
Subclinical Infection and Transmission of Clade 2.3.4.4 H5N6 Highly Pathogenic Avian Influenza Virus in Mandarin Duck (Aix galericulata) and Domestic Pigeon (Columbia livia domestica)

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Viruses
2.3. Experimental Design
2.4. Viral Quantification
2.5. Statistical Analysis
2.6. Serology
2.7. Immunohistochemistry
3. Results
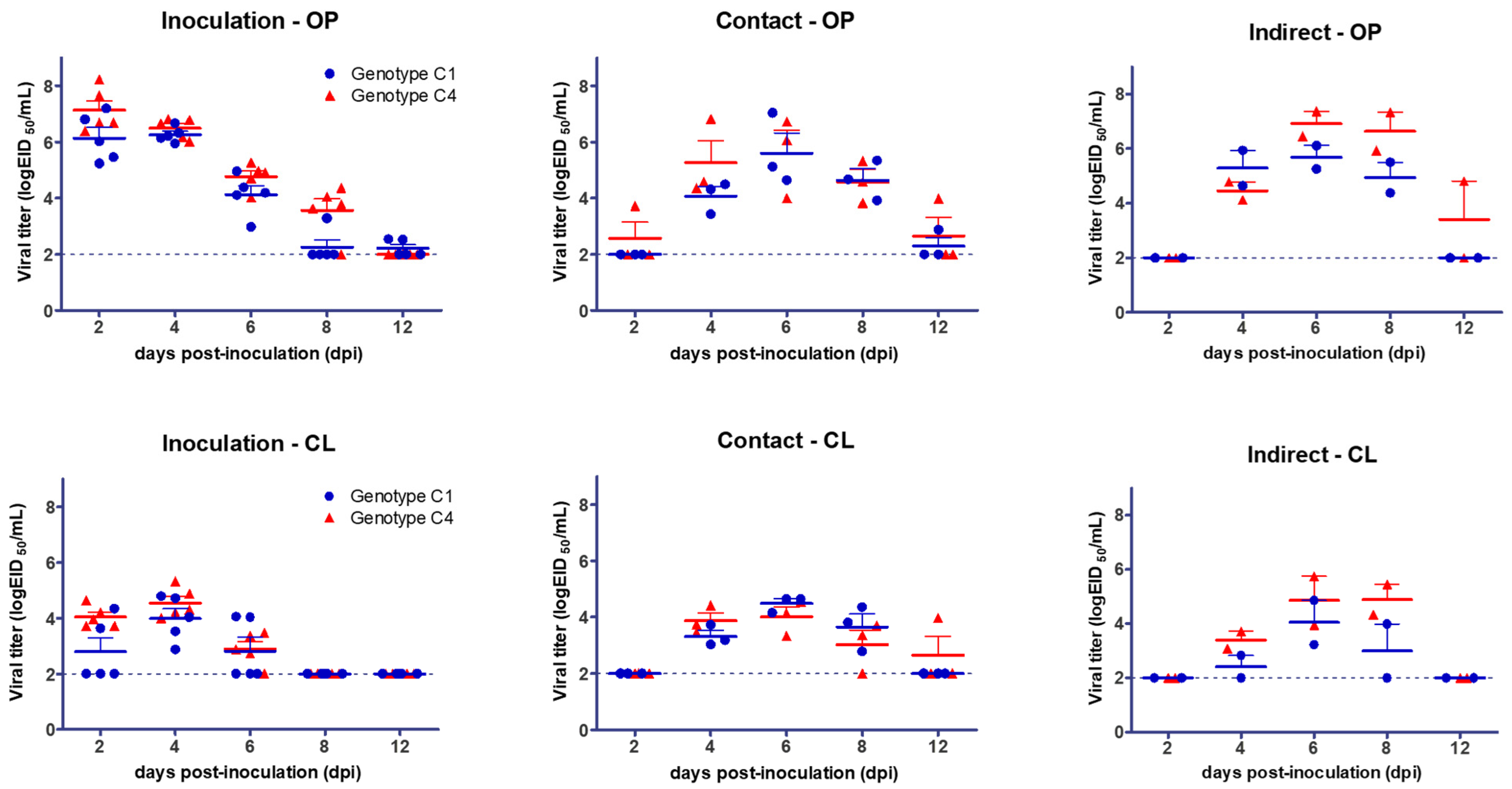
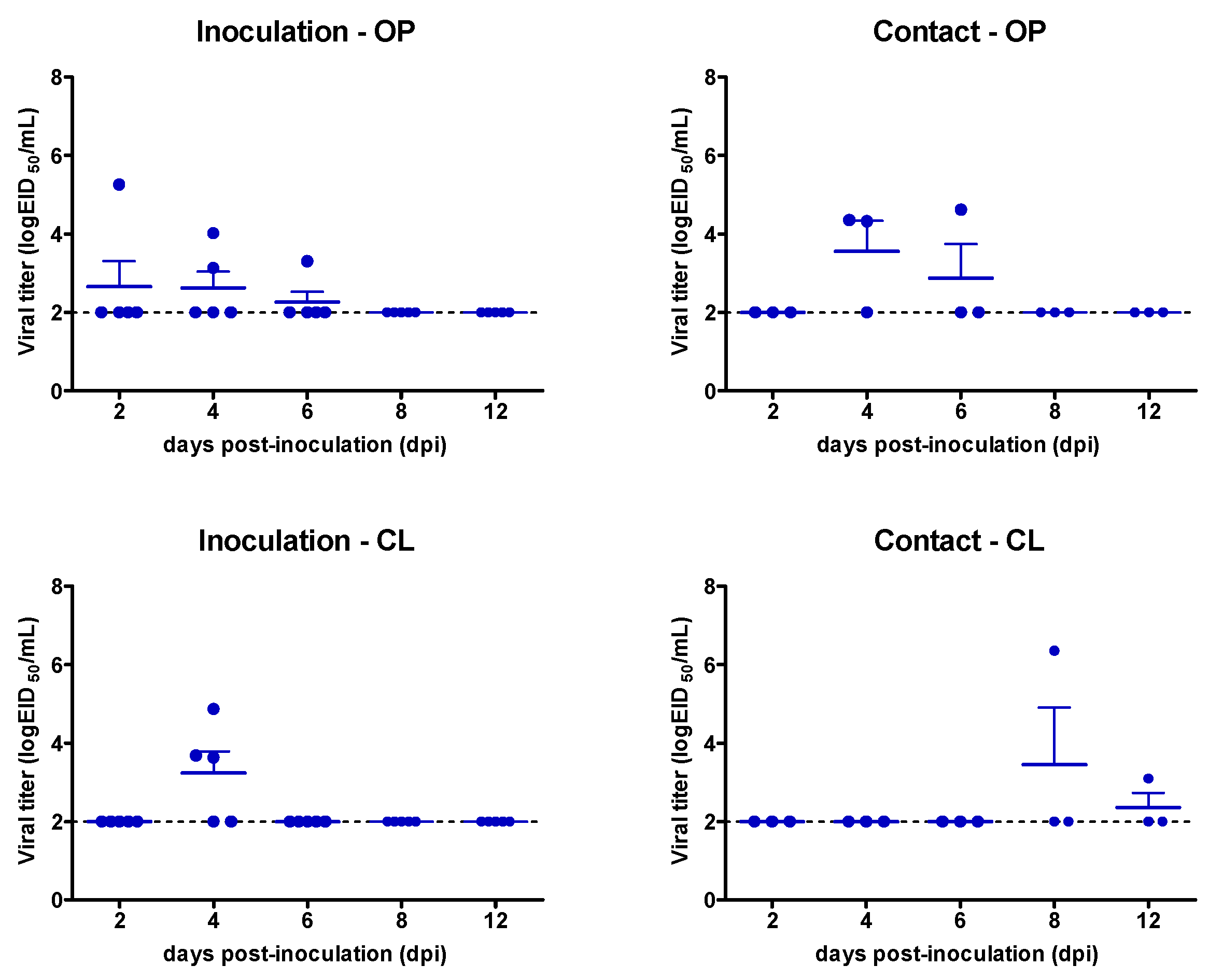
3.1. Clinical Signs of Infection, Viral Shedding, and Transmission
3.2. Serology
3.3. Immunohistochemistry
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sonnberg, S.; Webby, R.J.; Webster, R.G. Natural history of highly pathogenic avian influenza H5N. Virus Res. 2013, 178, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Bi, Y.; Wong, G.; Gray, G.C.; Gao, G.F.; Li, S. Epidemiology, Evolution and Recent Outbreaks of Avian Influenza Virus in China. J. Virol. 2015, 89, 8671–8676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Gu, M.; Zhong, L.; Duan, Z.; Zhang, Y.; Zhu, Y.; Zhao, G.; Zhao, M.; Chen, Z.; Hu, S.; et al. Characterization of three H5N5 and one H5N8 highly pathogenic avian influenza viruses in China. Vet. Microbiol. 2013, 163, 351–357. [Google Scholar] [CrossRef]
- Gu, M.; Liu, W.; Cao, Y.; Peng, D.; Wang, X.; Wan, H.; Zhao, G.; Xu, Q.; Zhang, W.; Song, Q.; et al. Novel Reassortant Highly Pathogenic Avian Influenza (H5N5) Viruses in Domestic Ducks, China. Emerg. Infect. Dis. 2011, 17, 1060–1063. [Google Scholar] [CrossRef] [PubMed]
- Bouwstra, R.; Heutink, R.; Bossers, A.; Harders, F.; Koch, G.; Elbers, A. Full-Genome Sequence of Influenza A(H5N8) Virus in Poultry Linked to Sequences of Strains from Asia, the Netherlands, 2014. Emerg. Infect. Dis. 2015, 21, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Harder, T.; Maurer-Stroh, S.; Pohlmann, A.; Starick, E.; Höreth-Böntgen, D.; Al., T.H.E.; Pannwitz, G.; Teifke, J.; Gunalan, V.; Lee, R.T.; et al. Influenza A(H5N8) Virus Similar to Strain in Korea Causing Highly Pathogenic Avian Influenza in Germany. Emerg. Infect. Dis. 2015, 21, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Ip, H.S.; Torchetti, M.K.; Crespo, R.; Kohrs, P.; DeBruyn, P.; Mansfield, K.G.; Baszler, T.; Badcoe, L.; Bodenstein, B.; Shearn-Bochsler, V.; et al. Novel Eurasian Highly Pathogenic Avian Influenza A H5 Viruses in Wild Birds, Washington, USA, 2014. Emerg. Infect. Dis. 2015, 21, 886–890. [Google Scholar] [CrossRef]
- Kanehira, K.; Uchida, Y.; Takemae, N.; Hikono, H.; Tsunekuni, R.; Saito, T. Characterization of an H5N8 influenza A virus isolated from chickens during an outbreak of severe avian influenza in Japan in April 2014. Arch. Virol. 2015, 160, 1629–1643. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kang, H.M.; Lee, E.K.; Song, B.M.; Jeong, J.; Kwon, Y.K.; Kim, H.R.; Lee, K.J.; Hong, M.S.; Jang, I.; et al. Novel reassortant influenza A(H5N8) viruses, South Korea, 2014. Emerg. Infect. Dis. 2014, 20, 1087–1089. [Google Scholar] [CrossRef]
- Claes, F.; Morzaria, S.P.; Donis, R.O. Emergence and dissemination of clade 2.3.4.4 H5Nx influenza viruses—How is the Asian HPAI H5 lineage maintained. Curr. Opin. Virol. 2016, 16, 158–163. [Google Scholar] [CrossRef]
- Wong, F.Y.; Phommachanh, P.; Kalpravidh, W.; Chanthavisouk, C.; Gilbert, J.; Bingham, J.; Davies, K.R.; Cooke, J.; Eagles, D.; Phiphakhavong, S.; et al. Reassortant Highly Pathogenic Influenza A(H5N6) Virus in Laos. Emerg. Infect. Dis. 2015, 21, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Firestone, S.M.; Stevenson, M.A.; Young, N.D.; Sims, L.D.; Chu, D.H.; Nguyen, T.N.; Van Nguyen, L.; Le, T.T.; Van Nguyen, H.; et al. A systematic study towards evolutionary and epidemiological dynamics of currently predominant H5 highly pathogenic avian influenza viruses in Vietnam. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Takemae, N.; Tsunekuni, R.; Sharshov, K.; Tanikawa, T.; Uchida, Y.; Ito, H.; Soda, K.; Usui, T.; Sobolev, I.; Shestopalov, A.; et al. Five distinct reassortants of H5N6 highly pathogenic avian influenza A viruses affected Japan during the winter of 2016–2017. Virology 2017, 512, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-K.; Song, B.-M.; Lee, Y.-N.; Heo, G.-B.; Bae, Y.-C.; Joh, S.-J.; Park, S.-C.; Choi, K.-S.; Lee, H.-J.; Jang, I.; et al. Multiple novel H5N6 highly pathogenic avian influenza viruses, South Korea, 2016. Infect. Genet. Evol. 2017, 51, 21–23. [Google Scholar] [CrossRef]
- Bi, Y.; Chen, Q.; Wang, Q.; Chen, J.; Jin, T.; Wong, G.; Quan, C.; Liu, J.; Wu, J.; Yin, R.; et al. Genesis, Evolution and Prevalence of H5N6 Avian Influenza Viruses in China. Cell Host Microbe 2016, 20, 810–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Antigenic and Genetic Characteristics of Zoonotic Influenza A Viruses and Development of Candidate Vaccine Viruses for Pandemic Preparedness. Available online: https://www.who.int/influenza/vaccines/virus/201909_zoonotic_vaccinevirusupdate.pdf?ua=1 (accessed on 6 April 2021).
- Tsunekuni, R.; Yaguchi, Y.; Kashima, Y.; Yamashita, K.; Takemae, N.; Mine, J.; Tanikawa, T.; Uchida, Y.; Saito, T. Spatial transmission of H5N6 highly pathogenic avian influenza viruses among wild birds in Ibaraki Prefecture, Japan, 2016–2017. Arch. Virol. 2018, 163, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Kuiken, T. Changing Role of Wild Birds in the Epidemiology of Avian Influenza A Viruses. Int. Rev. Cytol. 2018, 100, 279–307. [Google Scholar] [CrossRef]
- World Health Organization. Avian Influenza Weekly Update Number 784. Available online: https://www.who.int/docs/default-source/wpro---documents/emergency/surveillance/avian-influenza/ai-20210319.pdf?sfvrsn=30d65594_115 (accessed on 6 April 2021).
- Kwon, J.-H.; Lee, D.-H.; Swayne, D.E.; Noh, J.-Y.; Yuk, S.-S.; Erdene-Ochir, T.-O.; Hong, W.-T.; Jeong, J.-H.; Jeong, S.; Gwon, G.-B.; et al. Reassortant Clade 2.3.4.4 Avian Influenza A(H5N6) Virus in a Wild Mandarin Duck, South Korea, 2016. Emerg. Infect. Dis. 2017, 23, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Cui, J.; Song, Y.; Song, H.; Zhao, Z.; Wu, S.; Qu, N.; Wang, N.; Ouyang, G.; Liao, M. New Reassortant H5N6 Highly Pathogenic Avian Influenza Viruses in Southern China, 2014. Front. Microbiol. 2016, 7, 754. [Google Scholar] [CrossRef]
- Li, H.; Li, Q.; Li, B.; Guo, Y.; Xing, J.; Xu, Q.; Liu, L.; Zhang, J.; Qi, W.; Jia, W.; et al. Continuous Reassortment of Clade 2.3.4.4 H5N6 Highly Pathogenetic Avian Influenza Viruses Demonstrating High Risk to Public Health. Pathogens 2020, 9, 670. [Google Scholar] [CrossRef]
- Lever, C. The Mandarin Duck, 1st ed.; Bloomsbury Publishing: London, UK, 2013. [Google Scholar]
- Lee, D.-H.; Park, J.-K.; Youn, H.-N.; Lee, Y.-N.; Lim, T.-H.; Kim, M.-S.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; Song, C.-S. Surveillance and Isolation of HPAI H5N1 from Wild Mandarin Ducks (Aix galericulata). J. Wildl. Dis. 2011, 47, 994–998. [Google Scholar] [CrossRef] [Green Version]
- Sakoda, Y.; Ito, H.; Uchida, Y.; Okamatsu, M.; Yamamoto, N.; Soda, K.; Nomura, N.; Kuribayashi, S.; Shichinohe, S.; Sunden, Y.; et al. Reintroduction of H5N1 highly pathogenic avian influenza virus by migratory water birds, causing poultry outbreaks in the 2010–2011 winter season in Japan. J. Gen. Virol. 2012, 93, 541–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, J.-H.; Lee, D.-H.; Swayne, D.E.; Noh, J.-Y.; Yuk, S.-S.; Erdene-Ochir, T.-O.; Hong, W.-T.; Jeong, J.-H.; Jeong, S.; Gwon, G.-B.; et al. Highly Pathogenic Avian Influenza A(H5N8) Viruses Reintroduced into South Korea by Migratory Waterfowl, 2014–2015. Emerg. Infect. Dis. 2016, 22, 507–510. [Google Scholar] [CrossRef]
- Usui, T.; Soda, K.; Tomioka, Y.; Ito, H.; Yabuta, T.; Takakuwa, H.; Otsuki, K.; Ito, T.; Yamaguchi, T. Characterization of clade 2.3.4.4 H5N8 highly pathogenic avian influenza viruses from wild birds possessing atypical hemagglutinin polybasic cleavage sites. Virus Genes 2017, 53, 44–51. [Google Scholar] [CrossRef]
- He, G.; Zhou, L.; Zhu, C.; Shi, H.; Li, X.; Wu, D.; Liu, J.; Lv, J.; Hu, C.; Li, Z.; et al. Identification of two novel avian influenza a (H5N6) viruses in wild birds, Shanghai, in 2016. Vet. Microbiol. 2017, 208, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.-H.; Jeong, S.; Lee, N.-H.; Swayne, D.E.; Kim, Y.-J.; Lee, S.-H.; Noh, J.-Y.; Erdene-Ochir, T.-O.; Jeong, J.-H.; Song, C.-S. New Reassortant Clade 2.3.4.4b Avian Influenza A(H5N6) Virus in Wild Birds, South Korea, 2017–18. Emerg. Infect. Dis. 2018, 24, 1953–1955. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.; Lee, D.-H.; Kwon, J.-H.; Kim, Y.-J.; Lee, S.-H.; Cho, A.Y.; Kim, T.-H.; Park, J.-E.; Lee, S.-I.; Song, C.-S. Highly Pathogenic Avian Influenza Clade 2.3.4.4b Subtype H5N8 Virus Isolated from Mandarin Duck in South Korea, 2020. Viruses 2020, 12, 1389. [Google Scholar] [CrossRef]
- European Food Safety Authority(EFSA). Statement on migratory birds and their possible role in the spread of highly pathogenic avian influenza by the Scientific Panel on Animal Health an Welfare (AHAW). EFSA J. 2006, 4, 357. [Google Scholar]
- Liu, Y.; Zhou, J.; Yang, H.; Yao, W.; Bu, W.; Yang, B.; Song, W.; Meng, Y.; Lin, J.; Han, C.; et al. Susceptibility and transmissibility of pigeons to Asian lineage highly pathogenic avian influenza virus subtype H5N1. Avian Pathol. 2007, 36, 461–465. [Google Scholar] [CrossRef]
- Achenbach, J.E.; Bowen, R.A. Transmission of Avian Influenza A Viruses among Species in an Artificial Barnyard. PLoS ONE 2011, 6, 17643. [Google Scholar] [CrossRef]
- Kohls, A.; Lüschow, D.; Lierz, M.; Hafez, H.M. Influenza A Virus Monitoring in Urban and Free-Ranging Pigeon Populations in Germany, 2006–2008. Avian Dis. Dig. 2011, 6, 19–20. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Nakamura, K.; Yamada, M.; Mase, M. Limited Susceptibility of Pigeons Experimentally Inoculated with H5N1 Highly Pathogenic Avian Influenza Viruses. J. Vet. Med. Sci. 2012, 74, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.-H.; Noh, Y.K.; Lee, D.-H.; Yuk, S.-S.; Erdene-Ochir, T.-O.; Noh, J.-Y.; Hong, W.-T.; Jeong, J.-H.; Jeong, S.; Gwon, G.-B.; et al. Experimental infection with highly pathogenic H5N8 avian influenza viruses in the Mandarin duck (Aix galericulata) and domestic pigeon (Columba livia domestica). Vet. Microbiol. 2017, 203, 95–102. [Google Scholar] [CrossRef]
- Brown, J.D.; Stallknecht, D.E.; Berghaus, R.D.; Swayne, D.E. Infectious and Lethal Doses of H5N1 Highly Pathogenic Avian Influenza Virus for House Sparrows (Passer Domesticus) and Rock Pigeons (Columbia Livia). J. Vet. Diagn. Investig. 2009, 21, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Shi, J.; Li, Y.; Shinya, K.; Muramoto, Y.; Zeng, X.; Tian, G.; Kawaoka, Y.; Chen, H. Pathogenicity of Chinese H5N1 highly pathogenic avian influenza viruses in pigeons. Arch. Virol. 2008, 153, 1821–1826. [Google Scholar] [CrossRef]
- Klopfleisch, R.; Werner, O.; Mundt, E.; Harder, T.; Teifke, J.P. Neurotropism of Highly Pathogenic Avian Influenza Virus A/Chicken/Indonesia/2003 (H5N1) in Experimentally Infected Pigeons (Columbia livia f. domestica). Vet. Pathol. 2006, 43, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Smietanka, K.; Minta, Z.; Wyrostek, K.; Jozwiak, M.; Olszewska, M.; Domanska-Blicharz, A.K.; Reichert, A.M.; Pikula, A.; Habyarimana, A.; van den Berg, T. Susceptibility of pigeons to clade 1 and 2.2 high pathogenicity avian influenza H5N1 virus. Avian Dis. 2011, 55, 106–112. [Google Scholar] [CrossRef]
- Liu, K.; Gao, R.; Wang, X.; Han, W.; Ji, Z.; Zheng, H.; Gu, M.; Hu, J.; Liu, X.; Hu, S.; et al. Pathogenicity and transmissibility of clade 2.3.4.4 highly pathogenic avian influenza virus subtype H5N6 in pigeons. Vet. Microbiol. 2020, 247, 108776. [Google Scholar] [CrossRef]
- Abolnik, C.; Stutchbury, S.; Hartman, M.J. Experimental infection of racing pigeons (Columba livia domestica) with highly pathogenic Clade 2.3.4.4 sub-group B H5N8 avian influenza virus. Vet. Microbiol. 2018, 227, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Spackman, E.; Senne, D.A.; Bulaga, L.L.; Myers, T.J.; Perdue, M.L.; Garber, L.P.; Lohman, K.; Daum, L.T.; Suarez, D. Development of Real-Time RT-PCR for the Detection of Avian Influenza Virus. Avian Dis. 2003, 47, 1079–1082. [Google Scholar] [CrossRef]
- World Organization for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.03.04_AI.pdf (accessed on 20 April 2021).
- Reed, L.J.; Muench, H. A Simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Lee, C.-W.; Suarez, D.L. Application of real-time RT-PCR for the quantitation and competitive replication study of H5 and H7 subtype avian influenza virus. J. Virol. Methods 2004, 119, 151–158. [Google Scholar] [CrossRef]
- Yu, J.E.; Kim, M.; Lee, J.-H.; Chang, B.-J.; Song, C.-S.; Nahm, S.-S. Neonatal influenza infection causes pathological changes in the mouse brain. Vet. Res. 2014, 45, 63. [Google Scholar] [CrossRef]
- Olsen, B.; Munster, V.; Wallensten, A.; Waldenström, J.; Osterhaus, A.; Fouchier, R.A.M. Global Patterns of Influenza A Virus in Wild Birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Chmura, A.A.; Gibbons, D.W.; Fleischer, R.C.; Marra, P.P.; Daszak, P. Predicting the global spread of H5N1 avian influenza. Proc. Natl. Acad. Sci. USA 2006, 103, 19368–19373. [Google Scholar] [CrossRef] [Green Version]
- Son, K.; Kim, Y.-K.; Oem, J.-K.; Jheong, W.-H.; Sleeman, J.M.; Jeong, J. Experimental infection of highly pathogenic avian influenza viruses, Clade 2.3.4.4 H5N6 and H5N8, in Mandarin ducks from South Korea. Transbound. Emerg. Dis. 2018, 65, 899–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-M.; Lee, E.-K.; Song, B.-M.; Heo, G.-B.; Jung, J.; Jang, I.; Bae, Y.-C.; Jung, S.C.; Lee, Y.-J. Experimental infection of mandarin duck with highly pathogenic avian influenza A (H5N8 and H5N1) viruses. Vet. Microbiol. 2017, 198, 59–63. [Google Scholar] [CrossRef]
- Abolnik, C.; Fehrsen, J.; Olivier, A.; Van Wyngaardt, W.; Fosgate, G.; Ellis, C. Serological investigation of highly pathogenic avian influenza H5N2 in ostriches (Struthio camelus). Avian Pathol. 2013, 42, 206–214. [Google Scholar] [CrossRef]
- Sánchez-González, R.; Ramis, A.; Nofrarías, M.; Wali, N.; Valle, R.; Pérez, M.; Perlas, A.; Majó, N. Infectivity and pathobiology of H7N1 and H5N8 high pathogenicity avian influenza viruses for pigeons (Columba livia var. domestica). Avian Pathol. 2021, 50, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Xiang, B.; Liang, J.; You, R.; Han, L.; Mei, K.; Chen, L.; Chen, R.; Zhang, Y.; Dai, X.; Gao, P.; et al. Pathogenicity and transmissibility of a highly pathogenic avian influenza virus H5N6 isolated from a domestic goose in Southern China. Vet. Microbiol. 2017, 212, 16–21. [Google Scholar] [CrossRef]
- Ellis, T.M.; Bousfield, R.B.; Bissett, L.A.; Dyrting, K.C.; Luk, G.S.M.; Tsim, S.T.; Sturm-Ramirez, K.; Webster, R.G.; Guan, Y.; Peiris, J.S.M. Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2020. Avian Pathol. 2004, 33, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.M.G.; ElBakrey, R.M.; Ali, H.R.S.; Knudsen, D.E.B.; Eid, A.A.M. Natural infection with highly pathogenic avian influenza virus H5N1 in domestic pigeons (Columba livia) in Egypt. Avian Pathol. 2014, 43, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.M.G.; Mohamed, F.; Eid, A.A.M.; Mor, S.K.; Goyal, S.M. Co-circulation of paramyxo- and influenza viruses in pigeons in Egypt. Avian Pathol. 2017, 46, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Boon, A.C.; Sandbulte, M.R.; Seiler, P.; Webby, R.J.; Songserm, T.; Guan, Y.; Webster, R.G. Role of Terrestrial Wild Birds in Ecology of Influenza A Virus (H5N1). Emerg. Infect. Dis. 2007, 13, 1720–1724. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E.; Prosser, D.J.; Pantin-Jackwood, M.J.; Berlin, A.M.; Stephens, C.B. The Pathogenesis of Clade 2.3.4.4 H5 Highly Pathogenic Avian Influenza Viruses in Ruddy Duck (Oxyura Jamaicensis) and Lesser Scaup (Aythya Affinis). J. Wildl. Dis. 2017, 53, 832–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, R.; Cui, J.; Zhang, S.; Cao, L.; Liu, X.; Kang, Y.; Song, Y.; Gong, L.; Jiao, P.; Liao, M. Pathogenicity and transmission of H5N1 avian influenza viruses in different birds. Vet. Microbiol. 2014, 168, 50–59. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Swayne, D.E. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev. Sci. Tech. 2009, 28, 113–136. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Costa-Hurtado, M.; Shepherd, E.; DeJesus, E.; Smith, D.; Spackman, E.; Kapczynski, D.R.; Suarez, D.; Stallknecht, D.E.; Swayne, D.E. Pathogenicity and Transmission of H5 and H7 Highly Pathogenic Avian Influenza Viruses in Mallards. J. Virol. 2016, 90, 9967–9982. [Google Scholar] [CrossRef] [Green Version]
- Pantin-Jackwood, M.J.; Costa-Hurtado, M.; Bertran, K.; DeJesus, E.; Smith, D.; Swayne, D.E. Infectivity, transmission and pathogenicity of H5 highly pathogenic avian influenza clade 2.3.4.4 (H5N8 and H5N2) United States index viruses in Pekin ducks and Chinese geese. Vet. Res. 2017, 48, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Li, J.; Hu, J.; Jiang, D.; Xing, C.; Zhan, T.; Liu, X. Genetic analysis and biological characteristics of different internal gene origin H5N6 reassortment avian influenza virus in China in 2016. Vet. Microbiol. 2018, 219, 200–211. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Viral Genotype | Group | Clinical Sign (Sick/Total) | Mortality (Dead/Total) | Viral Shedding (Max Mean Viral Titer 1 and dpi) | Serology | |
|---|---|---|---|---|---|---|---|
| NP-ELISA (Mean PI 2) | HI Assay (Mean Titer) | ||||||
| Mandarin duck | C1 | Inoculation | 0/5 | 0/5 | 5/5 (6.26, 4 dpi) | 5/5 (93.91) | 5/5 (5.2) |
| Contact | 0/3 | 0/3 | 3/3 (5.60, 6 dpi) | 3/3 (93.42) | 3/3 (5.67) | ||
| Indirect | 0/2 | 0/2 | 2/2 (5.69, 6 dpi) | 2/2 (92.16) | 2/2 (5.5) | ||
| C4 | Inoculation | 0/5 | 0/5 | 5/5 (7.13, 2 dpi) | 4/5 (95.56) | 5/5 (5.2) | |
| Contact | 0/3 | 0/3 | 3/3 (5.60, 6 dpi) | 3/3 (95.47) | 3/3 (4.33) | ||
| Indirect | 0/2 | 0/2 | 2/2 (6.91, 6 dpi) | 2/2 (94.37) | 2/2 (6) | ||
| Domestic pigeon | C1 | Inoculation | 0/5 | 1/5 | 3/5 (3.24, 4 dpi) | 4/4 3 (93.65) | 1/4 3 (6) |
| Contact | 0/3 | 0/3 | 2/3 (3.56, 4 dpi) | 1/3 (67.05) | 0/3 | ||
| Species | Viral Genotype | Brain | Heart | Trachea | Lung | Liver | Pancreas | Intestine | Kidney | Spleen | Cecal Tonsil | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mandarin duck | C1 | Bird 1 | + | − | + | − | + | − | + | + | + | + |
| Bird 2 | + | + | + | − | + | + | + | + | + | + | ||
| C4 | Bird 1 | + | − | + | + | + | + | + | + | + | + | |
| Bird 2 | − | − | + | + | + | + | + | + | + | + | ||
| Domestic pigeon | C1 | Bird 1 | + | − | + | + | + | + | + | + | + | + |
| Bird 2 | + | − | + | + | + | − | + | + | + | + | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Kwon, J.-H.; Lee, S.-H.; Kim, Y.-J.; Jeong, J.-H.; Park, J.-E.; Jheong, W.-H.; Lee, D.-H.; Song, C.-S. Subclinical Infection and Transmission of Clade 2.3.4.4 H5N6 Highly Pathogenic Avian Influenza Virus in Mandarin Duck (Aix galericulata) and Domestic Pigeon (Columbia livia domestica). Viruses 2021, 13, 1069. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061069
Jeong S, Kwon J-H, Lee S-H, Kim Y-J, Jeong J-H, Park J-E, Jheong W-H, Lee D-H, Song C-S. Subclinical Infection and Transmission of Clade 2.3.4.4 H5N6 Highly Pathogenic Avian Influenza Virus in Mandarin Duck (Aix galericulata) and Domestic Pigeon (Columbia livia domestica). Viruses. 2021; 13(6):1069. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061069
Chicago/Turabian StyleJeong, Sol, Jung-Hoon Kwon, Sun-Hak Lee, Yu-Jin Kim, Jei-Hyun Jeong, Jung-Eun Park, Weon-Hwa Jheong, Dong-Hun Lee, and Chang-Seon Song. 2021. "Subclinical Infection and Transmission of Clade 2.3.4.4 H5N6 Highly Pathogenic Avian Influenza Virus in Mandarin Duck (Aix galericulata) and Domestic Pigeon (Columbia livia domestica)" Viruses 13, no. 6: 1069. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061069