
RuvB-Like Protein 2 Interacts with the NS1 Protein of Influenza A Virus and Affects Apoptosis That Is Counterbalanced by Type I Interferons


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Virus Infection, and Plasmid Preparation
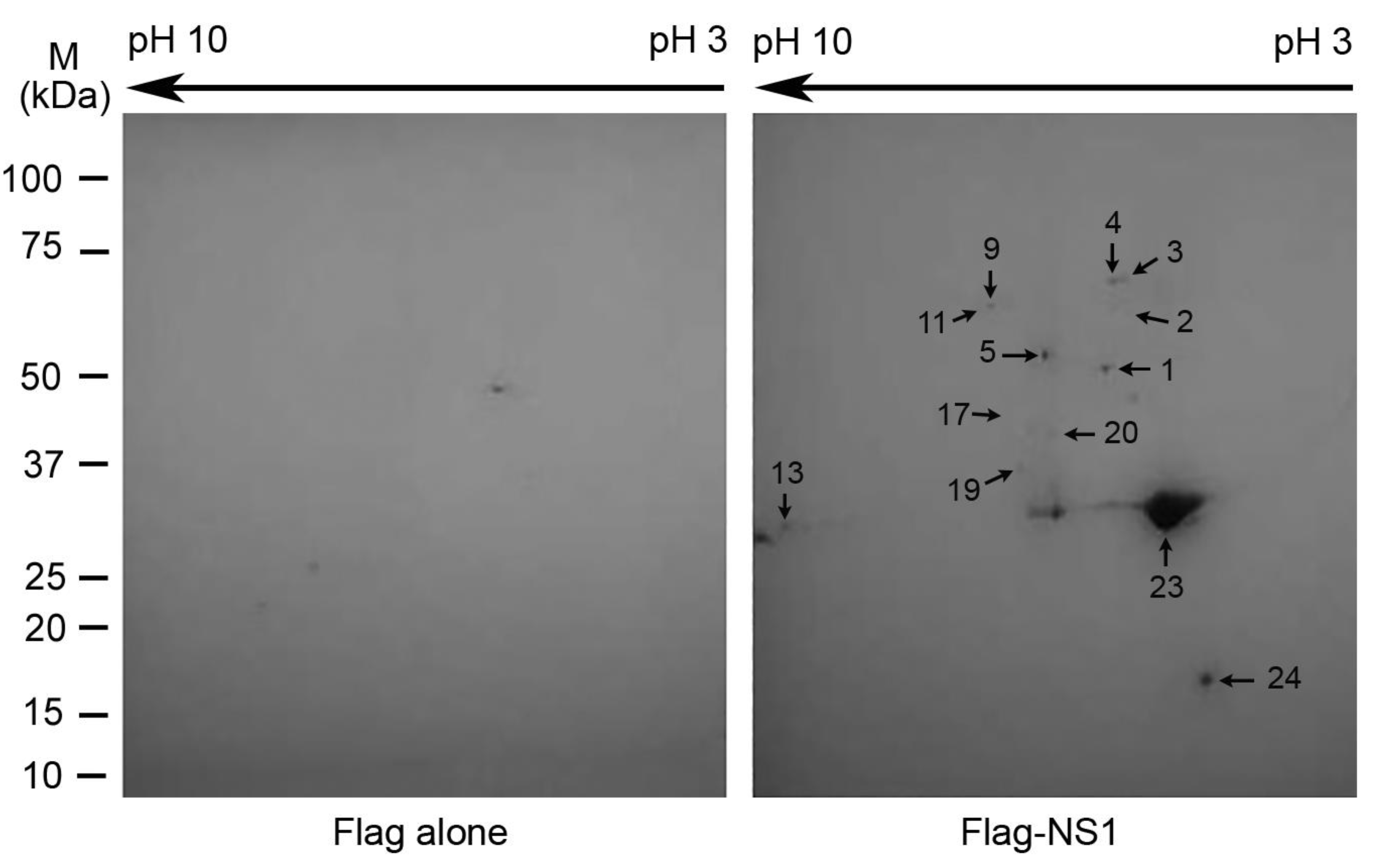
2.2. Two-Dimensional Gel Electrophoresis (2-DE), LC-MS/MS Analysis, and Database Search
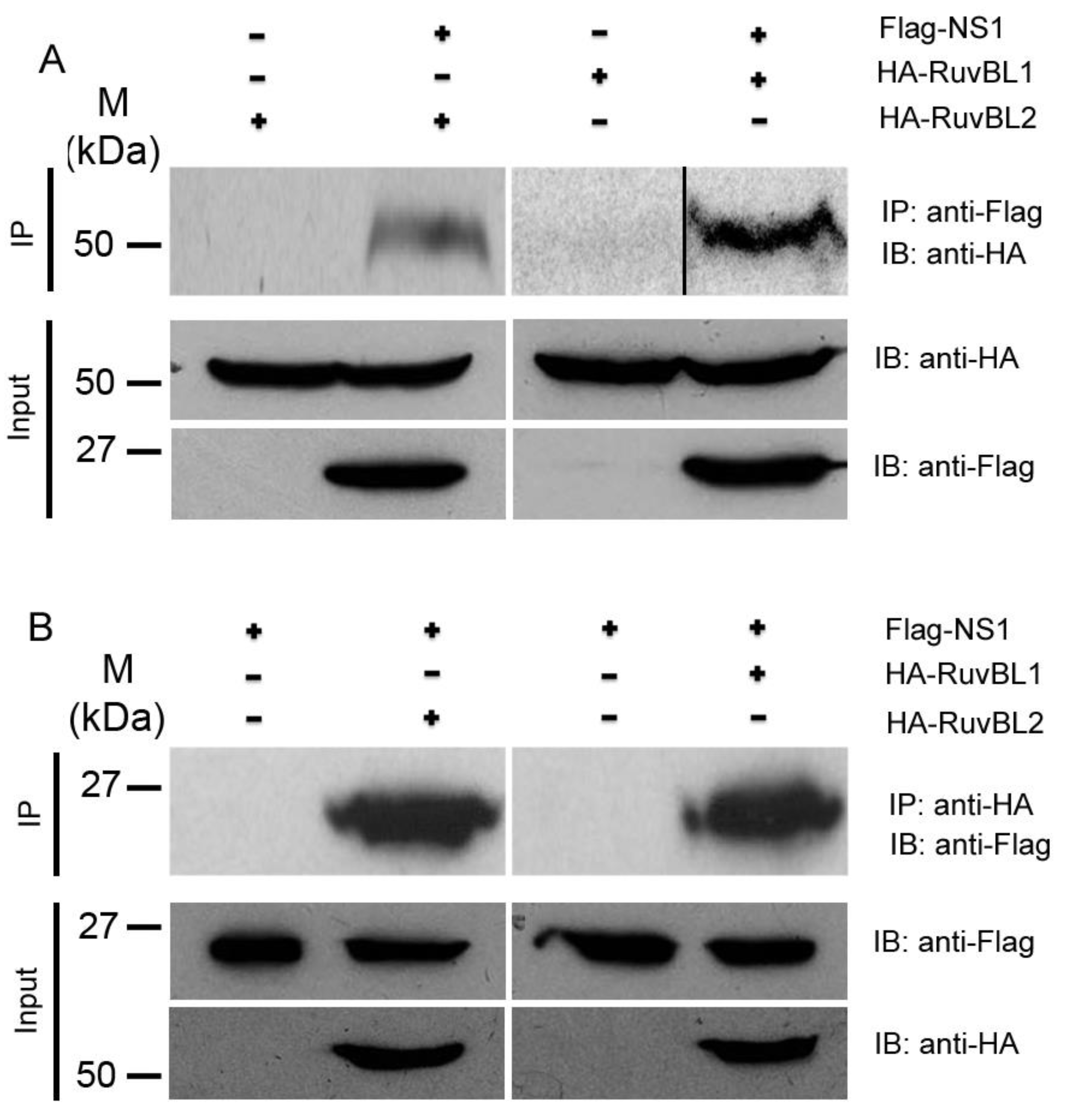
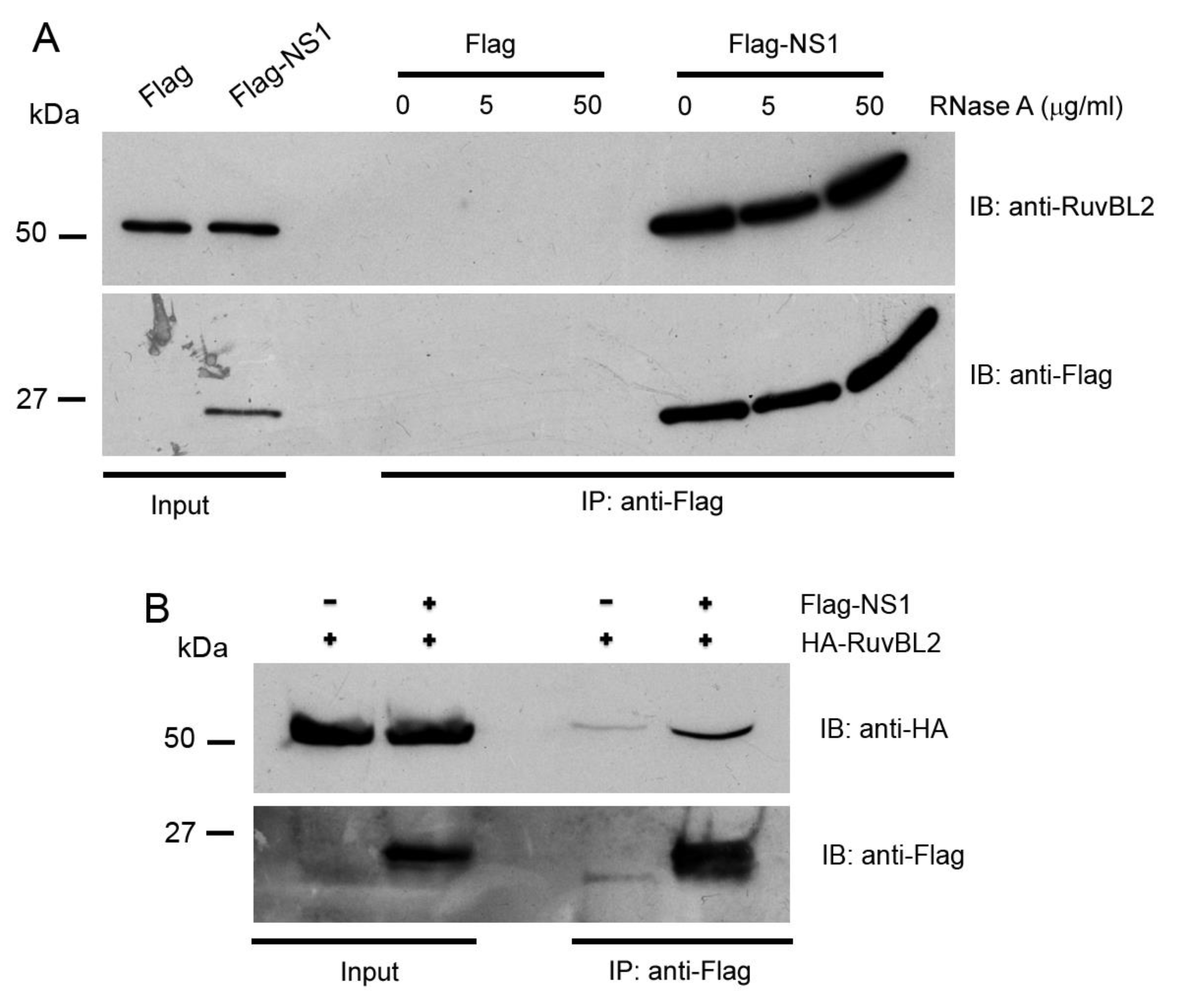
2.3. Immunoprecipitation (IP)
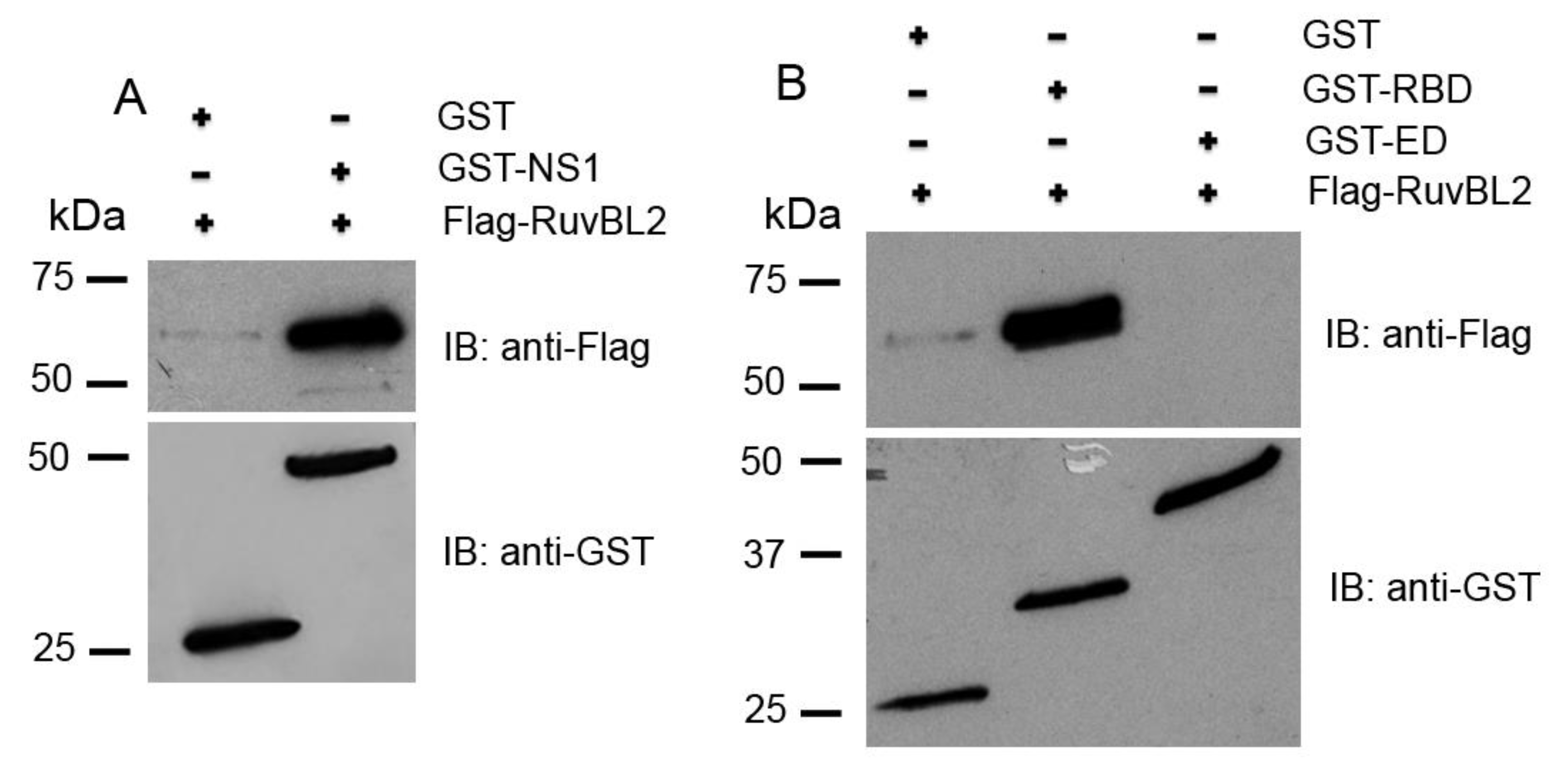
2.4. Protein Expression and In-Vitro Binding Assay
2.5. siRNAs and Overexpression
2.6. Western Blotting
2.7. Caspase 3/7 Assay
2.8. Pretreatment of Vero Cells with Recombinant IFN-α
2.9. Statistical Analysis
3. Results
3.1. Identifications of the Proteins That Potentially Interact with the NS1 Protein
3.2. RuvBL1 and RuvBL2 Interact with the NS1
3.3. RuvBL2 Physically Binds the RNA-Binding Domain (RBD) of the NS1
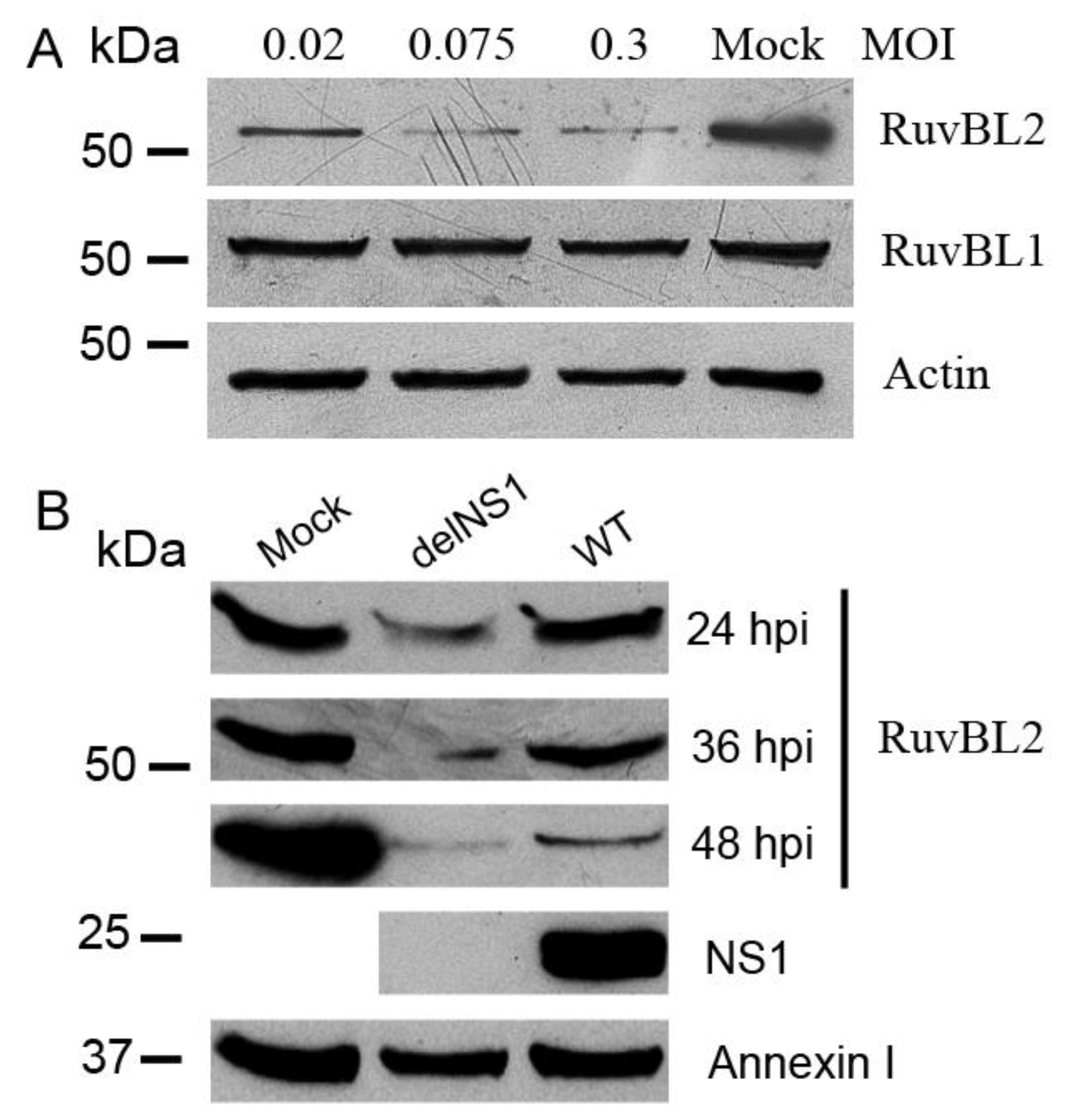
3.4. IAV Infection Leads to Reductions in RuvBL2 Protein Abundance in Infected Cells, and the NS1 Inhibits the Infection-Induced Reduction in RuvBL2 Abundance
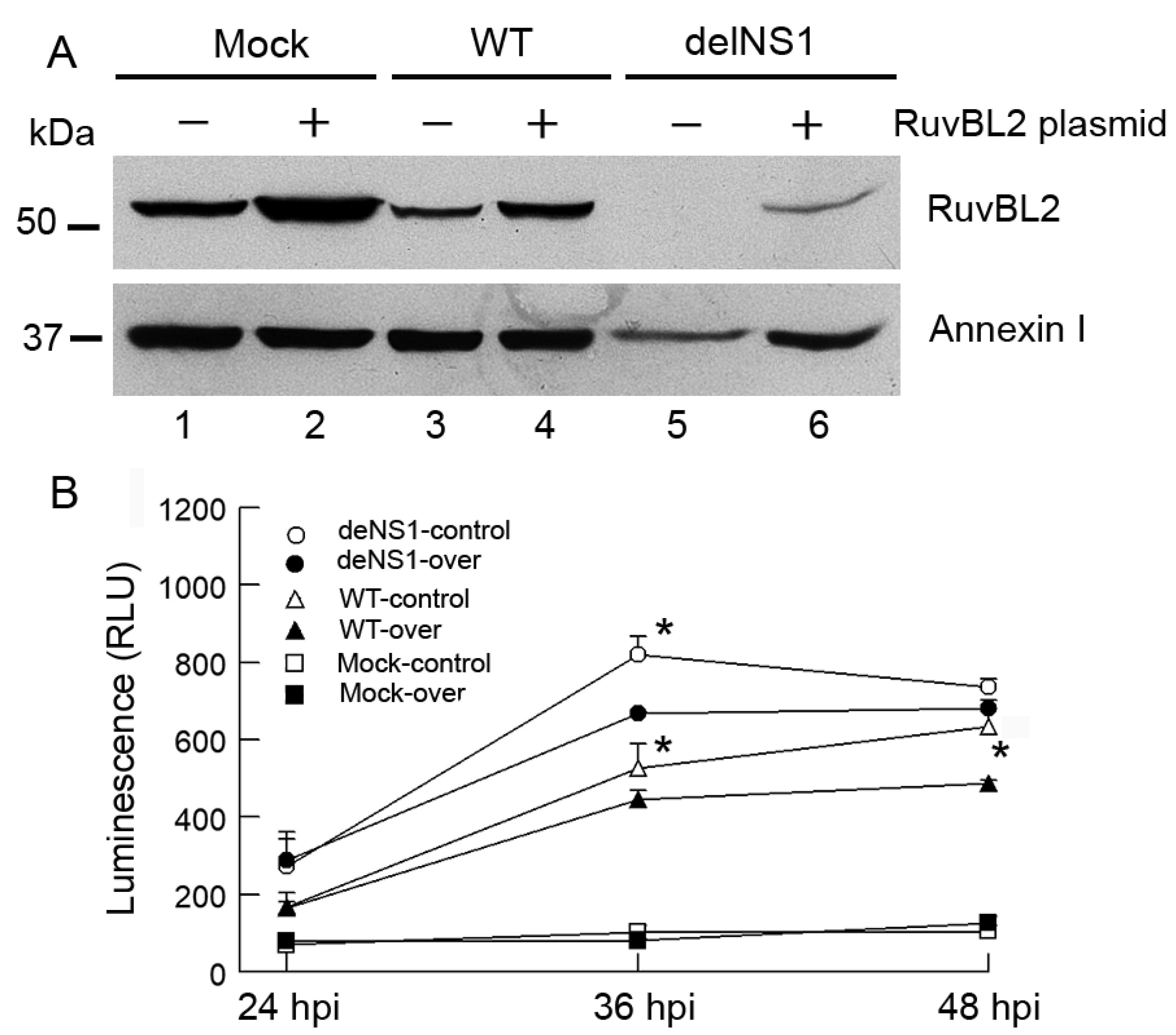
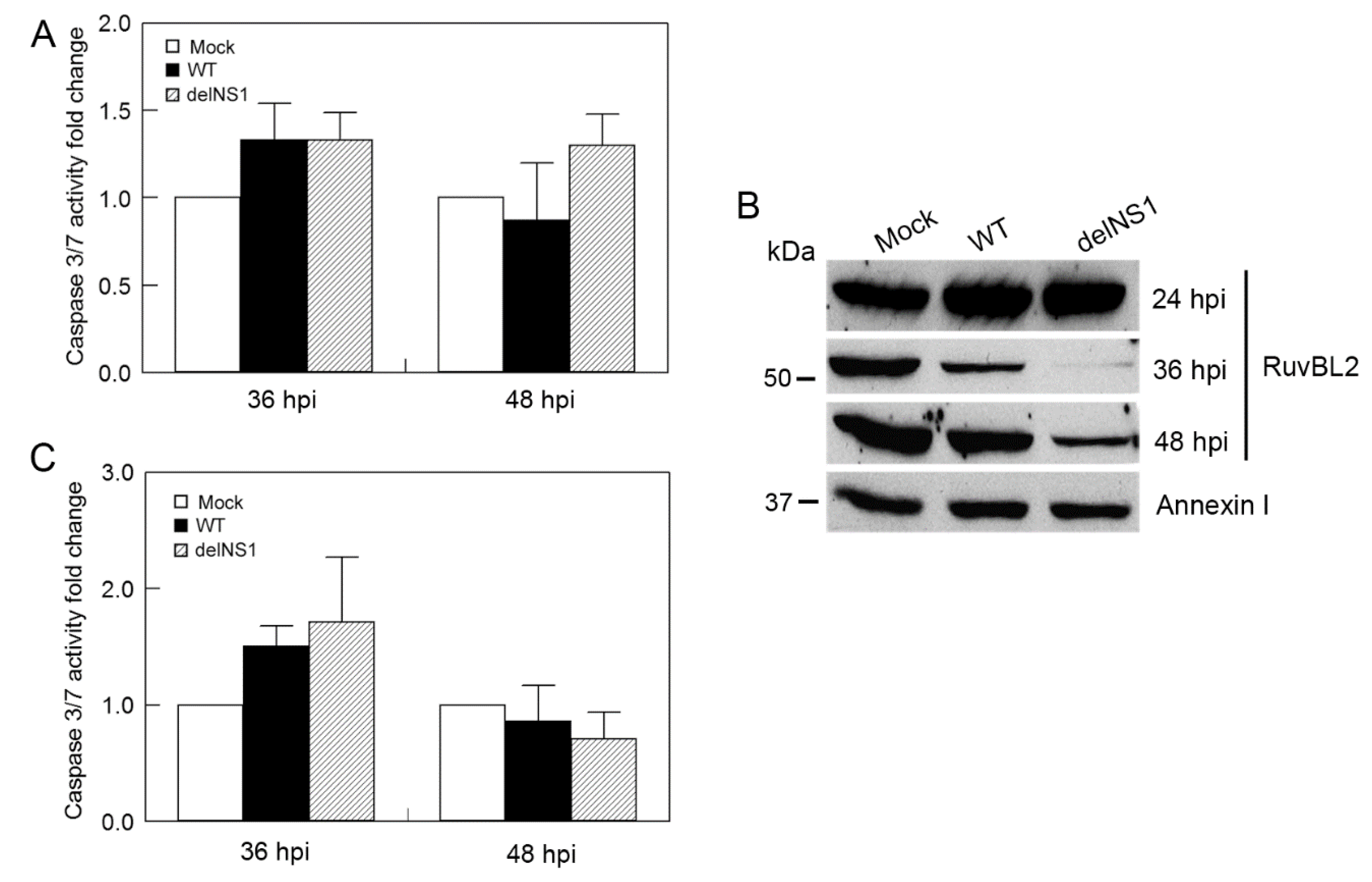
3.5. IAV Infection Induces Host Cell Apoptosis by Suppressing Cellular RuvBL2 Protein Abundance
3.6. Type I IFNs Counteract the Viral Infection-Induced, RuvBL2-Regulated Apoptosis
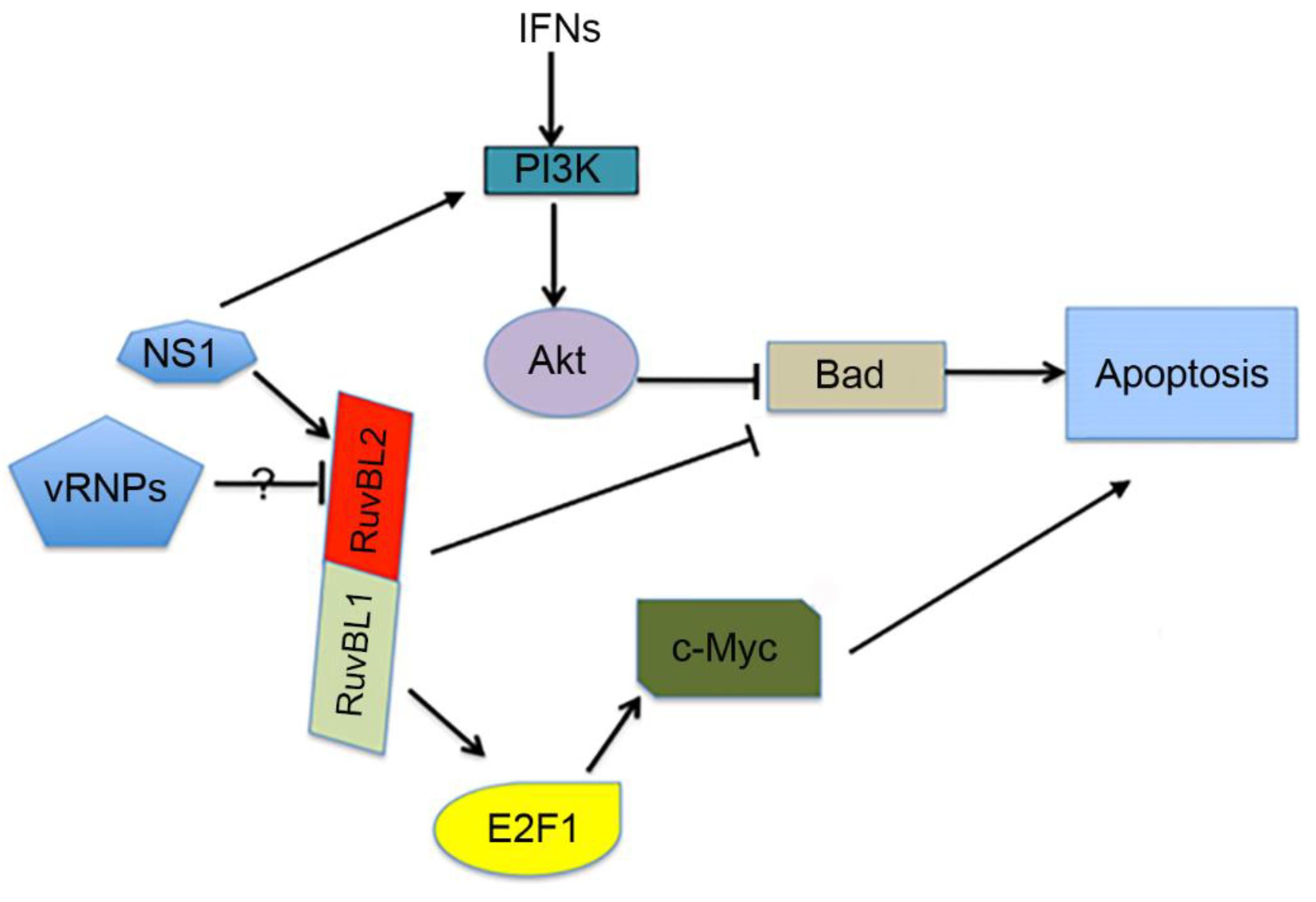
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-DE | two-dimensional gel electrophoresis |
| Bad | Bcl-2 antagonist of cell death |
| delNS1 | influenza A virus lacking the gene encoding the non-structural NS1 protein |
| ED | effector domain |
| GST | glutathione S-transferase |
| hnRNP | heterogeneous nuclear ribonucleoprotein |
| hpi | hours post-infection |
| IAV | influenza A virus |
| IB | immunoblotting |
| IFN | interferon |
| IP | immunoprecipitation |
| MOI | multiplicity of infection |
| NS1 | non-structural protein 1 |
| PARP | poly(ADP-ribose) polymerase |
| PI3K | phosphatidylinositol 3-kinase |
| RBD | RNA-binding domain |
| PR8 virus | influenza A/PR/8/34 virus |
| RuvBL | RuvB-like protein |
| siRNA | small interfering RNA |
| vRNP | viral ribonucleoprotein |
| WT | wild-type |
References
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89 Pt 1, 1–47. [Google Scholar] [CrossRef]
- Ayllon, J.; García-Sastre, A. The NS1 Protein: A Multitasking Virulence Factor. Curr. Top. Microbiol. Immunol. 2015, 386, 73–107. [Google Scholar] [CrossRef] [PubMed]
- Krug, R.M. Functions of the influenza A virus NS1 protein in antiviral defense. Curr. Opin. Virol. 2015, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Barber, G.N. Host defense, viruses and apoptosis. Cell Death Differ. 2001, 8, 113–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurokawa, M.; Koyama, A.H.; Yasuoka, S.; Adachi, A. Influenza virus overcomes apoptosis by rapid multiplication. Int. J. Mol. Med. 1999, 3, 527–557. [Google Scholar] [CrossRef] [PubMed]
- Hale, B.G.; Randall, R.E.; Ortín, J.; Jackson, D. The multifunctional NS1 protein of influenza A viruses. J. Gen. Virol. 2008, 89 Pt 10, 2359–2376. [Google Scholar] [CrossRef]
- Mibayashi, M.; Martínez-Sobrido, L.; Loo, Y.-M.; Cárdenas, W.; Gale, M.; García-SastreA. Inhibition of Retinoic Acid-Inducible Gene I-Mediated Induction of Beta Interferon by the NS1 Protein of Influenza A Virus. J. Virol. 2007, 81, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opitz, B.; Rejaibi, A.; Dauber, B.; Eckhard, J.; Vinzing, M.; Schmeck, B.; Hippenstiel, S.; Suttorp, N.; Wolff, T. IFNbeta induction by influenza A virus is mediated by RIG-I which is regulated by the viral NS1 protein. Cell. Microbiol. 2007, 9, 930–938. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.Y.; Wang, W.; Baker, C.C.; Krug, R.M. The 3′-end-processing factor CPSF is required for the splicing of single-intron pre-mRNAs in vivo. RNA 2001, 7, 920–931. [Google Scholar] [CrossRef] [Green Version]
- Nemeroff, M.E.; Barabino, S.M.; Li, Y.; Keller, W.; Krug, R.M. Influenza virus NS1 protein interacts with the cellular 30 kDa subunit of CPSF and inhibits 3′end formation of cellular pre-mRNAs. Mol. Cell 1998, 1, 991–1000. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Wolff, T.; Pleschka, S.; Planz, O.; Beermann, W.; Bode, J.G.; Schmolke, M.; Ludwig, S. Influenza A Virus NS1 Protein Activates the PI3K/Akt Pathway To Mediate Antiapoptotic Signaling Responses. J. Virol. 2007, 81, 3058–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hale, B.G.; Jackson, D.; Chen, Y.-H.; Lamb, R.A.; Randall, R.E. Influenza A virus NS1 protein binds p85beta and activates phosphatidylinositol-3-kinase signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 14194–14199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nano, N.; Houry, W.A. Chaperone-like activity of the AAA+ proteins Rvb1 and Rvb2 in the assembly of various complexes. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20110399. [Google Scholar] [CrossRef] [Green Version]
- Neuwald, A.F.; Aravind, L.; Spouge, J.L.; Koonin, E.V. AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999, 9, 27–43. [Google Scholar] [PubMed]
- Ogura, T.; Wilkinson, A.J. AAA+superfamily ATPases: Common structure-diverse function. Genes Cells 2001, 6, 575–597. [Google Scholar] [CrossRef] [PubMed]
- Gorynia, S.; Bandeiras, T.; Pinho, F.G.; McVey, C.; Vonrhein, C.; Round, A.; Svergun, D.I.; Donner, P.; Matias, P.M.; Carrondo, M.A. Structural and functional insights into a dodecameric molecular machine—The RuvBL1/RuvBL2 complex. J. Struct. Biol. 2011, 176, 279–291. [Google Scholar] [CrossRef]
- Dauden, M.I.; López-Perrote, A.; Llorca, O. RUVBL1–RUVBL2 AAA-ATPase: A versatile scaffold for multiple complexes and functions. Curr. Opin. Struct. Biol. 2020, 67, 78–85. [Google Scholar] [CrossRef]
- Gorynia, S.; Matias, P.M.; Bandeiras, T.M.; Donner, P.; Carrondo, M.A. Cloning, expression, purification, crystallization and preliminary X-ray analysis of the human RuvBL1–RuvBL2 complex. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2008, 64 Pt 9, 840–846. [Google Scholar] [CrossRef]
- Niewiarowski, A.; Bradley, A.S.; Gor, J.; McKay, A.R.; Perkins, S.J.; Tsaneva, I.R. Oligomeric assembly and interactions within the human RuvB-like RuvBL1 and RuvBL2 complexes. Biochem. J. 2010, 429, 113–125. [Google Scholar] [CrossRef]
- Gallant, P. Control of transcription by Pontin and Reptin. Trends Cell Biol. 2007, 17, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Tarangelo, A.; Lo, N.; Teng, R.; Kim, E.; Le, L.; Watson, D.; Furth, E.E.; Raman, P.; Ehmer, U.; Viatour, P. Recruitment of Pontin/Reptin by E2f1 amplifies E2f transcriptional response during cancer progression. Nat. Commun. 2015, 6, 10028. [Google Scholar] [CrossRef] [Green Version]
- Armenteros-Monterroso, E.; Zhao, L.; Gasparoli, L.; Brooks, T.; Pearce, K.; Mansour, M.R.; Martens, J.H.A.; De Boer, J.; Williams, O. The AAA+ATPase RUVBL2 is essential for the oncogenic function of c-MYB in acute myeloid leukemia. Leukemia 2019, 33, 2817–2829. [Google Scholar] [CrossRef]
- Mao, Y.-Q.; Houry, W.A. The Role of Pontin and Reptin in Cellular Physiology and Cancer Etiology. Front. Mol. Biosci. 2017, 4, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, B.; Menard, L.; Haurie, V.; Taras, D.; Blanc, J.F.; Moreau-Gaudry, F.; Metzler, P.; Hugues, M.; Boyault, S.; Lemiere, S.; et al. Overexpression and role of the ATPase and putative DNA helicase RuvB-like 2 in human hepatocellular carcinoma. Hepatology 2007, 46, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Yenerall, P.; Das, A.K.; Wang, S.; Kollipara, R.K.; Li, L.S.; Villalobos, P.; Flaming, J.; Lin, Y.-F.; Huffman, K.; Timmons, B.C.; et al. RUVBL1/RUVBL2 ATPase Activity Drives PAQosome Maturation, DNA Replication and Radioresistance in Lung Cancer. Cell Chem. Biol. 2020, 27, 105–121.e14. [Google Scholar] [CrossRef]
- Kakugawa, S.; Shimojima, M.; Neumann, G.; Goto, H.; Kawaoka, Y. RuvB-Like Protein 2 Is a Suppressor of Influenza A Virus Polymerases. J. Virol. 2009, 83, 6429–6434. [Google Scholar] [CrossRef] [Green Version]
- Mayer, D.; Molawi, K.; Martínez-Sobrido, L.; Ghanem, A.; Thomas, S.; Baginsky, S.; Grossmann, J.; García-Sastre, A.; Schwemmle, M. Identification of Cellular Interaction Partners of the Influenza Virus Ribonucleoprotein Complex and Polymerase Complex Using Proteomic-Based Approaches. J. Proteome Res. 2007, 6, 672–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, X.; Fu, Y.; Zhu, Y.; Wang, X.; Xuan, Y.; Shang, H.; Goff, S.P.; Gao, G. HIV-1 Exploits the Host Factor RuvB-like 2 to Balance Viral Protein Expression. Cell Host Microbe 2015, 18, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Morwitzer, M.J.; Tritsch, S.R.; Cazares, L.H.; Ward, M.D.; Nuss, J.E.; Bavari, S.; Reid, S.P. Identification of RUVBL1 and RUVBL2 as Novel Cellular Interactors of the Ebola Virus Nucleoprotein. Viruses 2019, 11, 372. [Google Scholar] [CrossRef] [Green Version]
- Yasunaga, A.; Hanna, S.L.; Li, J.; Cho, H.; Rose, P.P.; Spiridigliozzi, A.; Gold, B.; Diamond, M.S.; Cherry, S. Genome-Wide RNAi Screen Identifies Broadly-Acting Host Factors That Inhibit Arbovirus Infection. PLoS Pathog. 2014, 10, e1003914. [Google Scholar] [CrossRef] [Green Version]
- Olanubi, O.; Frost, J.R.; Radko-Juettner, S.; Pelka, P. Suppression of Type I Interferon Signaling by E1A via RuvBL1/Pontin. J. Virol. 2017, 91, e02484-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitsuji, H.; Ujino, S.; Harada, K.; Shimotohno, K. TIP60 Complex Inhibits Hepatitis B Virus Transcription. J. Virol. 2018, 92, 6. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhou, J.; Ruan, C.; Du, Y. Inhibition of Type I Interferon Production via Suppressing IKK-Gamma Expression: A New Strategy for Counteracting Host Antiviral Defense by Influenza A Viruses? J. Proteome Res. 2012, 11, 217–223. [Google Scholar] [CrossRef] [Green Version]
- García-Sastre, A.; Egorov, A.; Matassova, D.; Brandtbc, S.; Levy, D.E.; Durbin, J.E.; Palese, P.; Musterbc, T. Influenza A Virus Lacking the NS1 Gene Replicates in Interferon-Deficient Systems. Virology 1998, 252, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zheng, M.; Wang, P.; Mok, B.W.-Y.; Liu, S.; Lau, S.-Y.; Chen, P.; Liu, Y.-C.; Liu, H.; Chen, Y.; et al. An NS-segment exonic splicing enhancer regulates influenza A virus replication in mammalian cells. Nat. Commun. 2017, 8, 14751. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Zhou, J.; Du, Y. Estrogen Receptor Alpha Interacts with Mitochondrial Protein HADHB and Affects Beta-Oxidation Activity. Mol. Cell. Proteom. 2012, 11, M111-011056. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhou, J.; Du, Y. hnRNP A2/B1 interacts with influenza A viral protein NS1 and inhibits virus replication potentially through suppressing NS1 RNA/protein levels and NS1 mRNA nuclear export. Virology 2014, 449, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhou, J.; Wang, Y.; Mason, R.J.; Funk, C.J.; Du, Y. Proteome Alterations in Primary Human Alveolar Macrophages in Response to Influenza A Virus Infection. J. Proteome Res. 2012, 11, 4091–4101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, N.J.; Lemm, I.; Ingelfinger, D.; Schneider, C.; Hossbach, M.; Urlaub, H.; Luhrmann, R. Assembly and maturation of the U3 snoRNP in the nucleoplasm in a large dynamic multiprotein complex. Mol. Cell 2004, 16, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; He, Y.; Chen, Y.; Kung, H.-F.; He, M.-L. Potent inhibition of human enterovirus 71 replication by type I interferon subtypes. Antivir. Ther. 2011, 16, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlos, T.S.; Fearns, R.; Randall, R.E. Interferon-Induced Alterations in the Pattern of Parainfluenza Virus 5 Transcription and Protein Synthesis and the Induction of Virus Inclusion Bodies. J. Virol. 2005, 79, 14112–14121. [Google Scholar] [CrossRef] [Green Version]
- Kamma, H.; Horiguchi, H.; Wan, L.; Matsui, M.; Fujiwara, M.; Fujimoto, M.; Yazawa, T.; Dreyfuss, G. Molecular characterization of the hnRNP A2/B1 proteins: Tissue-specific expression and novel isoforms. Exp. Cell Res. 1999, 246, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Liu, C.-H.; Zhou, L.; Krug, R.M. Cellular DDX21 RNA Helicase Inhibits Influenza A Virus Replication but Is Counteracted by the Viral NS1 Protein. Cell Host Microbe 2014, 15, 484–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gack, M.U.; Albrecht, R.; Urano, T.; Inn, K.-S.; Huang, I.-C.; Carnero, E.; Farzan, M.; Inoue, S.; Jung, J.U.; García-Sastre, A. Influenza A Virus NS1 Targets the Ubiquitin Ligase TRIM25 to Evade Recognition by the Host Viral RNA Sensor RIG-I. Cell Host Microbe 2009, 5, 439–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Chen, L.-M.; Zeng, H.; Gomez, J.A.; Plowden, J.; Fujita, T.; Katz, J.M.; Donis, R.O.; Sambhara, S. NS1 Protein of Influenza A Virus Inhibits the Function of Intracytoplasmic Pathogen Sensor, RIG-I. Am. J. Respir. Cell Mol. Biol. 2007, 36, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.C.; Yates, J.R., 3rd. The application of mass spectrometry to membrane proteomics. Nat. Biotechnol. 2003, 21, 262–267. [Google Scholar] [CrossRef]
- Du, Y.-C.; Gu, S.; Zhou, J.; Wang, T.; Cai, H.; MacInnes, M.A.; Bradbury, E.M.; Chen, X. The Dynamic Alterations of H2AX Complex during DNA Repair Detected by a Proteomic Approach Reveal the Critical Roles of Ca2+/Calmodulin in the Ionizing Radiation-induced Cell Cycle Arrest. Mol. Cell. Proteom. 2006, 5, 1033–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmyter, J.; Melnick, J.L.; Rawls, W.E. Defectiveness of interferon production and of rubella virus interference in a line of African green monkey kidney cells (Vero). J. Virol. 1968, 2, 955–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, J.E.; Datan, E.; Matassov, D.; Zakeri, Z.F. Lack of Bax Prevents Influenza A Virus-Induced Apoptosis and Causes Diminished Viral Replication. J. Virol. 2009, 83, 8233–8246. [Google Scholar] [CrossRef] [Green Version]
- Tran, A.T.; Cortens, J.P.; Du, Q.; Wilkins, J.A.; Coombs, K.M. Influenza Virus Induces Apoptosis via BAD-Mediated Mitochondrial Dysregulation. J. Virol. 2013, 87, 1049–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurzer, W.J.; Planz, O.; Ehrhardt, C.; Giner, M.; Silberzahn, T.; Pleschka, S.; Ludwig, S. Caspase 3 activation is essential for efficient influenza virus propagation. EMBO J. 2003, 22, 2717–2728. [Google Scholar] [CrossRef] [Green Version]
- Herold, S.; Ludwig, S.; Pleschka, S.; Wolff, T. Apoptosis signaling in influenza virus propagation, innate host defense, and lung injury. J. Leukoc. Biol. 2012, 92, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Ruuth, K.; Carlsson, L.; Hallberg, B.; Lundgren, E. Interferon-α Promotes Survival of Human Primary B-Lymphocytes via Phosphatidylinositol 3-Kinase. Biochem. Biophys. Res. Commun. 2001, 284, 583–586. [Google Scholar] [CrossRef]
- Yang, C.H.; Murti, A.; Pfeffer, S.R.; Kim, J.G.; Donner, D.B.; Pfeffer, L.M. Interferon alpha/beta promotes cell survival by activating nuclear factor kappa B through phosphatidylinositol 3-kinase and Akt. J. Biol. Chem. 2001, 276, 13756–13761. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt Phosphorylation of BAD Couples Survival Signals to the Cell-Intrinsic Death Machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Del Peso, L.; González-García, M.; Page, C.; Herrera, R.; Nuñez, G. Interleukin-3-Induced Phosphorylation of BAD through the Protein Kinase Akt. Science 1997, 278, 687–689. [Google Scholar] [CrossRef]
- Dugan, K.A.; Wood, M.A.; Cole, M.D. TIP49, but not TRRAP, modulates c-Myc and E2F1 dependent apoptosis. Oncogene 2002, 21, 5835–5843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maher, S.G.; Romero-Weaver, A.L.; Scarzello, A.J.; Gamero, A.M. Interferon: Cellular executioner or white knight? Curr. Med. Chem. 2007, 14, 1279–1289. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wambach, M.; Katze, M.G.; Krug, R.M. Binding of the influenza virus NS1 protein to double-stranded RNA inhibits the activation of the protein kinase that phosphorylates the elF-2 translation initiation factor. Virology 1995, 214, 222–228. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Spot # b | Protein Name | SwissProt acc.# | MW (kDa)/pI | Mascot Protein Score | Unique Peptide Count | Exclusive Spectrum Count | Sequence Coverage (%) |
|---|---|---|---|---|---|---|---|---|
| ATPase | 1 | RuvBL2 | Q9Y230 | 51.2/5.5 | 1859 | 25 | 90 | 53 |
| 5 | RuvBL1 | Q9Y265 | 50.2/6.0 | 1402 | 22 | 57 | 56 | |
| Heterogeneous nuclear ribonucleoprotein | 2 | hnRNP K | P61978 | 51.0/5.4 | 1062 | 17 | 37 | 39 |
| 9 | hnRNP L | P14866 | 64.1/8.5 | 1113 | 15 | 25 | 39 | |
| 11 | hnRNP Q | O60506 | 69.6/8.7 | 675 | 13 | 24 | 18 | |
| 13 | hnRNP A2/B1 | P22626 | 37.4/9.0 | 1207 | 16 | 33 | 55 | |
| 17 | hnRNP D0 | Q14103 | 38.4/7.6 | 554 | 5 | 9 | 21 | |
| 19 | hnRNP D-like | O14979 | 46.4/9.6 | 499 | 8 | 12 | 16 | |
| 20 | hnRNP A/B | Q99729 | 36.2/8.2 | 362 | 2 | 7 | 11 | |
| Heat shock protein | 3 | Heat shock cognate 71 kDa protein | P11142 | 70.9/5.4 | 1566 | 18 | 45 | 32 |
| 4 | Heat shock 70 kDa protein 1A/1B | P0DMV8/P0DMV9 | 70.1/5.5 | 1395 | 16 | 45 | 32 | |
| Influenza A viral protein | 23 | NS1 | P03496 | 25.9/6.2 | 1279 | 18 | 107 | 80 |
| 24 | NS1 | P03496 | 25.9/6.2 | 1123 | 16 | 68 | 76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhou, J.; Mackintosh, S.G.; Du, Y. RuvB-Like Protein 2 Interacts with the NS1 Protein of Influenza A Virus and Affects Apoptosis That Is Counterbalanced by Type I Interferons. Viruses 2021, 13, 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061038
Wang Y, Zhou J, Mackintosh SG, Du Y. RuvB-Like Protein 2 Interacts with the NS1 Protein of Influenza A Virus and Affects Apoptosis That Is Counterbalanced by Type I Interferons. Viruses. 2021; 13(6):1038. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061038
Chicago/Turabian StyleWang, Yimeng, Jianhong Zhou, Samuel G. Mackintosh, and Yuchun Du. 2021. "RuvB-Like Protein 2 Interacts with the NS1 Protein of Influenza A Virus and Affects Apoptosis That Is Counterbalanced by Type I Interferons" Viruses 13, no. 6: 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061038