
Quality Assessment and Validation of High-Throughput Sequencing for Grapevine Virus Diagnostics

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants
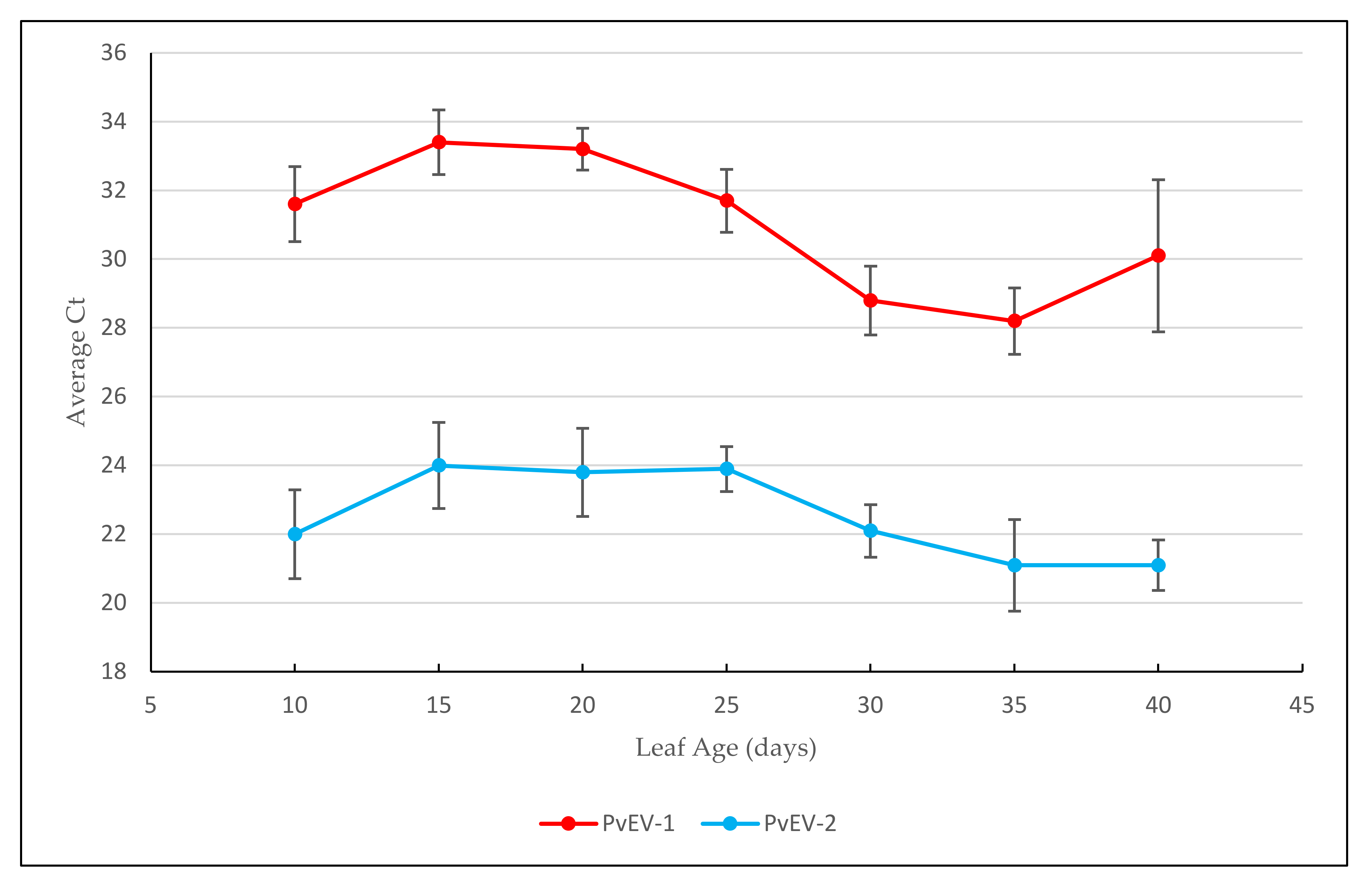
2.2. Relative Concentration of PvEV-1 and PvEV-2 in Common Bean Cv Black Turtle Soup (BTS) at Different Growth Stages
2.3. TNA and dsRNA Extraction
2.4. Preparation of Dilution Series
2.5. HTS
2.6. Determination of an HTS Positive
2.7. Characterizing Novel and Non-Grapevine Viruses
3. Results
3.1. Concentration of PvEV-1 and PvEV-2 in BTS at Different Growth Stages
3.2. Evaluating BTS as a Positive Control
3.3. Evaluation of Our TNA HTS Protocol
3.4. Comparison to dsRNA
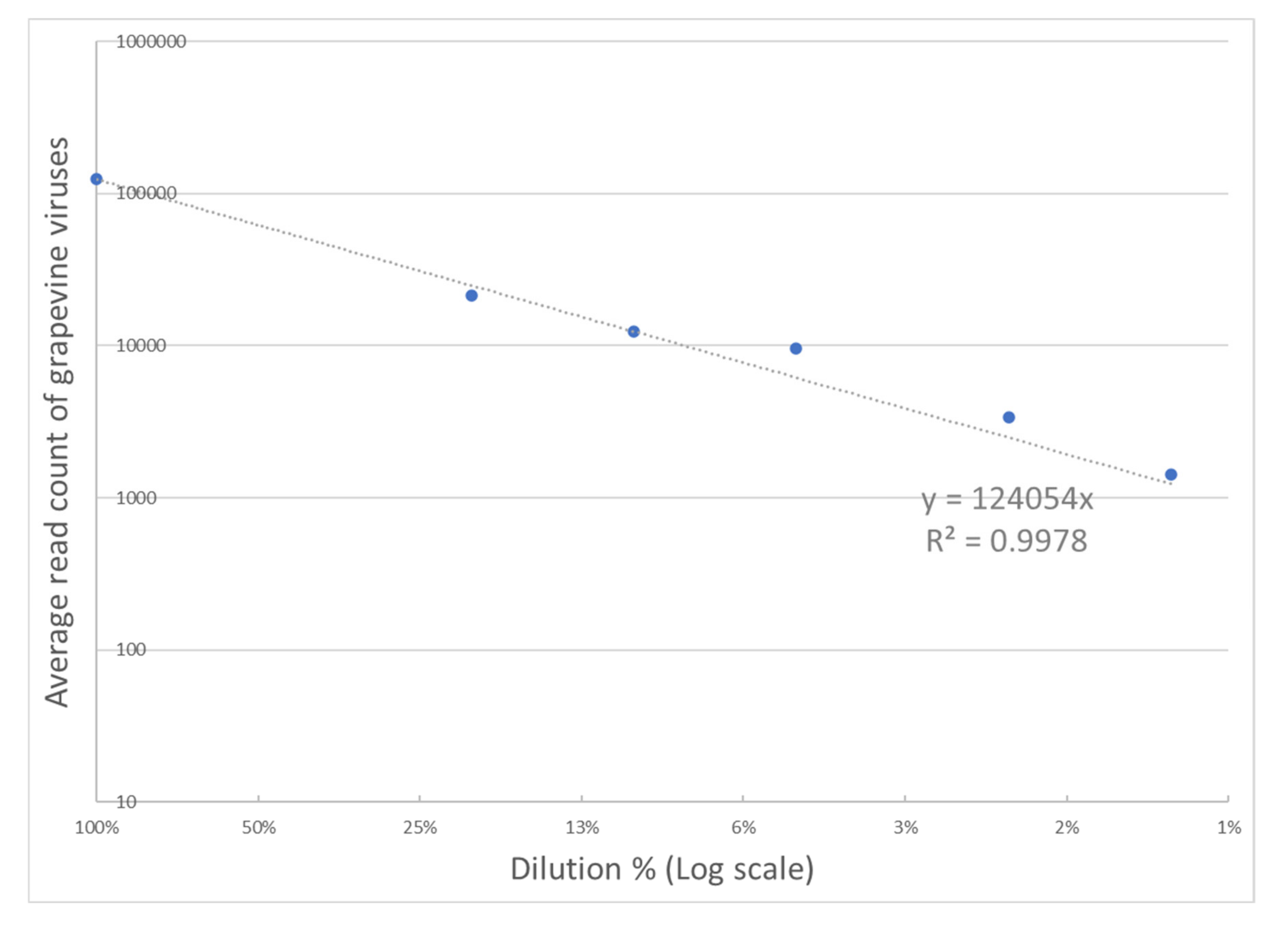
3.5. Dilution Series
3.6. Repeatability
3.7. Reproducibility
4. Discussion
4.1. Sensitivity
4.2. Specificity
4.3. Repeatability and Reproducibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuchs, M. Grapevine viruses: A multitude of diverse species with simple but overall poorly adopted management solutions in the vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Almeida, R.; Daane, K.; Bell, V.; Blaisdell, G.K.; Cooper, M.; Herrbach, E.; Pietersen, G. Ecology and management of grapevine leafroll disease. Front. Microbiol. 2013, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Cheon, J.Y.; Fenton, M.; Gjerdseth, E.; Wang, Q.; Gao, S.; Krovetz, H.; Lu, L.; Shim, L.; Williams, N.; Lybbert, T.J. Heterogeneous Benefits of Virus Screening for Grapevines in California. Am. J. Enol. Vitic. 2020, 71, 231–241. [Google Scholar] [CrossRef]
- Ricketts, K.D.; Gómez, M.I.; Fuchs, M.F.; Martinson, T.E.; Smith, R.J.; Cooper, M.L.; Moyer, M.M.; Wise, A. Mitigating the Economic Impact of Grapevine Red Blotch: Optimizing Disease Management Strategies in U.S. Vineyards. Am. J. Enol. Vitic. 2017, 68, 127–135. [Google Scholar] [CrossRef]
- Fuchs, M.; Almeyda, C.V.; Al Rwahnih, M.; Atallah, S.S.; Cieniewicz, E.J.; Farrar, K.; Foote, W.R.; Golino, D.A.; Gómez, M.I.; Harper, S.J.; et al. Economic Studies Reinforce Efforts to Safeguard Specialty Crops in the United States. Plant Dis. 2020, 105, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Rowhani, A.; Uyemoto, J.K.; Golino, D.A.; Martelli, G.P. Pathogen testing and certification of Vitis and Prunus species. Annu. Rev. Phytopathol. 2005, 43, 261–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constable, F.E.; Connellan, J.; Nicholas, P.; Rodoni, B.C. The reliability of woody indexing for detection of grapevine virus-associated diseases in three different climatic conditions in Australia. Aust. J. Grape Wine Res. 2013, 19, 74–80. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Islas, C.; Rowhani, A. Comparison of Next-Generation Sequencing Versus Biological Indexing for the Optimal Detection of Viral Pathogens in Grapevine. Phytopathology 2015, 105, 758–763. [Google Scholar] [CrossRef] [Green Version]
- Licciardello, G.; Ferraro, R.; Scuderi, G.; Russo, M.; Catara, A.F. A Simulation of the Use of High Throughput Sequencing as Pre-Screening Assay to Enhance the Surveillance of Citrus Viruses and Viroids in the EPPO Region. Agriculture 2021, 11, 400. [Google Scholar] [CrossRef]
- Maree, H.J.; Fox, A.; Al Rwahnih, M.; Boonham, N.; Candresse, T. Application of HTS for Routine Plant Virus Diagnostics: State of the Art and Challenges. Front. Plant Sci. 2018, 9, 1082. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.R.; Constable, F.; Tzanetakis, I.E. Quarantine Regulations and the Impact of Modern Detection Methods. Annu. Rev. Phytopathol. 2016, 54, 189–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golino, D.A.; Fuchs, M.; Al Rwahnih, M.; Farrar, K.; Schmidt, A.; Martelli, G.P. Regulatory Aspects of Grape Viruses and Virus Diseases: Certification, Quarantine, and Harmonization. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 581–598. ISBN 9783319577067. [Google Scholar]
- Villamor, D.E.V.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E. High Throughput Sequencing For Plant Virus Detection and Discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Rowhani, A. Deep sequencing analysis of RNAs from a grapevine showing Syrah decline symptoms reveals a multiple virus infection that includes a novel virus. Virology 2009, 387, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Saldarelli, P.; Giampetruzzi, A.; Maree, H.J.; Al Rwahnih, M. High-Throughput Sequencing: Advantages Beyond Virus Identification. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 625–642. ISBN 9783319577067. [Google Scholar]
- Massart, S.; Candresse, T.; Gil, J.; Lacomme, C.; Predajna, L.; Ravnikar, M.; Reynard, J.-S.; Rumbou, A.; Saldarelli, P.; Škorić, D.; et al. A Framework for the Evaluation of Biosecurity, Commercial, Regulatory, and Scientific Impacts of Plant Viruses and Viroids Identified by NGS Technologies. Front. Microbiol. 2017, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Donaire, L.; Wang, Y.; Gonzalez-Ibeas, D.; Mayer, K.F.; Aranda, M.A.; Llave, C. Deep-sequencing of plant viral small RNAs reveals effective and widespread targeting of viral genomes. Virology 2009, 392, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Elbeaino, T.; Giampetruzzi, A.; De Stradis, A.; Digiaro, M. Deep-sequencing analysis of an apricot tree with vein clearing symptoms reveals the presence of a novel betaflexivirus. Virus Res. 2014, 181, 1–5. [Google Scholar] [CrossRef]
- Pecman, A.; Kutnjak, D.; Gutiérrez-Aguirre, I.; Adams, I.; Fox, A.; Boonham, N.; Ravnikar, M. Next Generation Sequencing for Detection and Discovery of Plant Viruses and Viroids: Comparison of Two Approaches. Front. Microbiol. 2017, 8, 1998. [Google Scholar] [CrossRef] [Green Version]
- Poojari, S.; Alabi, O.J.; Fofanov, V.Y.; Naidu, R.A. A Leafhopper-Transmissible DNA Virus with Novel Evolutionary Lineage in the Family Geminiviridae Implicated in Grapevine Redleaf Disease by Next-Generation Sequencing. PLoS ONE 2013, 8, e64194. [Google Scholar] [CrossRef] [PubMed]
- Rott, M.; Kesanakurti, P.; Berwarth, C.; Rast, H.; Boyes, I.; Phelan, J.; Jelkmann, W. Discovery of Negative-Sense RNA Viruses in Trees Infected with Apple Rubbery Wood Disease by Next-Generation Sequencing. Plant Dis. 2017, 102, 1254–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rott, M.; Xiang, Y.; Boyes, I.; Belton, M.; Saeed, H.; Kesanakurti, P.; Hayes, S.; Lawrence, T.; Birch, C.; Bhagwat, B.; et al. Application of Next Generation Sequencing for Diagnostic Testing of Tree Fruit Viruses and Viroids. Plant Dis. 2017, 101, 1489–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burd, E.M. Validation of laboratory-developed molecular assays for infectious diseases. Clin. Microbiol. Rev. 2010, 23, 550–576. [Google Scholar] [CrossRef] [Green Version]
- Puckett, J. The Davis grapevine virus collection—A current perspective. In Proceedings of the 19th Congress of the ICVG, Santiago, Chile, 9–12 April 2018. [Google Scholar]
- Golino, A. The Davis Grapevine Virus Collection. Am. J. Enol. Vitic. 1992, 43, 6. [Google Scholar]
- Diaz-Lara, A.; Stevens, K.; Klaassen, V.; Golino, D.; Al Rwahnih, M. Comprehensive Real-Time RT-PCR Assays for the Detection of Fifteen Viruses Infecting Prunus spp. Plants 2020, 9, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Rwahnih, M.; Rowhani, A.; Westrick, N.; Stevens, K.; Diaz-Lara, A.; Trouillas, F.P.; Preece, J.; Kallsen, C.; Farrar, K.; Golino, D. Discovery of Viruses and Virus-Like Pathogens in Pistachio using High-Throughput Sequencing. Plant Dis. 2018, 102, 1419–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesanakurti, P.; Belton, M.; Saeed, H.; Rast, H.; Boyes, I.; Rott, M. Screening for plant viruses by next generation sequencing using a modified double strand RNA extraction protocol with an internal amplification control. J. Virol. Methods 2016, 236, 35–40. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massart, S.; Chiumenti, M.; De Jonghe, K.; Glover, R.; Haegeman, A.; Koloniuk, I.; Komínek, P.; Kreuze, J.; Kutnjak, D.; Lotos, L.; et al. Virus Detection by High-Throughput Sequencing of Small RNAs: Large-Scale Performance Testing of Sequence Analysis Strategies. Phytopathology 2018, 109, 488–497. [Google Scholar] [CrossRef] [Green Version]
- EPPO PM 7/98 (4). Specific requirements for laboratories preparing accreditation for a plant pest diagnostic activity. EPPO Bull. 2019, 49, 530–563. [Google Scholar] [CrossRef] [Green Version]
- Al Rwahnih, M.; Daubert, S.; Úrbez-Torres, J.R.; Cordero, F.; Rowhani, A. Deep sequencing evidence from single grapevine plants reveals a virome dominated by mycoviruses. Arch. Virol. 2011, 156, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Chiaki, Y.; Ito, T. Complete genome sequence of a novel putative polerovirus detected in grapevine. Arch. Virol. 2020, 165, 1007–1010. [Google Scholar] [CrossRef]
- Rwahnih, M.A.; Alabi, O.J.; Hwang, M.S.; Stevens, K.; Golino, D. Identification and genomic characterization of grapevine Kizil Sapak virus, a novel grapevine-infecting member of the family Betaflexiviridae. Arch. Virol. 2019, 164, 3145–3149. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Bester, R.; Cook, G.; Breytenbach, J.H.J.; Steyn, C.; De Bruyn, R.; Maree, H.J. Towards the validation of high-throughput sequencing (HTS) for routine plant virus diagnostics: Measurement of variation linked to HTS detection of citrus viruses and viroids. Virol. J. 2021, 18, 61. [Google Scholar] [CrossRef] [PubMed]
- Kutnjak, D.; Tamisier, L.; Adams, I.; Boonham, N.; Candresse, T.; Chiumenti, M.; De Jonghe, K.; Kreuze, J.F.; Lefebvre, M.; Silva, G.; et al. A Primer on the Analysis of High-Throughput Sequencing Data for Detection of Plant Viruses. Microorganisms 2021, 9, 841. [Google Scholar] [CrossRef] [PubMed]
- Tamisier, L.; Haegeman, A.; Foucart, Y.; Fouillien, N.; Al Rwahnih, M.; Buzkan, N.; Candresse, T.; Chiumenti, M.; De Jonghe, K.; Lefebvre, M.; et al. Semi-artificial datasets as a resource for validation of bioinformatics pipelines for plant virus detection. PCI Genom. 2021, 1–15. [Google Scholar] [CrossRef]
- Gaafar, Y.Z.A.; Ziebell, H. Comparative study on three viral enrichment approaches based on RNA extraction for plant virus/viroid detection using high-throughput sequencing. PLoS ONE 2020, 15, e0237951. [Google Scholar] [CrossRef]
- Olmedo-Velarde, A.; Navarro, B.; Hu, J.S.; Melzer, M.J.; Di Serio, F. Novel Fig-Associated Viroid-Like RNAs Containing Hammerhead Ribozymes in Both Polarity Strands Identified by High-Throughput Sequencing. Front. Microbiol. 2020, 11, 2214. [Google Scholar] [CrossRef]
- Candresse, T.; Marais, A.; Faure, C.; Gentit, P. Association of Little cherry virus 1 (LChV1) with the Shirofugen Stunt Disease and Characterization of the Genome of a Divergent LChV1 Isolate. Phytopathology 2013, 103, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Weber, F.; Wagner, V.; Rasmussen, S.B.; Hartmann, R.; Paludan, S.R. Double-Stranded RNA Is Produced by Positive-Strand RNA Viruses and DNA Viruses but Not in Detectable Amounts by Negative-Strand RNA Viruses. J. Virol. 2006, 80, 5059–5064. [Google Scholar] [CrossRef] [Green Version]
- Dodds, J.A.; Morris, T.J.; Jordan, R.L. Plant Viral Double-Stranded RNA. Annu. Rev. Phytopathol. 1984, 22, 151–168. [Google Scholar] [CrossRef]
- Beuve, M.; Hily, J.-M.; Alliaume, A.; Reinbold, C.; Le Maguet, J.; Candresse, T.; Herrbach, E.; Lemaire, O. A complex virome unveiled by deep sequencing analysis of RNAs from a French Pinot Noir grapevine exhibiting strong leafroll symptoms. Arch. Virol. 2018, 163, 2937–2946. [Google Scholar] [CrossRef] [Green Version]
- Dubrovina, A.S.; Kiselev, K.V. Exogenous RNAs for Gene Regulation and Plant Resistance. Int. J. Mol. Sci. 2019, 20, 2282. [Google Scholar] [CrossRef] [Green Version]
- Monis, J.; Bestwick, R.K. Detection and Localization of Grapevine Leafroll Associated Closteroviruses in Greenhouse and Tissue Culture Grown Plants. Am. J. Enol. Vitic. 1996, 47, 199–205. [Google Scholar]
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Shabanian, M.; Xiao, H.; Meng, B. Seasonal dynamics and tissue distribution of two major viruses associated with grapevine Leafroll under cool climate condition. Eur. J. Plant Pathol. 2020, 158, 1017–1031. [Google Scholar] [CrossRef]
- Tsai, C.W.; Daugherty, M.P.; Almeida, R.P.P. Seasonal dynamics and virus translocation of Grapevine leafroll-associated virus 3 in grapevine cultivars. Plant Pathol. 2012, 61, 977–985. [Google Scholar] [CrossRef]
- Kominek, P.; Glasa, M.; Kominkova, M. Analysis of multiple virus-infected grapevine plant reveals persistence but uneven virus distribution. Acta Virol. 2009, 53, 281–285. [Google Scholar] [CrossRef]
- Čepin, U.; Gutiérrez-Aguirre, I.; Balažic, L.; Pompe-Novak, M.; Gruden, K.; Ravnikar, M. A one-step reverse transcription real-time PCR assay for the detection and quantitation of Grapevine fanleaf virus. J. Virol. Methods 2010, 170, 47–56. [Google Scholar] [CrossRef]
- Setiono, F.J.; Chatterjee, D.; Fuchs, M.; Perry, K.L.; Thompson, J.R. The Distribution and Detection of Grapevine red blotch virus in its Host Depend on Time of Sampling and Tissue Type. Plant Dis. 2018, 102, 2187–2193. [Google Scholar] [CrossRef] [Green Version]
- Al Rwahnih, M.; Saldarelli, P.; Rowhani, A. Grapevineleafroll-associated virus 7. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 221–228. ISBN 9783319577067. [Google Scholar]
- Burger, J.T.; Maree, H.J.; Gouveia, P.; Naidu, R.A. Grapevine leafroll-associated virus 3. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 167–195. ISBN 9783319577067. [Google Scholar]
- Pawitan, Y.; Michiels, S.; Koscielny, S.; Gusnanto, A.; Ploner, A. False discovery rate, sensitivity and sample size for microarray studies. Bioinformatics 2005, 21, 3017–3024. [Google Scholar] [CrossRef]
- Kircher, M.; Sawyer, S.; Meyer, M. Double indexing overcomes inaccuracies in multiplex sequencing on the Illumina platform. Nucleic Acids Res. 2012, 40, e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Plant ID | AGVd | ArMV | GAMaV | GBV-1 | GEV-1 | GFkV | GFLV | GKSV | GLRaV-1 | GLRaV-2 | GLRaV-3 | GLRaV-4 | GLRaV-7 | GPoV-1 | GRBV | GRGV | GRLDaV | GRSPaV | GRVFV | GVA | GVB | GVD | GVE | GVF | GVL | GYSVd-1 | GYSVd-2 | HSVd | satGVV |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | x | x | x | x | x | x | x | ||||||||||||||||||||||
| 2 | x | x | x | x | x | x | x | x | x | x | |||||||||||||||||||
| 3 | x | x | x | x | x | x | |||||||||||||||||||||||
| 4 | x | x | x | x | x | x | x | ||||||||||||||||||||||
| 5 | x | x | x | x | x | x | x | x | x | ||||||||||||||||||||
| 6 | x | x | x | x | x | x | x | x | x | x | x | x | |||||||||||||||||
| 7 | x | x | x | x | x | x | x | x | x | x | x | ||||||||||||||||||
| 8 | x | x | x | x | x | ||||||||||||||||||||||||
| 9 | x | x | x | x | x | x | x | x | x | x | x | ||||||||||||||||||
| 10 | x | x | x | x | x | x | x | x | |||||||||||||||||||||
| 11 | x | x | x | x | x | x | |||||||||||||||||||||||
| 12 | x | x | x | x | x | x | x | ||||||||||||||||||||||
| 13 | x | x | x | ||||||||||||||||||||||||||
| 14 | x | x | x | x | x | x | x | x | |||||||||||||||||||||
| 15 | x | x | x | x | x | x | x | x | |||||||||||||||||||||
| 16 | x | x | x | x | x | ||||||||||||||||||||||||
| 17 | x | x | x | x | |||||||||||||||||||||||||
| 18 | x | x | x | x | |||||||||||||||||||||||||
| HC | x |
| HTS Run | Tissue | No. VP a | No. VN b | Template | Spike-In |
|---|---|---|---|---|---|
| 1 | Cane | 18 | 1 | TNA | - |
| 2 | Petiole | 18 | 1 | TNA | BTS |
| 3 | Cane/Petiole | 9/9 | 0/1 | dsRNA | BTS |
| 4 | Cane/Petiole | 9/9 | 1/0 | dsRNA | BTS |
| HTS Run # | Tissue | Template | Spike-In | No. Samples per Run | Sample Description |
|---|---|---|---|---|---|
| 1 | Petiole | TNA | BTS | 18 | Sample set 1, Extraction 1 Sample set 1, Extraction 2 Sample set 2, Extraction 1 |
| 2 | Cane | TNA | BTS | 18 | Sample set 2,Extraction 2 Sample set 3, Extraction 1 Sample set 3, Extraction 2 |
| 3 | Petiole | TNA | BTS | 18 | Sample set 2, Extraction 2 Sample set 3, Extraction 1 Sample set 3, Extraction 2 |
| 4 | Cane | TNA | BTS | 18 | Sample set 1, Extraction 1 Sample set 1, Extraction 2 Sample set 2, Extraction 1 |
| Infection Dilution (%) | Detection Sensitivity (%) | Average | |||||
|---|---|---|---|---|---|---|---|
| Sample Set 1 | Sample Set 2 | Sample Set 3 | |||||
| Cane | Petiole | Cane | Petiole | Cane | Petiole | ||
| 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| 20 | 95 | 100 | 100 | 90 | 100 | 100 | 98 |
| 10 | 90 | 100 | 100 | 90 | 100 | 100 | 98 |
| 5 | 90 | 100 | 100 | 90 | 100 | 100 | 98 |
| 2 | 85 | 80 | 100 | 90 | 100 | 100 | 93 |
| 1 | 80 | 70 | 70 | 90 | 100 | 100 | 85 |
| Average | 90 | 92 | 95 | 92 | 100 | 100 | 95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltani, N.; Stevens, K.A.; Klaassen, V.; Hwang, M.-S.; Golino, D.A.; Al Rwahnih, M. Quality Assessment and Validation of High-Throughput Sequencing for Grapevine Virus Diagnostics. Viruses 2021, 13, 1130. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061130
Soltani N, Stevens KA, Klaassen V, Hwang M-S, Golino DA, Al Rwahnih M. Quality Assessment and Validation of High-Throughput Sequencing for Grapevine Virus Diagnostics. Viruses. 2021; 13(6):1130. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061130
Chicago/Turabian StyleSoltani, Nourolah, Kristian A. Stevens, Vicki Klaassen, Min-Sook Hwang, Deborah A. Golino, and Maher Al Rwahnih. 2021. "Quality Assessment and Validation of High-Throughput Sequencing for Grapevine Virus Diagnostics" Viruses 13, no. 6: 1130. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061130