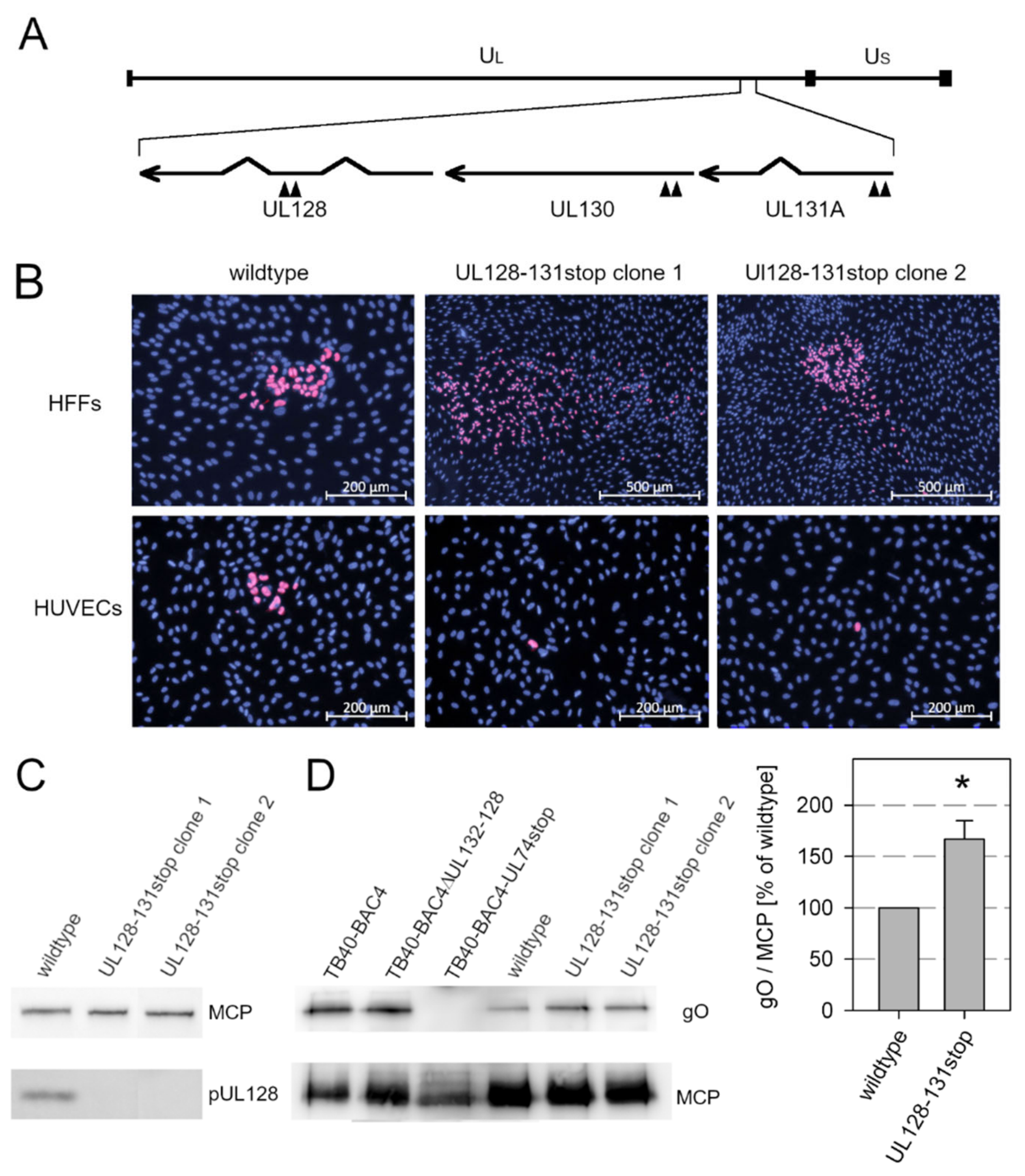
3.2. Knockout of the Pentamer Has Only Limited Effect on Expression of the Trimer in the Background of Strain Merlin
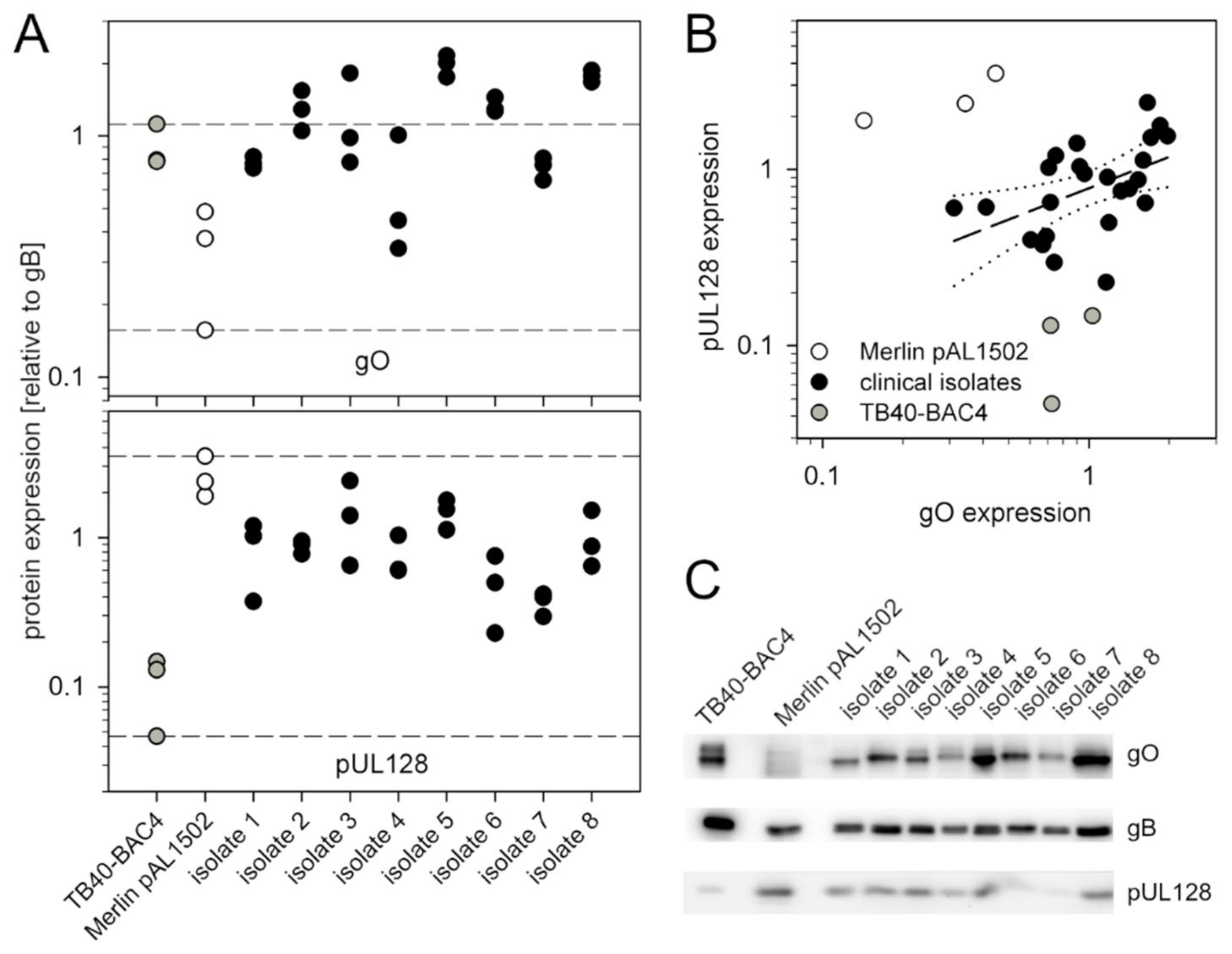
3.3. Recent Isolates of HCMV Express Higher Levels of gO Than the Cell-Associated Model Virus Merlin pAL1502
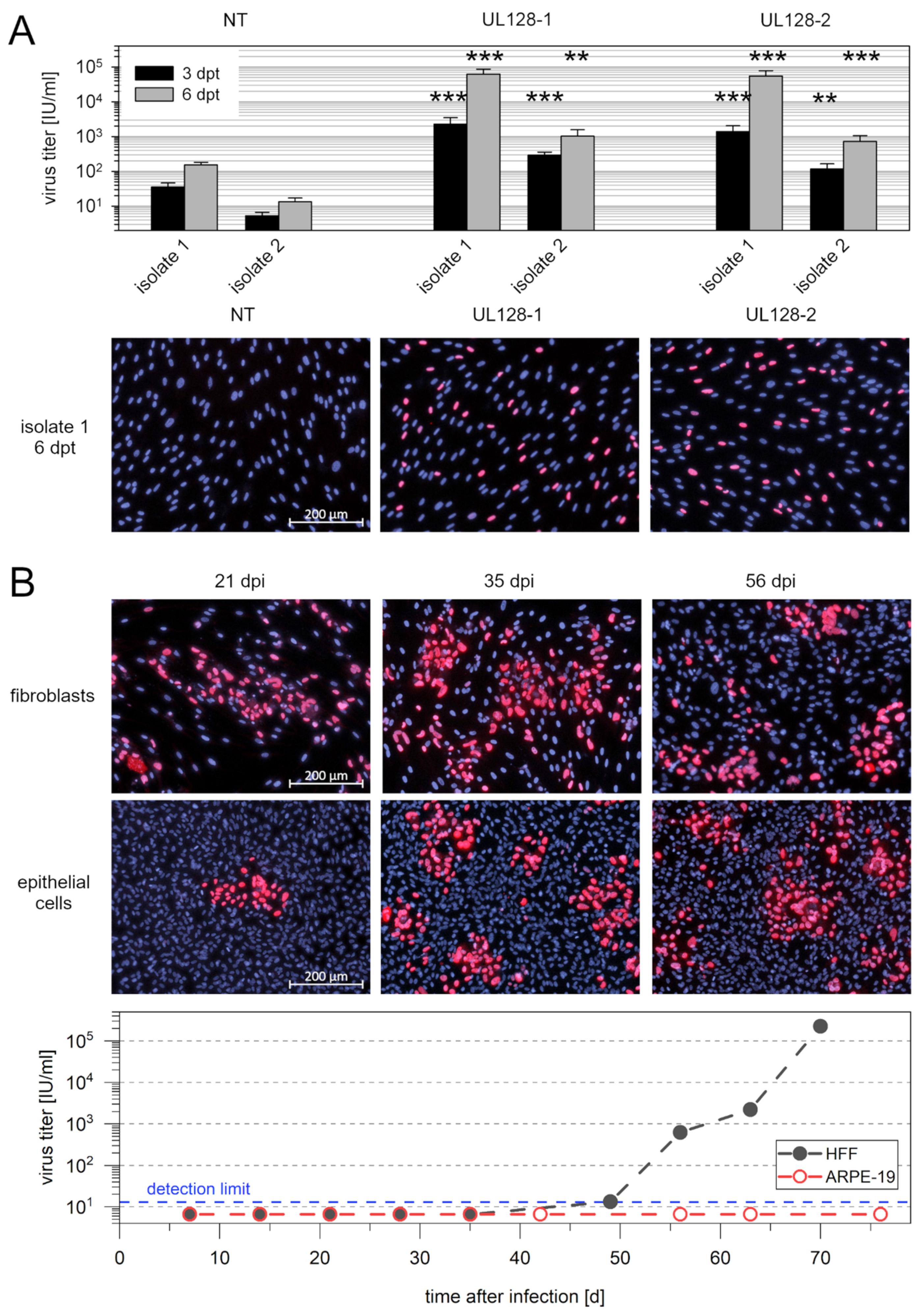
3.4. Spread of Recent Isolates in Fibroblast Culture Is Promoted by gO and Restricted by pUL128
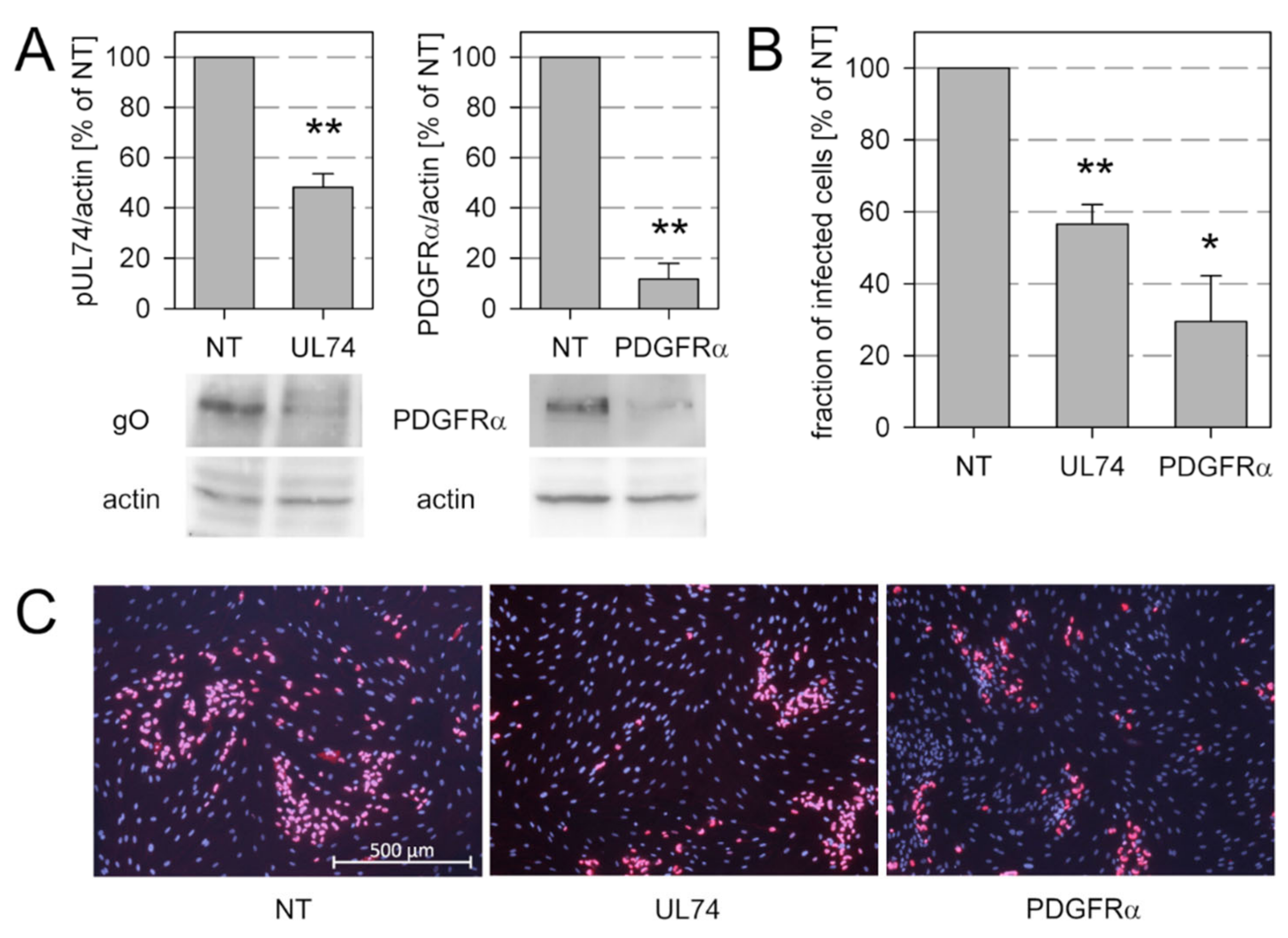
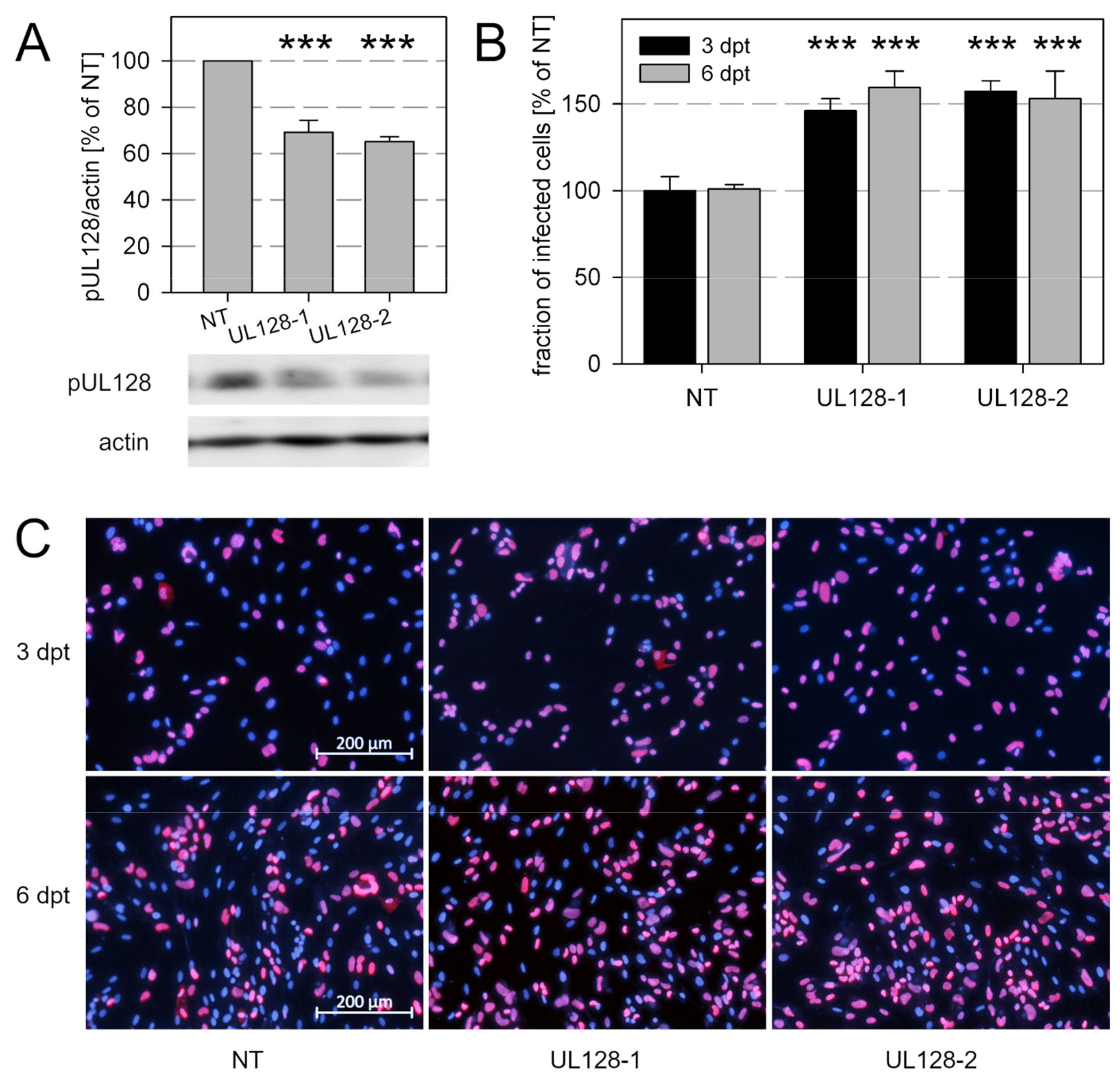
3.5. Knockdown of pUL128 Switches HCMV Isolates Transiently to the Cell-Free Transmission Mode
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of Cytomegalovirus Seroprevalence and Demographic Characteristics Associated with Infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Halenius, A.; Gerke, C.; Hengel, H. Classical and Non-Classical MHC I Molecule Manipulation by Human Cytomegalovirus: So Many Targets—but How Many Arrows in the Quiver? Cell Mol. Immunol 2015, 12, 139–153. [Google Scholar] [CrossRef] [Green Version]
- Falk, J.J.; Winkelmann, M.; Laib Sampaio, K.; Paal, C.; Schrezenmeier, H.; Alt, M.; Stanton, R.; Krawczyk, A.; Lotfi, R.; Sinzger, C. Large-Scale Screening of HCMV-Seropositive Blood Donors Indicates That HCMV Effectively Escapes from Antibodies by Cell-Associated Spread. Viruses 2018, 10, 500. [Google Scholar] [CrossRef] [Green Version]
- Gardner, T.J.; Tortorella, D. Virion Glycoprotein-Mediated Immune Evasion by Human Cytomegalovirus: A Sticky Virus Makes a Slick Getaway. Microbiol. Mol. Biol. Rev. 2016, 80, 663–677. [Google Scholar] [CrossRef] [Green Version]
- Sinzger, C.; Mangin, M.; Weinstock, C.; Topp, M.S.; Hebart, H.; Einsele, H.; Jahn, G. Effect of Serum and CTL on Focal Growth of Human Cytomegalovirus. J. Clin. Virol. 2007, 38, 112–119. [Google Scholar] [CrossRef]
- Murrell, I.; Bedford, C.; Ladell, K.; Miners, K.L.; Price, D.A.; Tomasec, P.; Wilkinson, G.W.G.; Stanton, R.J. The Pentameric Complex Drives Immunologically Covert Cell-Cell Transmission of Wild-Type Human Cytomegalovirus. Proc. Natl. Acad. Sci. USA 2017, 114, 6104–6109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, M.J.; Hyde, T.B.; Schmid, D.S. Review of Cytomegalovirus Shedding in Bodily Fluids and Relevance to Congenital Cytomegalovirus Infection. Rev. Med. Virol. 2011, 21, 240–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemmermann, N.A.W.; Krmpotic, A.; Podlech, J.; Brizic, I.; Prager, A.; Adler, H.; Karbach, A.; Wu, Y.; Jonjic, S.; Reddehase, M.J.; et al. Non-Redundant and Redundant Roles of Cytomegalovirus GH/GL Complexes in Host Organ Entry and Intra-Tissue Spread. PLoS Pathog. 2015, 11, e1004640. [Google Scholar] [CrossRef]
- Podlech, J.; Reddehase, M.J.; Adler, B.; Lemmermann, N.A.W. Principles for Studying in Vivo Attenuation of Virus Mutants: Defining the Role of the Cytomegalovirus GH/GL/GO Complex as a Paradigm. Med. Microbiol. Immunol. 2015, 204, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Bowden, R.A.; Slichter, S.J.; Sayers, M.; Weisdorf, D.; Cays, M.; Schoch, G.; Banaji, M.; Haake, R.; Welk, K.; Fisher, L.; et al. A Comparison of Filtered Leukocyte-Reduced and Cytomegalovirus (CMV) Seronegative Blood Products for the Prevention of Transfusion-Associated CMV Infection after Marrow Transplant. Blood 1995, 86, 3598–3603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, G.L.; Hayes, K.; Hudson, I.L.; James, J. Prevention of Transfusion-Acquired Cytomegalovirus Infection in Infants by Blood Filtration to Remove Leucocytes. Neonatal Cytomegalovirus Infection Study Group. Lancet 1989, 1, 1228–1231. [Google Scholar] [CrossRef]
- Lipson, S.M.; Shepp, D.H.; Match, M.E.; Axelrod, F.B.; Whitbread, J.A. Cytomegalovirus Infectivity in Whole Blood Following Leukocyte Reduction by Filtration. Am. J. Clin. Pathol. 2001, 116, 52–55. [Google Scholar] [CrossRef] [Green Version]
- Rinaldo, C.R.; Black, P.H.; Hirsch, M.S. Interaction of Cytomegalovirus with Leukocytes from Patients with Mononucleosis Due to Cytomegalovirus. J. Infect. Dis. 1977, 136, 667–678. [Google Scholar] [CrossRef]
- Boom, R.; Sol, C.J.A.; Schuurman, T.; Van Breda, A.; Weel, J.F.L.; Beld, M.; Ten Berge, I.J.M.; Wertheim-Van Dillen, P.M.E.; De Jong, M.D. Human Cytomegalovirus DNA in Plasma and Serum Specimens of Renal Transplant Recipients Is Highly Fragmented. J. Clin. Microbiol. 2002, 40, 4105–4113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Pang, X.L.; Mabilangan, C.; Preiksaitis, J.K. Determination of the Biological Form of Human Cytomegalovirus DNA in the Plasma of Solid-Organ Transplant Recipients. J. Infect. Dis. 2017, 215, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinzger, C.; Schmidt, K.; Knapp, J.; Kahl, M.; Beck, R.; Waldman, J.; Hebart, H.; Einsele, H.; Jahn, G. Modification of Human Cytomegalovirus Tropism through Propagation in Vitro Is Associated with Changes in the Viral Genome. J. Gen. Virol. 1999, 80 (Pt 11), 2867–2877. [Google Scholar] [CrossRef]
- Yamane, Y.; Furukawa, T.; Plotkin, S.A. Supernatant Virus Release as a Differentiating Marker between Low Passage and Vaccine Strains of Human Cytomegalovirus. Vaccine 1983, 1, 23–25. [Google Scholar] [CrossRef]
- Dargan, D.J.; Douglas, E.; Cunningham, C.; Jamieson, F.; Stanton, R.J.; Baluchova, K.; McSharry, B.P.; Tomasec, P.; Emery, V.C.; Percivalle, E.; et al. Sequential Mutations Associated with Adaptation of Human Cytomegalovirus to Growth in Cell Culture. J. Gen. Virol. 2010, 91, 1535–1546. [Google Scholar] [CrossRef]
- Connolly, S.A.; Jackson, J.O.; Jardetzky, T.S.; Longnecker, R. Fusing Structure and Function: A Structural View of the Herpesvirus Entry Machinery. Nat. Rev. Microbiol. 2011, 9, 369–381. [Google Scholar] [CrossRef]
- Cooper, R.S.; Heldwein, E.E. Herpesvirus GB: A Finely Tuned Fusion Machine. Viruses 2015, 7, 6552–6569. [Google Scholar] [CrossRef] [Green Version]
- Heldwein, E.E. GH/GL Supercomplexes at Early Stages of Herpesvirus Entry. Curr. Opin. Virol. 2016, 18, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.C.; Kamil, J.P. Pathogen at the Gates: Human Cytomegalovirus Entry and Cell Tropism. Viruses 2018, 10, 704. [Google Scholar] [CrossRef] [Green Version]
- Sathiyamoorthy, K.; Chen, J.; Longnecker, R.; Jardetzky, T.S. The COMPLEXity in Herpesvirus Entry. Curr. Opin. Virol. 2017, 24, 97–104. [Google Scholar] [CrossRef]
- Huber, M.T.; Compton, T. The Human Cytomegalovirus UL74 Gene Encodes the Third Component of the Glycoprotein H-Glycoprotein L-Containing Envelope Complex. J. Virol. 1998, 72, 8191–8197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Nelson, J.A.; Britt, W.J. Glycoprotein H-Related Complexes of Human Cytomegalovirus: Identification of a Third Protein in the GCIII Complex. J. Virol. 1997, 71, 3090–3097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, B.; Scrivano, L.; Ruzcics, Z.; Rupp, B.; Sinzger, C.; Koszinowski, U. Role of Human Cytomegalovirus UL131A in Cell Type-Specific Virus Entry and Release. J. Gen. Virol. 2006, 87, 2451–2460. [Google Scholar] [CrossRef]
- Ryckman, B.J.; Rainish, B.L.; Chase, M.C.; Borton, J.A.; Nelson, J.A.; Jarvis, M.A.; Johnson, D.C. Characterization of the Human Cytomegalovirus GH/GL/UL128-131 Complex That Mediates Entry into Epithelial and Endothelial Cells. J. Virol. 2008, 82, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Shenk, T. Human Cytomegalovirus Virion Protein Complex Required for Epithelial and Endothelial Cell Tropism. Proc. Natl. Acad. Sci. USA 2005, 102, 18153–18158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Lanchy, J.-M.; Ryckman, B.J. Human Cytomegalovirus GH/GL/GO Promotes the Fusion Step of Entry into All Cell Types, Whereas GH/GL/UL128-131 Broadens Virus Tropism through a Distinct Mechanism. J. Virol. 2015, 89, 8999–9009. [Google Scholar] [CrossRef] [Green Version]
- Hahn, G.; Revello, M.G.; Patrone, M.; Percivalle, E.; Campanini, G.; Sarasini, A.; Wagner, M.; Gallina, A.; Milanesi, G.; Koszinowski, U.; et al. Human Cytomegalovirus UL131-128 Genes Are Indispensable for Virus Growth in Endothelial Cells and Virus Transfer to Leukocytes. J. Virol. 2004, 78, 10023–10033. [Google Scholar] [CrossRef] [Green Version]
- Kabanova, A.; Marcandalli, J.; Zhou, T.; Bianchi, S.; Baxa, U.; Tsybovsky, Y.; Lilleri, D.; Silacci-Fregni, C.; Foglierini, M.; Fernandez-Rodriguez, B.M.; et al. Platelet-Derived Growth Factor-α Receptor Is the Cellular Receptor for Human Cytomegalovirus GHgLgO Trimer. Nat. Microbiol. 2016, 1, 16082. [Google Scholar] [CrossRef]
- Stegmann, C.; Hochdorfer, D.; Lieber, D.; Subramanian, N.; Stöhr, D.; Laib Sampaio, K.; Sinzger, C. A Derivative of Platelet-Derived Growth Factor Receptor Alpha Binds to the Trimer of Human Cytomegalovirus and Inhibits Entry into Fibroblasts and Endothelial Cells. PLoS Pathog. 2017, 13, e1006273. [Google Scholar] [CrossRef]
- Wu, Y.; Prager, A.; Boos, S.; Resch, M.; Brizic, I.; Mach, M.; Wildner, S.; Scrivano, L.; Adler, B. Human Cytomegalovirus Glycoprotein Complex GH/GL/GO Uses PDGFR-α as a Key for Entry. PLoS Pathog. 2017, 13, e1006281. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Martin, N.; Marcandalli, J.; Huang, C.S.; Arthur, C.P.; Perotti, M.; Foglierini, M.; Ho, H.; Dosey, A.M.; Shriver, S.; Payandeh, J.; et al. An Unbiased Screen for Human Cytomegalovirus Identifies Neuropilin-2 as a Central Viral Receptor. Cell 2018, 174, 1158–1171.e19. [Google Scholar] [CrossRef] [Green Version]
- Stegmann, C.; Abdellatif, M.E.A.; Laib Sampaio, K.; Walther, P.; Sinzger, C. Importance of Highly Conserved Peptide Sites of Human Cytomegalovirus GO for Formation of the GH/GL/GO Complex. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.J.; Adler, B.; Sampaio, K.L.; Digel, M.; Jahn, G.; Ettischer, N.; Stierhof, Y.-D.; Scrivano, L.; Koszinowski, U.; Mach, M.; et al. UL74 of Human Cytomegalovirus Contributes to Virus Release by Promoting Secondary Envelopment of Virions. J. Virol. 2008, 82, 2802–2812. [Google Scholar] [CrossRef] [Green Version]
- Laib Sampaio, K.; Stegmann, C.; Brizic, I.; Adler, B.; Stanton, R.J.; Sinzger, C. The Contribution of PUL74 to Growth of Human Cytomegalovirus Is Masked in the Presence of RL13 and UL128 Expression. J. Gen. Virol. 2016, 97, 1917–1927. [Google Scholar] [CrossRef] [Green Version]
- Wille, P.T.; Knoche, A.J.; Nelson, J.A.; Jarvis, M.A.; Johnson, D.C. A Human Cytomegalovirus GO-Null Mutant Fails to Incorporate GH/GL into the Virion Envelope and Is Unable to Enter Fibroblasts and Epithelial and Endothelial Cells. J. Virol. 2010, 84, 2585–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanton, R.J.; Baluchova, K.; Dargan, D.J.; Cunningham, C.; Sheehy, O.; Seirafian, S.; McSharry, B.P.; Neale, M.L.; Davies, J.A.; Tomasec, P.; et al. Reconstruction of the Complete Human Cytomegalovirus Genome in a BAC Reveals RL13 to Be a Potent Inhibitor of Replication. J. Clin. Investig. 2010, 120, 3191–3208. [Google Scholar] [CrossRef] [Green Version]
- Murrell, I.; Tomasec, P.; Wilkie, G.S.; Dargan, D.J.; Davison, A.J.; Stanton, R.J. Impact of Sequence Variation in the UL128 Locus on Production of Human Cytomegalovirus in Fibroblast and Epithelial Cells. J. Virol. 2013, 87, 10489–10500. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.J.; Sampaio, K.L.; Ettischer, N.; Stierhof, Y.-D.; Jahn, G.; Kropff, B.; Mach, M.; Sinzger, C. UL74 of Human Cytomegalovirus Reduces the Inhibitory Effect of GH-Specific and GB-Specific Antibodies. Arch. Virol. 2011, 156, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- Gatherer, D.; Seirafian, S.; Cunningham, C.; Holton, M.; Dargan, D.J.; Baluchova, K.; Hector, R.D.; Galbraith, J.; Herzyk, P.; Wilkinson, G.W.G.; et al. High-Resolution Human Cytomegalovirus Transcriptome. Proc. Natl. Acad. Sci. USA 2011, 108, 19755–19760. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, M.; Mach, M.; Britt, W.J. Human Cytomegalovirus Infection Elicits a Glycoprotein M (GM)/GN-Specific Virus-Neutralizing Antibody Response. J. Virol. 2006, 80, 4591–4600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, W.; Chou, C.; Li, H.; Hai, R.; Patterson, D.; Stolc, V.; Zhu, H.; Liu, F. Functional Profiling of a Human Cytomegalovirus Genome. Proc. Natl. Acad. Sci. USA 2003, 100, 14223–14228. [Google Scholar] [CrossRef] [Green Version]
- Hobom, U.; Brune, W.; Messerle, M.; Hahn, G.; Koszinowski, U.H. Fast Screening Procedures for Random Transposon Libraries of Cloned Herpesvirus Genomes: Mutational Analysis of Human Cytomegalovirus Envelope Glycoprotein Genes. J. Virol. 2000, 74, 7720–7729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Silva, M.C.; Shenk, T. Functional Map of Human Cytomegalovirus AD169 Defined by Global Mutational Analysis. Proc. Natl. Acad. Sci. USA 2003, 100, 12396–12401. [Google Scholar] [CrossRef] [Green Version]
- Sinzger, C.; Hahn, G.; Digel, M.; Katona, R.; Sampaio, K.L.; Messerle, M.; Hengel, H.; Koszinowski, U.; Brune, W.; Adler, B. Cloning and Sequencing of a Highly Productive, Endotheliotropic Virus Strain Derived from Human Cytomegalovirus TB40/E. J. Gen. Virol. 2008, 89, 359–368. [Google Scholar] [CrossRef]
- Schuessler, A.; Sampaio, K.L.; Scrivano, L.; Sinzger, C. Mutational Mapping of UL130 of Human Cytomegalovirus Defines Peptide Motifs within the C-Terminal Third as Essential for Endothelial Cell Infection. J. Virol. 2010, 84, 9019–9026. [Google Scholar] [CrossRef] [Green Version]
- Talbot, P.; Almeida, J.D. Human Cytomegalovirus: Purification of Enveloped Virions and Dense Bodies. J. Gen. Virol. 1977, 36, 345–349. [Google Scholar] [CrossRef]
- Tischer, B.K.; Smith, G.A.; Osterrieder, N. En Passant Mutagenesis: A Two Step Markerless Red Recombination System. Methods Mol. Biol. 2010, 634, 421–430. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Gerna, G.; Sarasini, A.; Patrone, M.; Percivalle, E.; Fiorina, L.; Campanini, G.; Gallina, A.; Baldanti, F.; Revello, M.G. Human Cytomegalovirus Serum Neutralizing Antibodies Block Virus Infection of Endothelial/Epithelial Cells, but Not Fibroblasts, Early during Primary Infection. J. Gen. Virol. 2008, 89, 853–865. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, M.; Stanton, R.; Kamil, J.; Ryckman, B.J. Expression Levels of Glycoprotein O (GO) Vary between Strains of Human Cytomegalovirus, Influencing the Assembly of GH/GL Complexes and Virion Infectivity. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Yu, Q.; Wechsler, A.; Ryckman, B.J. Comparative Analysis of GO Isoforms Reveals That Strains of Human Cytomegalovirus Differ in the Ratio of GH/GL/GO and GH/GL/UL128-131 in the Virion Envelope. J. Virol. 2013, 87, 9680–9690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Nguyen, C.C.; Ryckman, B.J.; Britt, W.J.; Kamil, J.P. A Viral Regulator of Glycoprotein Complexes Contributes to Human Cytomegalovirus Cell Tropism. Proc. Natl. Acad. Sci. USA 2015, 112, 4471–4476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, B. A Viral Pilot for HCMV Navigation? Viruses 2015, 7, 3857–3862. [Google Scholar] [CrossRef] [Green Version]
- Bronzini, M.; Luganini, A.; Dell’Oste, V.; De Andrea, M.; Landolfo, S.; Gribaudo, G. The US16 Gene of Human Cytomegalovirus Is Required for Efficient Viral Infection of Endothelial and Epithelial Cells. J. Virol. 2012, 86, 6875–6888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Kamil, J.P. Viral Regulation of Cell Tropism in Human Cytomegalovirus. J. Virol. 2016, 90, 626–629. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Read, C.; Nguyen, C.C.; Siddiquey, M.N.A.; Shang, C.; Hall, C.M.; von Einem, J.; Kamil, J.P. The Human Cytomegalovirus Nonstructural Glycoprotein UL148 Reorganizes the Endoplasmic Reticulum. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| UL55stop_for | ggaatccaggatctggtgcctggtagtctgcgttaacttgtgaatcgtccgtctgggttaagcggtttcctcatcttctacaggatgacgacgataagtaggg |
| UL55stop_rev | gagtagcagaagttccacgagtagaagatgaggaaaccgcttaacccagacggacgattcacaagttaacgcagactaccacaaccaattaaccaattctgattag |
| UL55stop_short_for | ggaatccaggatctggtgcc |
| UL73stop_for | taagcatcgtggcggtggtgtgatggagtggaacacactatgattaggtcttttggtttaatcggtagtggcaagttccaaaggatgacgacgataagtaggg |
| UL73stop_rev | tgctagcagtcgacgtattgttggaacttgccactaccgattaaaccaaaagacctaatcatagtgtgttccactccatcacaaccaattaaccaattctgattag |
| UL73stop_short_for | taagcatcgtggcggtggtg |
| UL73stop_for M3 | caacgtgatgagaccacatgctcacaatgatttttacaattgacattgtacatcgcattagtatgagctttcactgtccagaggatgacgacgataagtaggg |
| UL73stop_rev M3 | tattccaccaggctgcaaagctggacagtgaaagctcatactaatgcgatgtacaatgtcaattgtaaaaatcattgtgagcaaccaattaaccaattctgattag |
| UL73stop_for_short M3 | caacgtgatgagaccacatg |
| UL75 stop_for | cgctatgcggcccggcctccccttctacctcaccgtcttctaggtctacctccttagttgactaccttcgcaacgatatggaggatgacgacgataagtaggg |
| UL75 stop_rev | cttcggatgcggcgtctgcgccatatcgttgcgaaggtagtcaactaaggaggtagacctagaagacggtgaggtagaaggcaaccaattaaccaattctgattag |
| UL75 stop_short_for | cgctatgcggcccggcctcc |
| UL100stop_for | cgtggactttgaaaggctcaacatgtcggcctacaacgtatgacacctgcacacgccttaacttttcttagactcggtgcaaggatgacgacgataagtaggg |
| UL100stop_rev | acacggcgtagcacaccaactgcaccgagtctaagaaaagttaaggcgtgtgcaggtgtcatacgttgtaggccgacatgtcaaccaattaaccaattctgattag |
| UL100stop_for_short | cgtggactttgaaaggctca |
| UL115stop_for | ctctcatcgtgccgcagacttgatgtgccgccgcccggattgaggcttctctttctcataaggaccggtggtactgctgtgaggatgacgacgataagtaggg |
| UL115stop_rev | tgggcagcagaaggcaacaccacagcagtaccaccggtccttatgagaaagagaagcctcaatccgggcggcggcacatcacaaccaattaaccaattctgattag |
| UL115stop_short_for | ctctcatcgtgccgcagact |
| Kanamycin universal reverse | caaccaattaaccaattctga |
| Primer | Sequence (5′-3′) |
|---|---|
| UL128stop_for | acggctgagattcgcgggatcgtcaccaccatgacctagtcattgacatgacaggtcgtacacaacaaggatgacgacgataagt |
| UL128stop_rev | gtagttgcagctcgtcagtttgttgtgtacgacctgtcatgtcaatgactaggtcatggtggtgacgcaaccaattaaccaattctga |
| UL128stop_short_for | acggctgagattcgcgggat |
| UL130stop_for | ctgcctgcttctgtgcgcggtttgggcaacgccctgtctgtagtctccgtggtcgtaactaacagcaaaccagaatccaggatgacgacgataagt |
| UL130stop_rev | gtttagaccatggcggggacggattctggtttgctgttagttacgaccacggagactacagacagggcgttgcccaaacaaccaattaaccaattctga |
| UL130stop_short_for | ctgcctgcttctgtgcgcgg |
| UL131 stop_for | gtctgtttgtctgtgcgccgtggtgctgggtcagtgccagtaggaaaccgcggaataaaacgattattaccgagtaccaggatgacgacgataagt |
| UL131 stop_rev | agcacgcgtcccagtaatgcggtactcggtaataatcgttttattccgcggtttcctactggcactgacccagcaccacaaccaattaaccaattctga |
| UL131 stop_short_for | gtctgtttgtctgtgcgccg |
| Kanamycin universal reverse | caaccaattaaccaattctga |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weiler, N.; Paal, C.; Adams, K.; Calcaterra, C.; Fischer, D.; Stanton, R.J.; Stöhr, D.; Laib Sampaio, K.; Sinzger, C. Role of Envelope Glycoprotein Complexes in Cell-Associated Spread of Human Cytomegalovirus. Viruses 2021, 13, 614. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040614
Weiler N, Paal C, Adams K, Calcaterra C, Fischer D, Stanton RJ, Stöhr D, Laib Sampaio K, Sinzger C. Role of Envelope Glycoprotein Complexes in Cell-Associated Spread of Human Cytomegalovirus. Viruses. 2021; 13(4):614. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040614
Chicago/Turabian StyleWeiler, Nina, Caroline Paal, Kerstin Adams, Christopher Calcaterra, Dina Fischer, Richard James Stanton, Dagmar Stöhr, Kerstin Laib Sampaio, and Christian Sinzger. 2021. "Role of Envelope Glycoprotein Complexes in Cell-Associated Spread of Human Cytomegalovirus" Viruses 13, no. 4: 614. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040614