1. Introduction
Influenza viruses cause acute respiratory tract infections that occur periodically in the form of epidemics and pandemics and remain a major unresolved public health problem worldwide. Annual influenza epidemics cause 3 to 5 million cases of severe respiratory diseases, up to 650,000 of which are fatal [
1]. Despite the decreased influenza activity in the 2020–2021 season due to the measures taken to combat the new coronavirus disease COVID-19 [
2], influenza viruses continue circulating in human and animal reservoirs with the potential to cause severe epidemics. Vaccination is by far the most effective measure to prevent the spread of influenza. However, seasonal vaccines induce primarily strain-specific immunity and are not effective against drifted seasonal or a new pandemic influenza virus. One of the possible ways to prevent epidemics caused by antigenically evolved viruses or pandemics caused by new influenza viruses could be the creation of a universal influenza vaccine, i.e., an influenza vaccine that has a broader protective effect than the seasonal vaccine [
3,
4]. One of the viral antigens which is most often used to design a universal influenza vaccine is an extracellular domain of matrix 2 protein (M2e) due to its high conservancy rate among all influenza A viruses. However, the M2e fragment itself is a weak immunogen due to its size, the small number of copies per virion, and also due to the shielding effect of the larger surface proteins of the influenza A virus. Therefore, multiple strategies have been proposed to increase the immunogenic and protective potential of this small protein [
5]. Previously, we have developed M2e-based universal influenza vaccine prototypes by genetic fusion of four M2e tandem repeats to the N-terminus of hemagglutinin (HA) subunit 1 and rescue of live attenuated influenza vaccine (LAIV) viruses based on a cold-adapted A/Leningrad/134/17/57 backbone. Mouse studies confirmed that these modified LAIVs were capable of inducing higher levels of M2e-specific antibodies compared to classical LAIVs, and these antibodies were associated with enhanced protection against a panel of heterologous and heterosubtypic influenza viruses [
6,
7]. To further optimize these vaccine candidates, we adjusted the consensus amino acid sequences of the M2e epitopes according to our comprehensive analysis of all influenza A viruses circulating in human and animal reservoirs [
5]. The new 4M2e construct consisted of four 23 amino acids-long M2e epitopes separated by flexible linkers, including consensus human (classical human, hM2e), swine (M2 of classical swine lineage, sM2e), avian/swine (M2 of swine influenza viruses derived from Eurasian avian H1N1, a/sM2e), and human/swine (M2 of pandemic H1N1, h/sM2e) M2 lineages. This construct was inserted in full-length HA or truncated NS1 genes of an H3N2 LAIV virus and two recombinant LAIV+4M2e viruses were assessed in a pre-clinical ferret model. Ferrets are considered the best test system for studying influenza in humans, as they are most susceptible to infection with virulent strains of influenza virus when administered intranasally and their infection proceeds as in humans with similar clinical symptoms [
8].
2. Materials and Methods
2.1. Cells, Viruses and Proteins
African green monkey kidney (Vero) cells were cultured in OptiPRO serum-free media supplemented with antibiotic-antimycotic and GlutaMAX (all from Thermo Fisher Scientific, Waltham, MA, USA) at 37 °C in the atmosphere of 5% CO2. Madin–Darby Canine Kidney (MDCK) cells were maintained in DMEM media with 10% fetal bovine serum (FBS) and 1x antibiotic-antimycotic (all from Thermo Fisher Scientific) at the same conditions as Vero cells.
An H3N2 LAIV reassortant virus carrying HA and NA genes of a recent H3N2 virus and six remaining genes of A/Leningrad/134/17/57 (H2N2) cold-adapted master donor virus was used as a viral vector for designing more broadly protective M2e-based vaccines. A set of dual-promoter plasmids encoding all genes of A/Leningrad/134/17/57 (H2N2) and A/Puerto Rico/8/34 (H1N1) viruses were described earlier [
9]. To rescue universal vaccine prototypes, as well as a control LAIV strain, wild-type HA and NA gene of A/Switzerland/9715293/2013 (H3N2) virus were cloned into pCIPolISapIT vector. To rescue a panel of recombinant PR8-based viruses, M genes of A/duck/Potsdam/1402-6/1986 (H5N2) and A/Aichi/2/68 (H3N2) viruses were also cloned into pCIPolISapIT vector by standard procedures. To rescue a PR8-based virus with M gene of swine-origin lineage, site-directed mutagenesis of A/Leningrad/134/17/57 was used to generate the consensus M2e sequence. This gene was also cloned into pCIPolISapIT vector to rescue the corresponding recombinant virus.
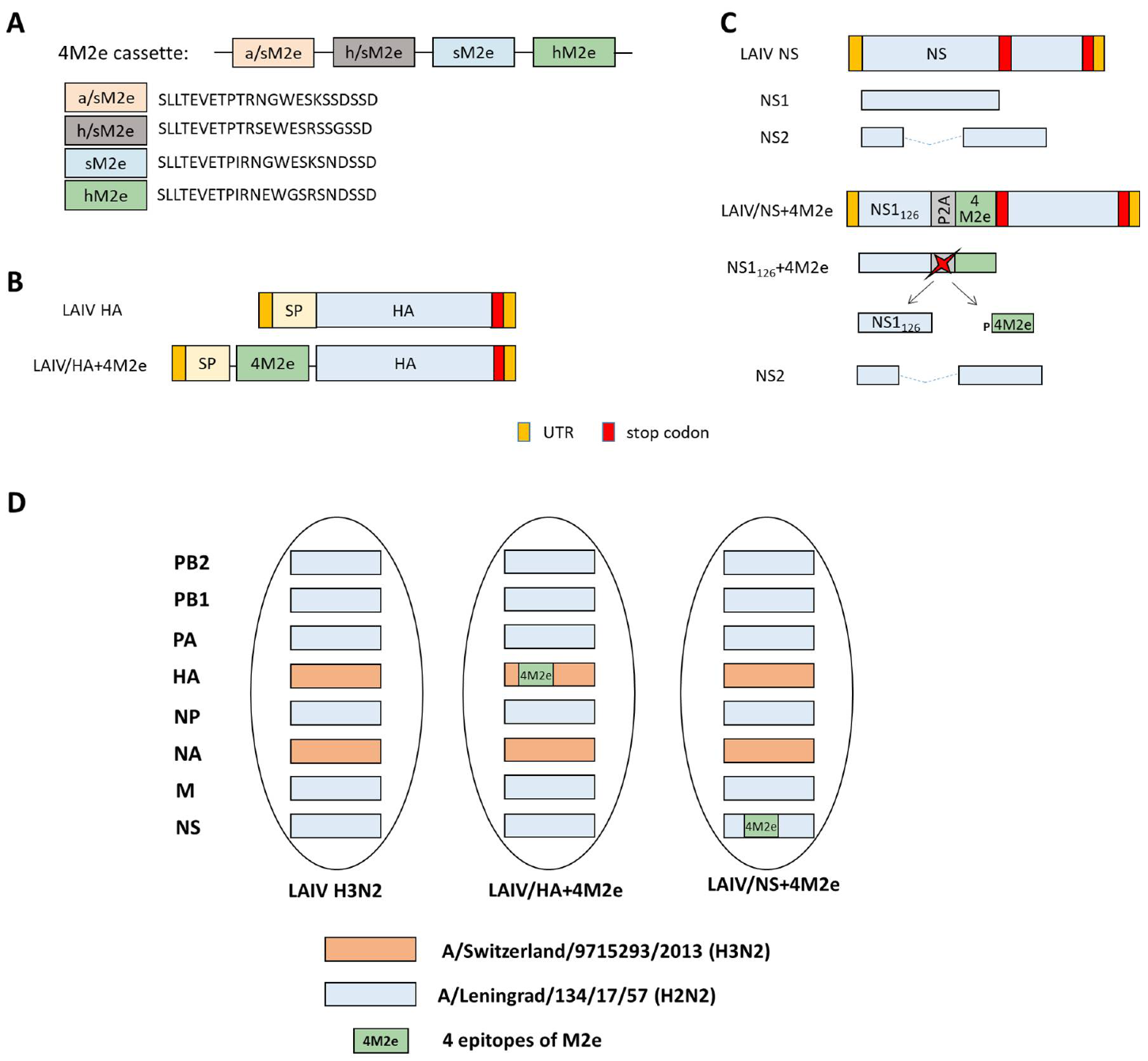
A DNA fragment encoding 4 M2e epitopes separated by flexible linkers (
Figure 1A) was chemically synthesized by Evrogen Ltd. (Moscow, Russia). The 4M2e construct was cloned from the N-terminus of the HA1 subunit and also in-frame of the NS1 gene truncated to 126 amino acids, via the P2A self-cleavage site, as previously described [
7,
10] (
Figure 1B,C). Briefly, the BsmBI restriction sites were inserted between the signal peptide and the HA1 subunit of the H3 HA molecule and at the residue NS1
126 of the A/Leningrad/134/17/57 NS gene using standard gene engineering approaches. Then, the 4M2e cassette was am amplified using sequence-specific primers with extended ends containing sites for the BsmBI restriction enzyme. Following restriction of the modified plasmids and the amplified insert with subsequent ligation resulted in generation of the HA+4M2e and NS+4M2e recombinant genes.
Two recombinant H3N2 LAIV viruses, LAIV/HA+4M2e and LAIV/NS+4M2e were rescued by electroporation of Vero cells using Neon transfection system (Thermo Fisher Scientific, USA), amplified in eggs and stored in single-use aliquots at −70 °C.
Influenza A virus A/South Africa/3626/2013 (pH1N1) (S.A. H1N1) was from the influenza virus repository of the Department of Virology, Institute of Experimental Medicine (Saint Petersburg, Russia). A mouse-adapted A/California/7/2009 (pH1N1) virus was obtained from the repository of respiratory viruses of Smorodintsev Research Institute of Influenza (Saint Petersburg, Russia).
A recombinant 3M2e protein expressed in
E. coli was kindly provided by Dr. Andris Kazaks (Latvian Biomedical Research and Study Centre, Riga, Latvia). The sequence of this protein was reported earlier [
11].
2.2. In Vitro Studies
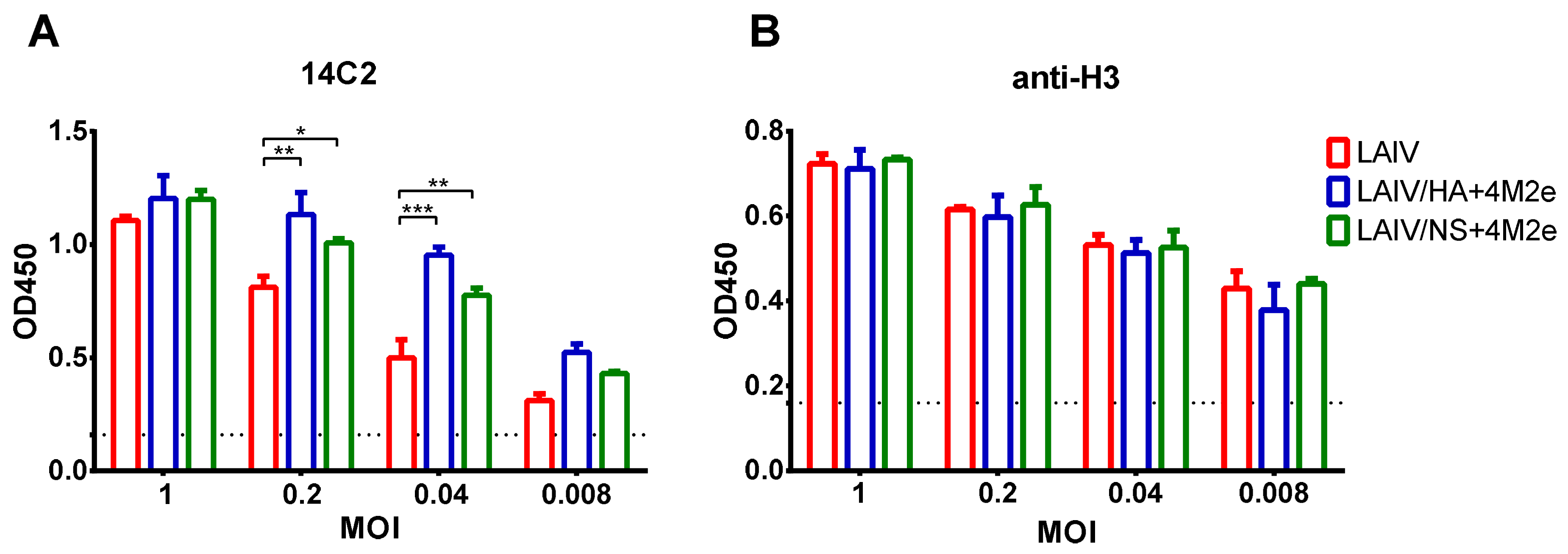
2.2.1. Expression of M2e Epitopes by Recombinant LAIV+4M2e Viruses
To assess the expression of M2e epitopes within an infected cell, MDCK cell monolayers were infected with studied viruses at various multiplicities of infection (MOI, number of TCID50 per cell): 1.0, 0.2, 0.04, and 0.008. The next day, the cells were fixed with 80% acetone, washed twice with PBS supplemented with 3% tween (PBST) and blocked with 5% non-fat dry milk in PBS. Then the cells were incubated with primary 14C2 antibody, followed by addition of secondary anti-mouse IgG antibody (Abcam, Cambridge, UK). The antibody binding was detected with 1-Step Ultra TMB-ELISA Substrate Solution (Thermo Fisher Scientific) and the color development was stopped by the addition of 1M H2SO4. The absorbance was measured at 450 nm using an xMark Microplate Spectrophotometer (BioRad, Hercules, CA, USA).
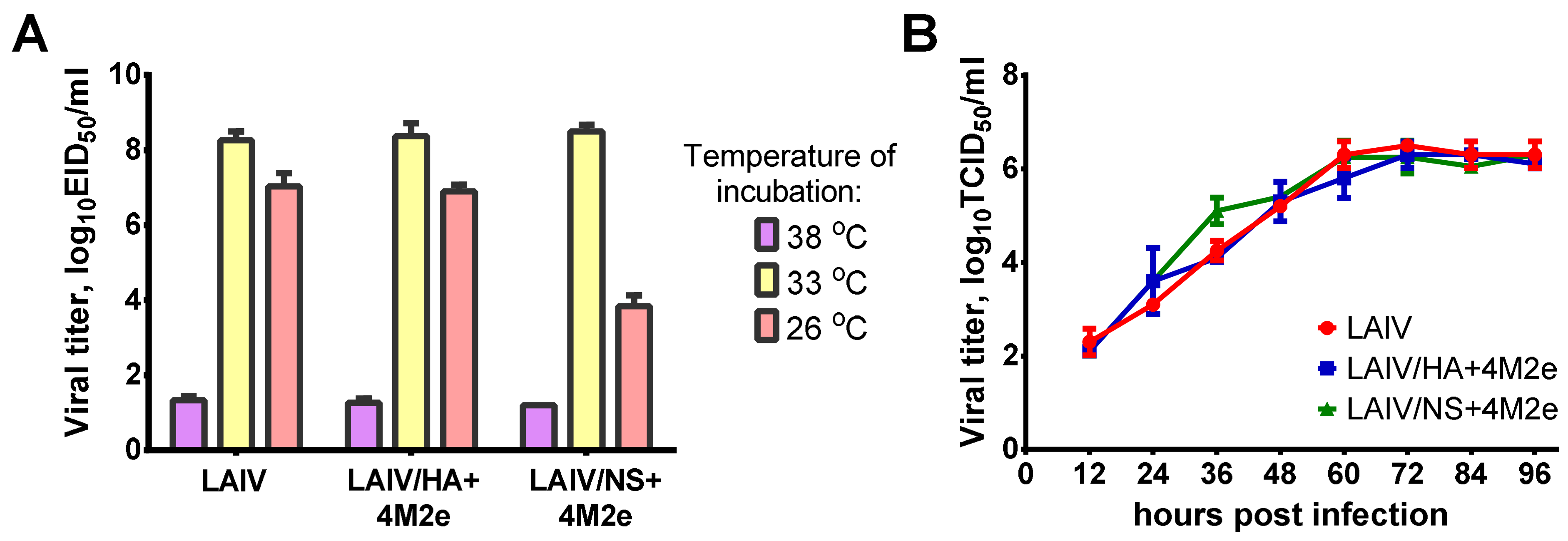
2.2.2. Growth Characteristics of LAIV Viruses
Growth characteristics of recombinant LAIV viruses were assessed in eggs and MDCK cells. Infectious titers in eggs were determined by end-point titration and titer calculation by the Reed and Muench method [
12] and expressed in log
10 50% egg infectious doses (log
10EID
50/mL). The virus was considered temperature-sensitive (ts) if the titer at 38 °C was reduced ≥5.0 log
10EID
50 compared to 33 °C. The virus was considered cold-adapted if the titer at 26 °C is reduced by ≤3.0 log
10EID
50 compared to 33 °C.
2.3. Animals, Immunization and Challenge
Male ferrets (aged 5–6 months and weighing 1.1–1.9 kg at the beginning of the experiment) were supplied by Scientific-Production Organization House of Pharmacy JSC (St. Petersburg, Russia). All animal experiments were approved by the Local Ethics Committee of the Institute of Preclinical Research Ltd., Saint Petersburg (Approval No. 4.51/20 from 11 September 2020).
All inoculations, nasal washes and blood sample collections were performed with the animal under short-term anesthesia induced by intramuscular injection of Zoletil 100, 12.5 mg/kg of body weight; every effort was made to minimize suffering. At the end of the study, animals were euthanized with an overdose of Zoletil-xylazine combination.
Before the study all ferrets were prescreened by hemagglutination inhibition test (HAI) to verify their negativity to circulating H1N1 and H3N2 human influenza viruses.
For the serum transfer experiments six to eight-week-old female C57BL/6J mice were purchased from Stolbovaya animal breeding nursery laboratory (Moscow region, Russia). Mice were anesthetized with isoflurane and all efforts were made to minimize animal suffering.
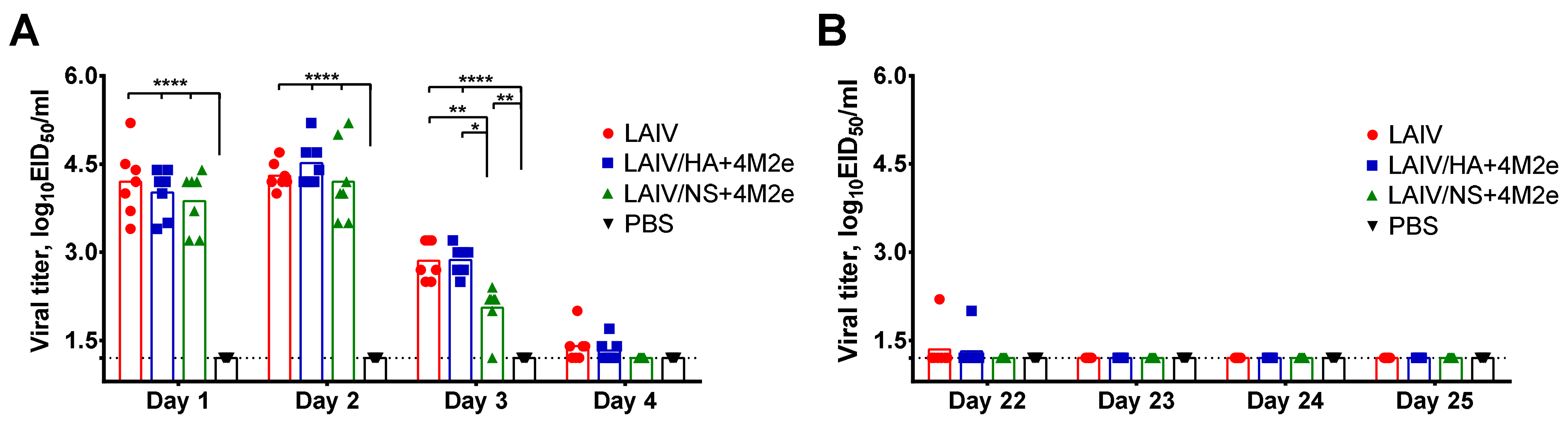
2.4. Replication of LAIV Viruses in the Upper Respiratory Tract
To determine the ability of the classical and recombinant LAIV viruses to replicate in the upper respiratory tract (URT) of immunized ferrets, nasal wash samples were collected from each animal on days 1, 2, 3 and 4 after each vaccine dose. 1 mL of sterile PBS was administered to one nostril of anesthetized ferret and the fluid excreting from the other nostril was collected using a petri dish. Ten-fold dilutions of the nasal wash (NW) fluids were inoculated into eggs and were incubated for 2 days at 33 °C and positive eggs were determined by HA assay using chicken red blood cells. Viral titers were calculated using Reed and Muench method and expressed in log10EID50/mL.
2.5. Safety Assessment
Assessment of the safety of LAIV candidates included several parameters, such as analysis of body weight and temperature of animals, data of clinical observations, as well as pathological examination of the respiratory tract of animals after immunization. Analysis of temperature, body weight of ferrets and clinical symptoms was carried out within 28 days of experiment, i.e., until day 7 after the second dose. At this time point, three animals per group were euthanized and subjected to autopsy. During autopsy, the external state of the body, chest and abdominal cavity with organs and tissues were examined in detail. Organ tissues (heart, trachea, lungs, liver, kidneys, adrenal glands, spleen, thymus, regional lymph node) were subjected to histological examination. The local irritant effect was assessed by histological analysis of the site of vaccine administration—the mucous membrane of the nasal cavity after decalcifying treatment. Tissues were fixed in a 10% buffered formalin solution and subjected to standard processing on an automatic Histo-Tek VP1 (Sakura, Alphen aan den Rijn, The Netherlands) histoprocessor with subsequent conclusion on Histomix medium (BioVitrum, St. Petersburg, Russia). Sections were made from paraffin blocks and were stained with hematoxylin and eosin, as well as with alcian blue dyeing for mucopolysaccharides (for the nasal cavity). Histological preparations were examined in a light microscope DM1000 (Leica, Wetzlar, Germany).
2.6. Assessment of Immune Responses
2.6.1. Hemagglutination Inhibition (HAI) Assay
HAI antibody titers in ferret sera were determined by the standard protocol described elsewhere [
13]. Briefly, ferrets’ sera were treated with receptor destroying enzyme (RDE, Sigma Aldrich, St. Louis, MO, USA) at 1:3 ratio and incubated for 16 h at 37 °C. Then, the treated samples were inactivated for 30 min at 56 °C. Serial 2-fold dilutions of each serum sample were incubated with four HA units of S.A. H1N1 and H3N2 LAIV viruses for 30 min and then mixed with 0.5% chicken red blood cells in the v-bottom 96-well microtiter plates. HAI titers were determined as the last serum dilution with complete inhibition of hemagglutination.
2.6.2. ELISA
Serum IgG antibody titers in ferret sera were determined on day 21 after each vaccine dose using enzyme-linked immunosorbent assay (ELISA). High-sorbent 96-well plates (Greiner Bio-One, Frickenhausen, Germany) were coated with 50 ng/well of either S.A. H1N1 or H3N2 LAIV sucrose-purified whole virus or recombinant 3M2e protein in a carbonate-bicarbonate buffer, in a volume of 50 µL per well at 4 °C overnight.
Two-fold dilutions of sera were prepared starting from 1:20 (for M2e antigen) or 1:200 (for H3N2 LAIV and S.A. H1N1 antigens) and added to the coated wells and incubated for 1 h at 37 °C, followed by incubation with anti-ferret IgG conjugated to horseradish peroxidase (Sigma). Antibody binding was detected with 1-Step Ultra TMB-ELISA Substrate Solution after incubation for 15 min at room temperature (Thermo Fisher Scientific). Optical density was measured at 450 nm using an xMark Microplate Spectrophotometer (BioRad). The area under the curve (AUC) of the OD450 values for all dilutions of each individual serum sample was calculated using the trapezoidal rule and expressed in arbitrary units.
2.6.3. Cell-Based ELISA
MDCK cells were used to assess the expression of M2e epitopes on the surface of infected cells, which were infected with 5-fold dilutions of LAIV, LAIV/HA+4M2e or LAIV/NS+4M2e, starting with 1 MOI (number of TCID50 per cell). As a control of equality of MOI doses of each virus H3N2 HA-specific antibody was used.
After 24 h of incubation, plates were fixed with cold 80% acetone and incubated on ice for 20 min. Then, the plates were washed twice with PBS supplemented with 3% tween (PBST) and blocked with 50 µL of 5% non-fat dry milk in PBS at 37 °C. After 1 h incubation, plates were washed with PBST and quenched with 50 µL of 0.8 M H2O2 at room temperature for 15 min. After an additional wash, primary 14C2 (1 µg/mL) or H3N2 HA-specific (1 µg/mL) antibodies diluted in 5% non-fat dry milk in PBS were added and plates were incubated at 37 °C for 1 h, followed by washing with PBST and addition of diluted 1:3000 secondary anti-mouse IgG antibody (Abcam, UK).
Afterwards, plates were washed 3 times, and antibody binding was detected with 1-Step Ultra TMB-ELISA Substrate Solution (Thermo Fisher Scientific). Once the desired color developed (approximately 15 min), 50 µL of H2SO4 stop solution were added to each well. The absorbance was measured at 450 nm using an xMark Microplate Spectrophotometer (BioRad).
2.6.4. Assessment of Cell-Mediated Immune Responses
Virus-specific and M2e-specific cellular responses were assessed by an intracellular cytokine staining (ICS) assay. For this, ferret splenocytes were isolated from small pieces of ferret spleen collected five days post-challenge using 70 µm cell strainers (Becton Dickinson, Franklin Lakes, NJ, USA). Red blood cells were lysed by an ammonium-chloride potassium lysing buffer (Thermo Fisher Scientific) and the single cell suspensions were maintained in CR-0 media (RPMI supplemented with 5 mM HEPES, 1× antibiotic-antimycotic, 50 µM β-mercaptoethanol, and 40 U/mL human IL-2). For
in vitro stimulation, 10
6 cells were incubated either with 1 MOI of sucrose-purified H3N2 LAIV and S.A. H1N1 virus or with 1 µg/well of 3M2e recombinant protein for 1 h, followed by addition of RPMI medium supplemented with 30% FBS to reach final serum concentration 10%. Antigen-stimulated cells were incubated at 37 °C, 5% CO
2 for 16 h, and then 50 μL of GolgiPlug solution (Becton Dickinson, USA) was added to stop protein transport. For positive assay control, a phorbol myristate acetate (PMA) was added to some wells, and the mixture was incubated for another five hours. After stimulation, the cells were stained with ZombieAqua fixable viability dye (BioLegend, San Diego, CA, USA), APC anti-CD4 (60003-MM02-A, Sino Biological, Beijing, China), and FITC anti-CD8 (ab 210326, Abcam) antibodies for 20 min in the dark. Then, the samples were washed twice with 200 μL of a staining buffer. For intracellular staining, a Cytofix/Cytoperm kit (Becton Dickinson, USA) was used according to the manufacturer’s instructions, followed by staining lymphocytes with a phycoerythrin (PE) anti-IFNγ (ab27866, Abcam) and Pacific Blue anti-CD3 (MCA1477PB, BioRad) antibodies for another 20 min. Then, the samples were washed twice with 200 μL of a wash buffer. The cells were fixed in 100 µL of Cyto-last buffer (BioLegend) and stored in a dark cool place prior to the flow cytometric analysis. At least 100,000 events were measured using a Navios flow cytometer (Beckman Coulter, Brea, CA, USA). The data were analyzed using the FlowJo software (TriStar Inc., El Segundo, CA, USA); the proportion of antigen-specific T cells was calculated by subtracting the negative control from the IFNγ-positive T cells. The gating strategy for the ICS assay is shown in
Figure S1.
2.6.5. Indirect Protective Effect of Immune Sera
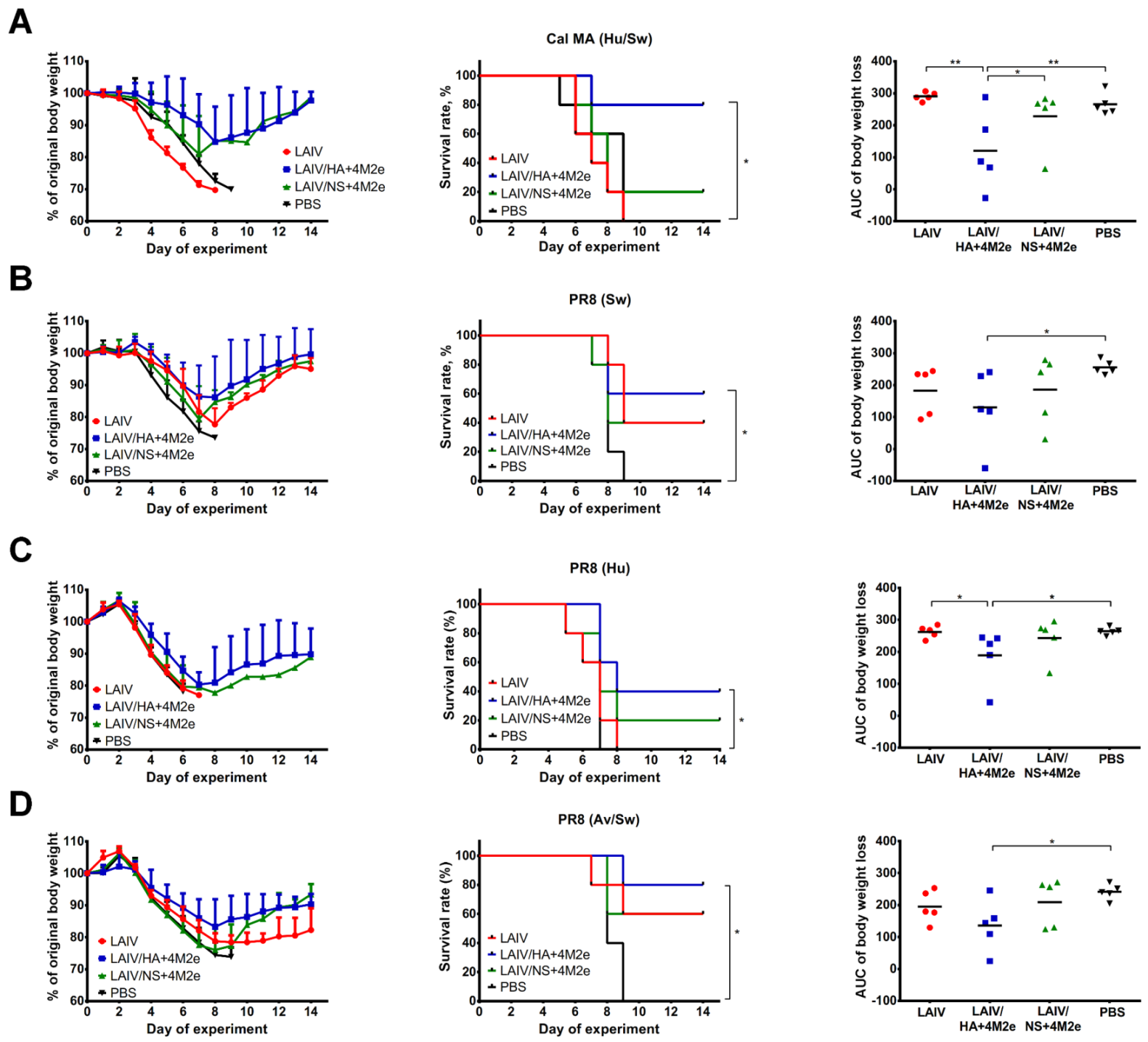
Indirect protective effect of antibodies in serum specimens collected on day 21 after the second vaccine dose was determined in a mouse in vivo protection model as previously described [
14]. Briefly, pooled sera from each vaccine group or placebo were mixed with PBS at a 1:1 ration and then heat-inactivated at 56 °C for 1 h. Then, the treated samples were mixed with 3 50% lethal doses (LD
50) of one of the following challenge viruses: (i) a mouse-adapted H1N1 A/California/7/2009—
Cal MA (Hu/Sw); (ii) a recombinant A/PR8-based virus carrying M gene of A/duck/Potsdam/1402-6/1986 (H5N2) virus—
PR8 (Av/Sw); (iii) a recombinant A/PR8-based virus carrying M gene of A/Aichi/2/1968 (H3N2) virus—
PR8 (Hu), and (iv) a recombinant A/PR8-based virus carrying artificial M gene of swine-origin lineage—
PR8 (Sw). The serum-virus mixtures were incubated at RT for 30 min and administered intranasally to naïve BALB/c mice in a volume of 50 μL. Protective effect of ferret sera was assessed by monitoring mouse survival rates and body weight loss during 14 days post-infection. Mice losing 30% of original body weight were humanely euthanized and recorded as dead.
2.6.6. Antibodies Produced by Lymphocytes in Mediastinal Lymph Nodes
Mediastinal lymph nodes (MLN) of ferrets were collected 5 days after the challenge. Single-cell suspension was prepared using 70 µm cell strainer (Becton Dickinson) and resuspended in 2 mL of CR-10 media (RPMI media supplemented with Ab/Am and 10% of FBS). After centrifugation at 300 RCF, 7 min, 22 °C and resuspension in 1 mL of CR-10, the MLN cells were counted. The isolated MLN cells were examined for their capacity to secrete both total IgG and antigen-specific IgG antibodies.
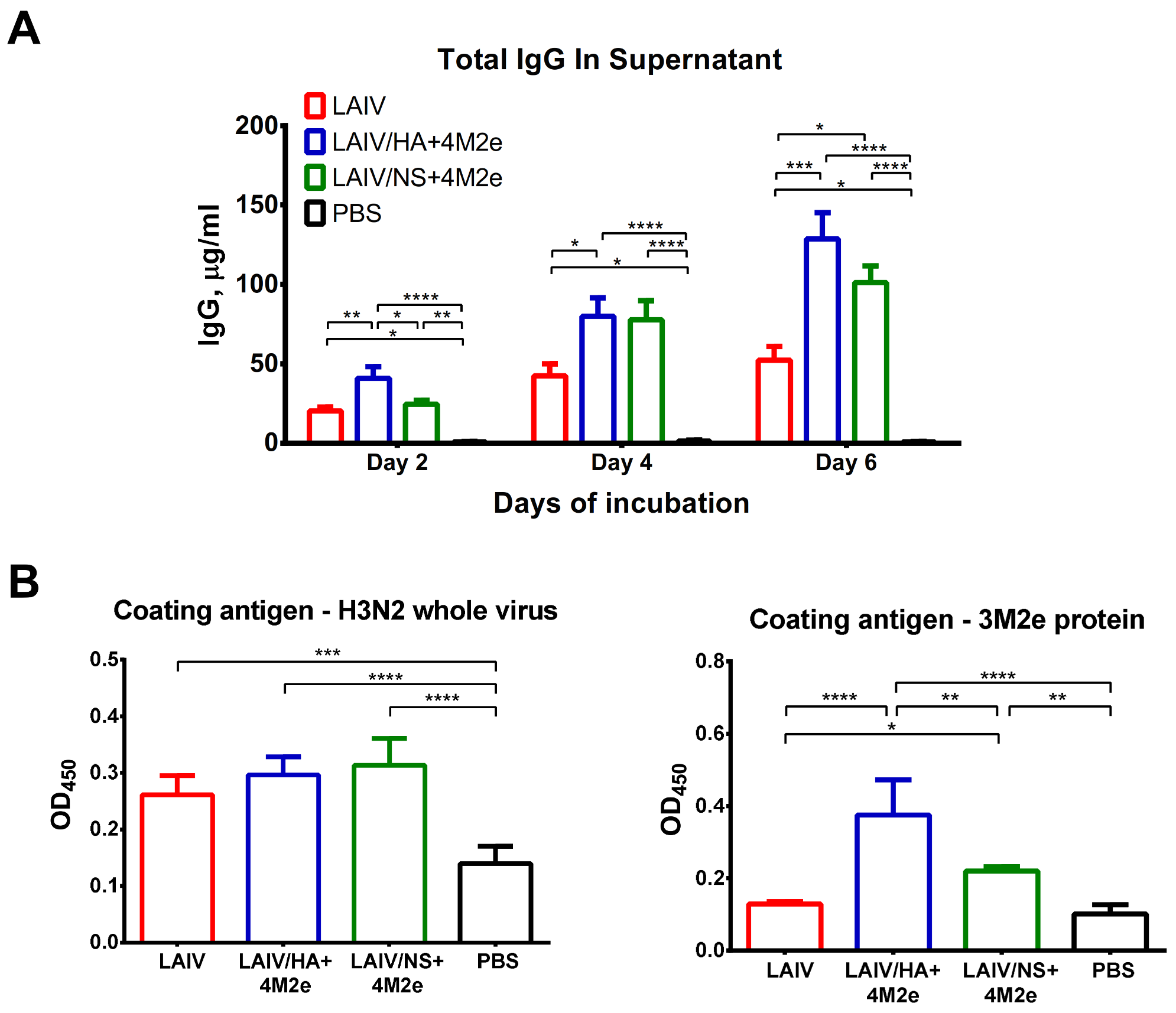
Total IgG in MLN
To determine the level of total IgG secreted by MLN lymphocytes, the isolated MLN cells from each ferret were resuspended in CR-0 media at 2 million cells per ml and incubated in non-binding 96-well plates in triplicates for 6 days at 37 °C, 5% CO2. The supernatants were collected on 2nd, 4th and 6th days of incubation (50 µL each) and kept at −20 °C until further processing. The concentration of total IgG in the samples was determined with a two-antibody sandwich ELISA. High-sorbent 96-well plates (Greiner Bio-One) were coated with a 100 ng/well of goat anti-ferret IgG (Sigma) diluted in PBS. Plates were incubated overnight at 4 °C, washed with PBST, and blocked with 1% of BSA in PBS for 30 min at 37 °C. Collected MLN supernatants were serially diluted starting from 1/10 and added to the wells of anti-ferret IgG-coated plates. Serial dilutions of purified ferret IgG native protein standard (2 mg/mL, Antibody Research Corporation, St. Charles, MO, USA) were added to the wells of each plate to generate standard curve. After 1 h incubation at 37 °C, all plates were washed with PBST and incubated with anti-ferret IgG conjugated to horseradish peroxidase (Sigma). The chromogen was produced using 1-Step Ultra TMB-ELISA Substrate Solution (Thermo Fisher Scientific) and measured at an absorbance of 450 nm. The concentration of total IgG was expressed in µg/mL from the regression curve of the standard ferret IgG titration.
Antigen-Specific IgG in MLN
Standard high-binding ELISA plates were coated with 50 ng/well of either H3N2 LAIV sucrose gradient purified whole virus or 3M2e protein at 4 °C overnight. The next day, the plates were washed with PBST and blocked with 1% BSA in PBS, and 106 of isolated MLN cells were added in triplicates to the ELISA plates, followed by incubation for 2 days at 37 °C, 5% CO2. Then, after three washes with PBST, all plates were incubated with anti-ferret IgG antibody conjugated to horseradish peroxidase (Sigma). Antibody binding was detected with 1-Step Ultra TMB-ELISA Substrate Solution (Thermo Fisher Scientific) after incubation for 15 min at room temperature and stopping with H2SO4. Optical density was measured at 450 nm using an xMark Microplate Spectrophotometer (BioRad).
2.7. Assessment of Protection against Heterologous Virus Challenge
Groups of four ferrets immunized with each tested vaccine or placebo were challenged with 106 log10EID50 of heterologous wild-type A/South Africa/3626/2013 (H1N1) virus on day 21 after the second vaccine dose. The virus was administered intranasally to anesthetized ferrets in a volume of 0.5 mL. The challenge phase lasted for five days, when the animals were euthanized for blood and tissue collection for further immunological, virological and/or histopathological analyses.
2.7.1. Protection by Clinical Outcomes
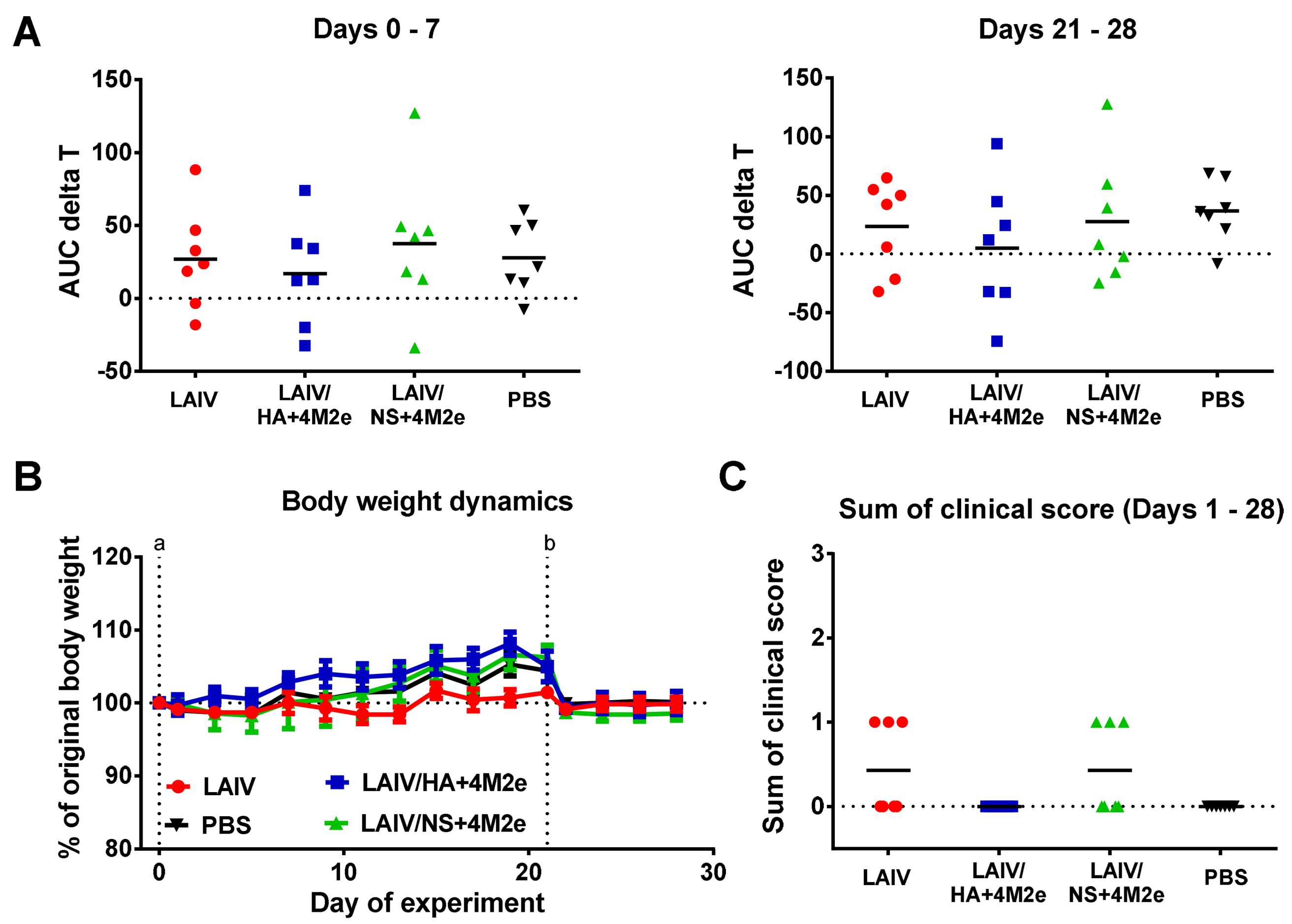
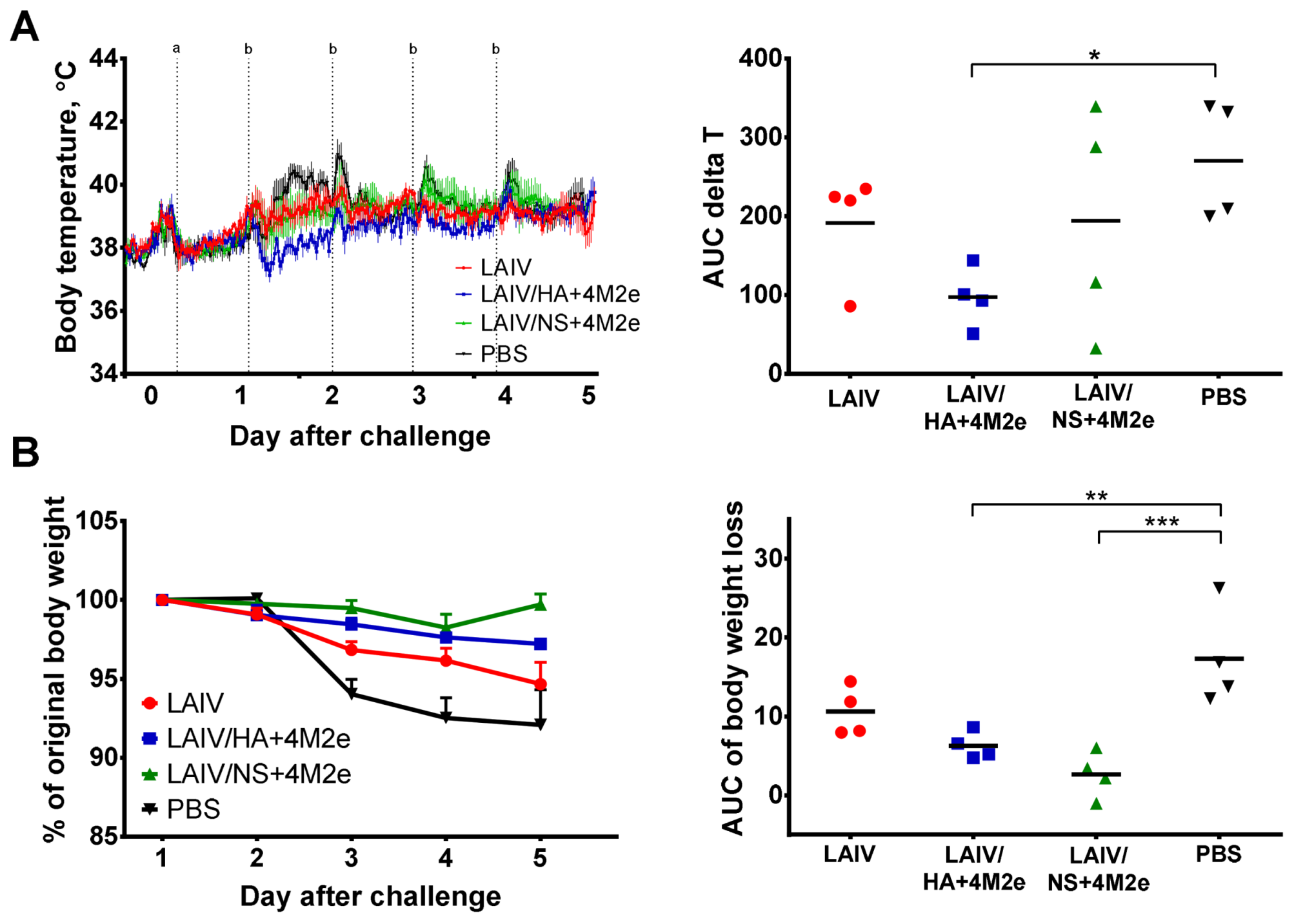
Body weight and clinical symptoms of disease (sneezing, nasal discharge and reduction in overall animal activity) were daily recorded for each animal. Temperature increase over the challenge phase was monitored by implanted temperature loggers that recorded body temperature every 30 min. The data from the temperature loggers was retrieved during the autopsy on day 47 of the experiment, and the temperature increase was calculated as an area under the curve (AUC) of the temperature data relative to the basal temperature previously determined for each animal (AUC delta T parameter). Body weight loss (% decrease relative to the weight on day 42) was also analyzed by the AUC parameter. In both cases the AUC data was calculated by the trapezoidal rule.
2.7.2. Protection by Virological Outcomes
Nasal wash specimens were collected daily until day five post challenge. Infectious viral titers were determined by end-point titration in eggs as described above and expressed as log10EID50/mL. In addition, viral titers were determined in nasal turbinates, trachea and lungs collected on day 5 after challenge. The lung tissues of each animal were harvested from all five lung lobes in separate 2 mL round-bottom tubes. Tissue samples were weighed and stored frozen at −70 °C until further processing. 1 mL of sterile cold PBS was added to each tissue sample and tissue homogenates were prepared using a small bead mill (TissueLyser LT, Qiagen, Hilden, Germany). Viral titers in the supernatants were determined by titration in eggs as described above and expressed as log10EID50/gram tissue.
2.7.3. Protection by Histopathological Outcomes
For histopathological evaluation of the damage to the lower respiratory tract, small pieces were excised from four lung lobes of each sacrificed animal and were placed in a 10% neutral buffered formalin solution for fixation. Then the samples were cut and placed into cassettes, followed by an automatic processing using a Tissue-Tek VP1 automated histoprocessor (Sakura). The resulting paraffin blocks were cut into sections with a thickness of 3 μm, which, after drying, were stained with hematoxylin and eosin. Histological preparations were examined under a Leica DM1000 microscope at 400× magnification. For each lung lobe, two sections were placed in a glass slide and all samples (32 per study group, 128 in total) were analyzed by a blinded histologist. The decrease in the airiness of the pulmonary lobes’ parenchyma was treated as an affective tissue. The area of affected lung tissue in % of all field area was measures by a microscope at 200× magnification with a calibrated objectives micrometer taking into account 15–40% compression of wet tissues due to fixation and sample histoprocessing [
15]. Histological evaluation of tissue injury also included the assessment of alveolar septum thickening by a microscope with calibrated objectives [
16].
2.8. Statistical Analyses
Data were analyzed with the statistical module of Graph Pad Prism 6 software (GraphPad Software, San Diego, CA, USA). Statistically significant differences between study groups were determined by ANOVA with Tukey’s multiple comparison test (in vitro and in vivo studies). Differences in the survival rates after challenge were analyzed by a log-rank Mantel–Cox test. p values of <0.05 were considered significant.
4. Discussion
Traditional influenza vaccines have been shown to substantially reduce the burden of influenza among various populations [
19]. However, these vaccines mainly induce neutralizing antibodies to viral epitopes that are mutation-prone, and the virus easily escapes the vaccine-induced immunity. To overcome the issue of constant antigenic drift, the composition of influenza vaccines needs to be updated frequently, and annual vaccination is recommended to maintain a sufficient level of immunity. However, a recent systematic review and meta-analysis revealed the negative impact of repeated vaccination on the effectiveness of licensed influenza vaccines [
20]. Furthermore, a wide range of natural hosts and different mutation pathways do not yet allow predicting a future circulating strain, despite significant progress in this area [
21]. All these problems explain the need to develop universal influenza vaccines that would induce broadly reactive and durable immunity, thereby protecting vaccinated people against the evolved seasonal influenza viruses, as well as against new pandemic variants [
22].
A variety of strategies have been explored to redirect the vaccine-induced immune responses from variable to more conserved protein targets, and the M2e epitope is one of the most commonly used target antigens in these studies. To increase the immunogenicity of this small protein, different carriers are being used, and several successful prototypes have reached clinical phase of development (reviewed in [
5,
23,
24]). In our early proof-of-concept study, we demonstrated that cold-adapted Leningrad/17-based LAIV viruses can serve as viral vectors to deliver tandem M2e repeats to target cells and stimulate the induction of high levels of M2e-specific antibodies that improve cross-protective potential of the vaccine [
6,
7]. In general, most of the studies of M2e-based universal vaccine candidates are limited to their assessment in a mouse model of infection. There are only a few published studies where such vaccines were assessed in other animals (ferrets, monkeys, pigs, dogs) [
25,
26,
27,
28]. In the current study, we evaluated two universal LAIV prototypes in ferrets, genetically outbred animals, which are considered the best test system for studying influenza in humans, since they have distinct patterns of T-cell reactivity as a result of heterogeneity at the MHC locus, and are also susceptible to infection with virulent influenza viruses with similar to humans manifestation of the disease [
8].
The two universal LAIV+4M2e candidates differed by the site of insertion of the 4M2e cassette. The first recombinant virus expressed the 4M2e foreign antigen from the N-terminus of the H3 HA1 subunit, similar to the previously reported H1N1 and H7N9 LAIV+4M2e candidates studied in mice [
6,
7]. Since this strategy will require regular update of the corresponding HA molecule to match the antigenic properties of circulating seasonal influenza viruses, we created another recombinant virus in which the 4M2e cassette is encoded in the open reading frame of the NS1 protein, which, in turn has been truncated to 126 residues. Since the NS gene originates from the Leningrad/17 master donor virus, there is no need to update this gene when seasonal viruses evolve. Here, the transgene is not present in the viral particle, but should be translated within the infected cell, because the NS1 protein is abundantly expressed in the early stages of infection [
29]. Such modification of LAIV genome has been previously shown as a promising strategy for the induction of robust cytotoxic T-cell responses to the inserted immunodominant CD8 T-cell epitope of a foreign pathogen [
30,
31,
32]. In vitro studies confirmed that regardless of how the 4M2e cassette was inserted into the LAIV genome, M2e proteins were expressed at significantly higher levels when mammalian cells were infected with the LAIV+4M2e recombinant candidates than with the classic LAIV strain, demonstrating the LAIV’s ability to properly deliver additional M2e antigens to target cells. It is important that the inserted 4M2e cassette did not disturb the replicative properties of the LAIV virus: both chimeric vaccines grew to comparable titers in eggs and MDCK cells, as well as in the upper respiratory tract of ferrets. Of note, our previous studies of NS1-modified recombinant LAIV viruses carrying immunogenic epitopes of other respiratory viruses demonstrated their inability to replicate in the mouse respiratory tract, suggesting that modulation of the LAIV virus growth properties by modification of NS1 protein is host-specific [
31,
32]. Importantly, all LAIVs studied in this experiment were safe and did not induce any infection-related pathology in immunized ferrets.
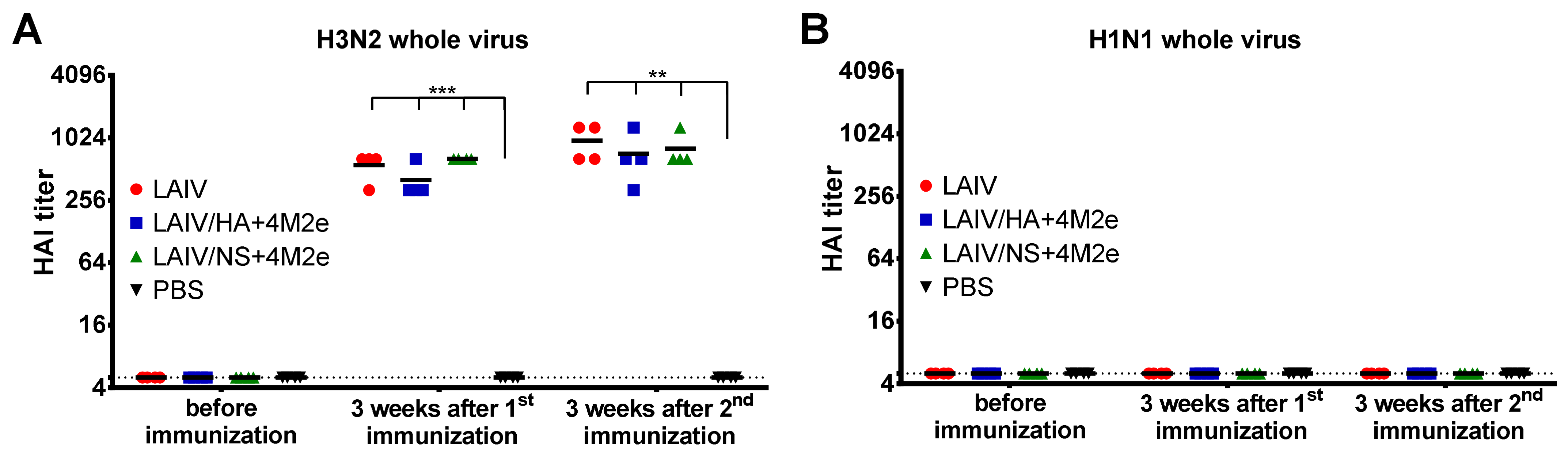
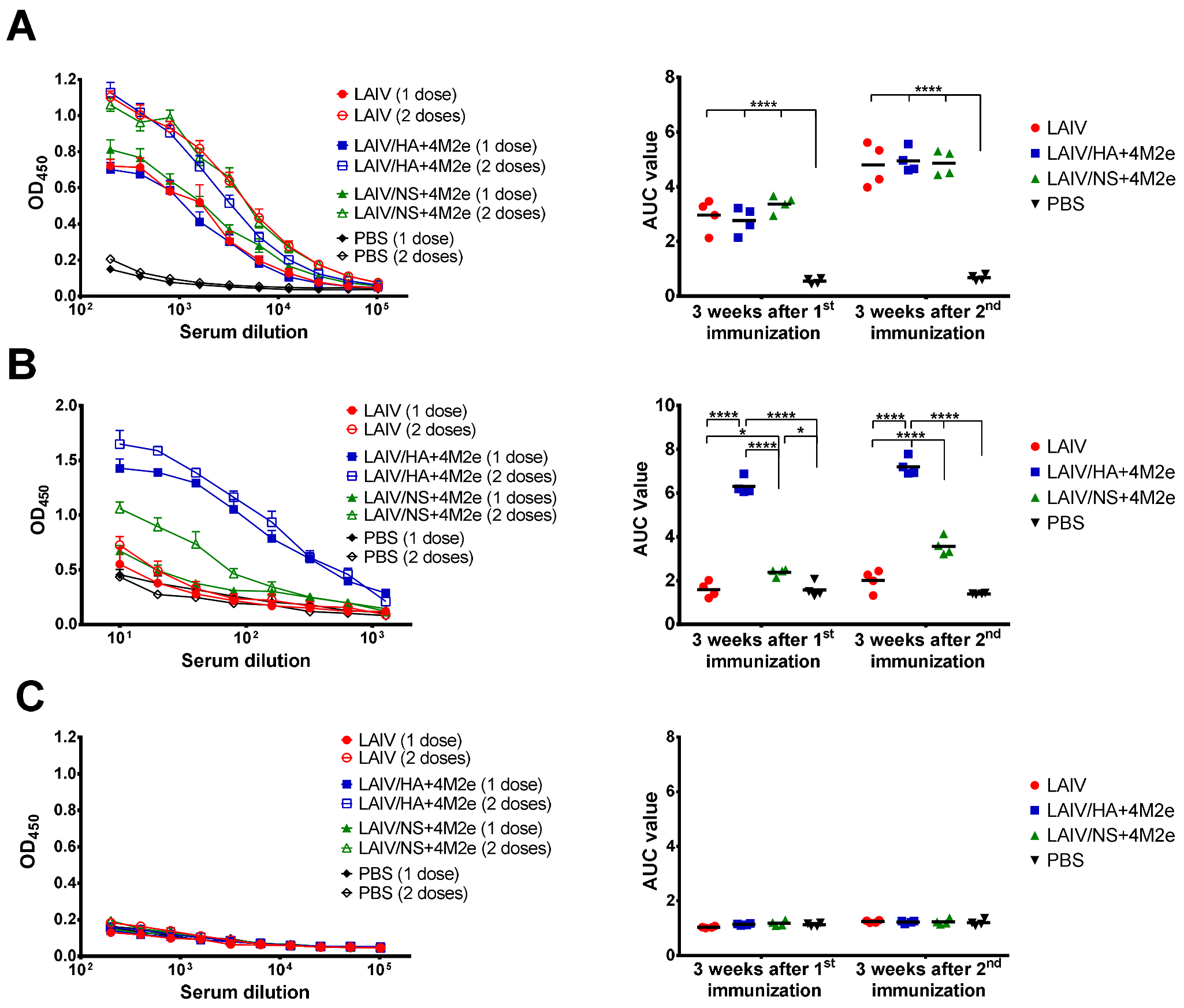
As expected, active replication of all three LAIV viruses promoted the development of a potent humoral immune response to the whole virus antigen, with significant boosting effect of the second dose. Importantly, there was no reduction in homologous HAI or serum IgG titers in ferrets immunized with recombinant LAIV+4M2e candidates, relative to the traditional LAIV virus, suggesting that none of the genetic modifications had an impact of influenza virus-specific humoral immunity. In line with previously published data, immunization with unmodified influenza virus (i.e., H3N2 LAIV) did not elicit notable levels of M2e-binding antibodies in ferrets [
33]. However, the insertion of the 4M2e cassette into LAIV genome significantly increased the M2e-specific antibody levels, with the LAIV/HA+4M2e variant being remarkably more immunogenic than the LAIV/NS+4M2e counterpart. Moreover, a single dose of the HA-modified vaccine candidate was sufficient for eliciting powerful M2e response, whereas at least two doses of the NS1-modified vaccine was required for induction of significant M2e-specific antibody levels. These data indicate that distinct modes of expression of the 4M2e insert from the two viral proteins result in different pathways for antigen presentation, thus resulting in such a divergent magnitude of the response. To date, a fairly large amount of data has been accumulated regarding the development of viral-vectored vaccines based on the influenza virus, where the transgene was inserted into the NS1 open reading frame. In most strategies, the gene of interest is either fused to a truncated NS1 protein or inserted via a stop-start cassette, and only a few studies report transgene insertion using the 2A auto-cleavage site (reviewed in [
34]). However, whether the insertion of 4M2e cassette into NS1 using a different cloning strategy will improve the immunogenicity of the M2e epitopes is unknown and requires further research.
The main objective of this study was the assessment of cross-protective potential of the new universal influenza vaccine candidates in the ferret model. For this purpose, the immunized ferrets were challenged with the S.A. H1N1 virus, which is known to cause significant pathology in this animal model [
18,
35]. Unlike H3N2 vaccine viruses, this subtype belongs to the Group 1 hemagglutinin, and no cross-reactive IgG responses to the challenge virus were found in ferret immune sera. It is known that classic LAIVs have a potential to protect against heterologous influenza viruses due to the induction of T-cell immune responses, as well as mucosal IgA antibody [
36,
37,
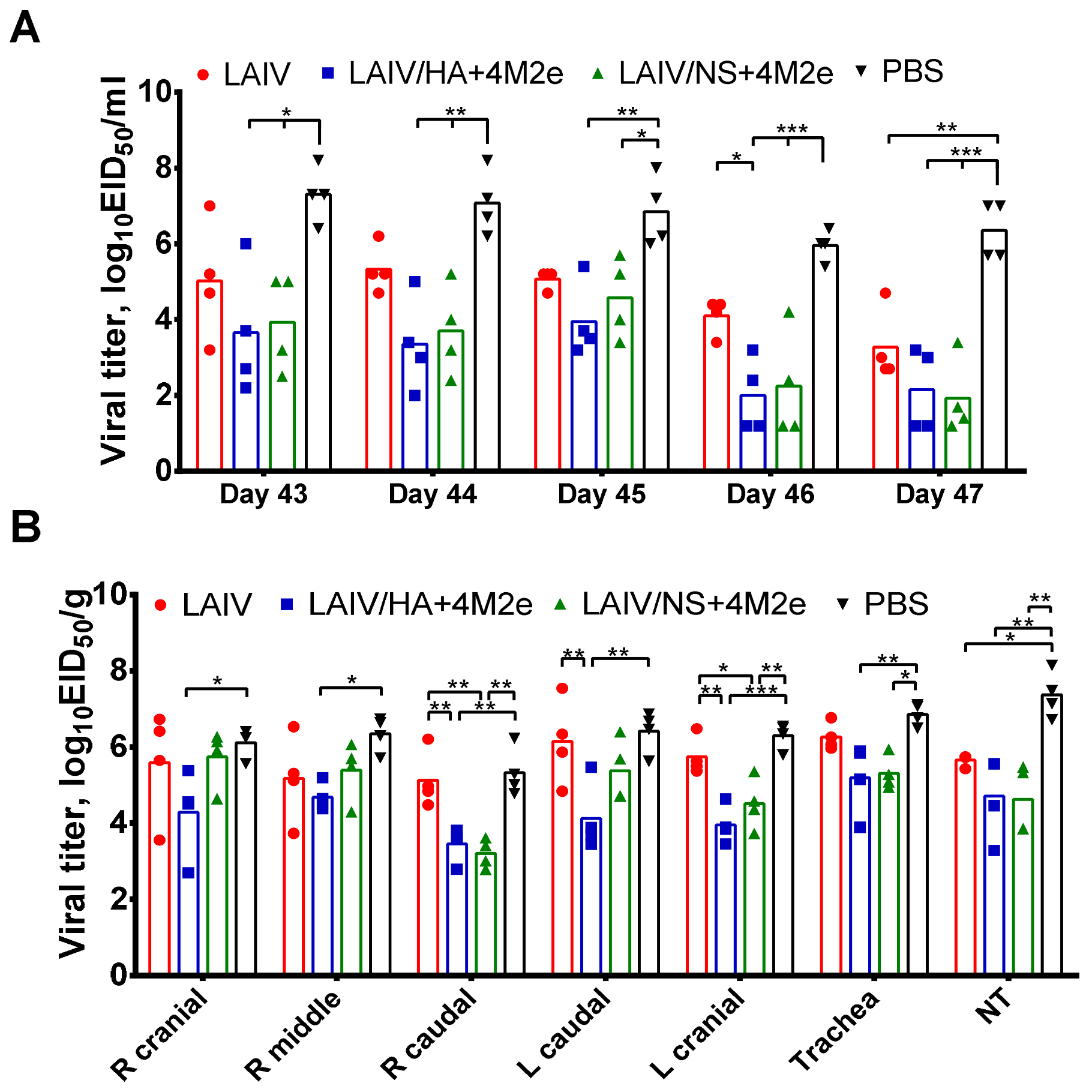
38]. Indeed, on day 5 post-challenge, there was a decrease in upper respiratory viral load in the LAIV group compared to the PBS group. In addition, LAIV-immunized ferrets showed less severe signs of lung pathology than control animals. However, the LAIV itself was unable to protect ferrets against H1N1 virus pulmonary replication and body weight loss, whereas both recombinant LAIV+4M2e vaccines induced sufficient immunity to reduce viral titers in all tested respiratory tissues.
Importantly, since the two universal LAIV candidates induced comparable humoral responses to whole influenza virus but were differently immunogenic in terms of inducing M2e-specific antibodies, it became possible to investigate the cross-protective potential of the M2e antibodies. Thus, ferrets immunized with LAIV/HA+4M2e were better protected against viral replication and clinical manifestation of the disease than the LAIV/NS+4M2e-vaccinated animals. Subsequent analysis revealed a strong correlation between virus titers in the respiratory tract and the M2e antibody levels, but not H3N2, prior to challenge. Furthermore, the protection was strongly associated with the levels of M2e-specific antibodies secreted by the MLN cells five days post-challenge. These data suggest that protection against a heterologous influenza virus infection was mediated by recall M2e-specific IgG responses in draining lymph nodes. Similar results were observed in a mouse model, where the improved cross-protection afforded by a recombinant H3N2 influenza virus expressing the 4M2e cassette in a chimeric hemagglutinin was associated with significant increases of M2e specific IgG levels in sera and MLN after challenge, if compared to the control vaccine virus [
39].
The limitation of the study is that we did not assess functional activity of the vaccine-induced M2e antibodies It is supposed that the M2e antibody can mediate protection via the antibody-dependent cellular cytotoxicity (ADCC) or complement-dependent cytotoxicity mechanisms [
40,
41,
42], however direct measurement of ADCC activity in ferrets is challenging due to the lack of ferret-specific reagents [
43]. Another limitation is that we did not assess the levels of mucosal IgA responses after vaccination. It is known that the nasal IgA antibody can play a role in reducing the load of challenge virus in the URT, since IgA antibody are naturally cross-neutralizing [
44]. However, in the current study, the decrease in viral titers in the URT did not correlate with the virus clearance from the lungs, suggesting that the sIgA antibody were not the main immune factors involved in the enhanced cross-protection. Nevertheless, the mucosal IgA and T-cell immune responses induced after LAIV immunization should also contribute to the overall protective potential of the vaccine.
It is interesting to compare the protective effect of two different universal LAIV strategies. We previously developed a panel of reassortant LAIV strains expressing stem-based chimeric HA molecules and evaluated them in sequential immunization regimens in ferrets [
18]. Both ferret studies used the same S.A. H1N1 challenge virus, which made it possible to compare the protective effect of the induced immune responses. Strikingly, the levels of stalk-HA-reactive antibodies induced by a three-dose sequential immunization with cHA-based LAIVs were not sufficient to significantly reduced viral titers and lung pathology after the H1N1 challenge, and the inclusion of a NP gene from the H1N1 virus into LAIV genome was required to achieve statistically significant reduction in viral pulmonary titers and clinical signs of disease [
18]. In contrast, the two-dose immunization with the LAIV+4M2e candidates induced strong M2e-specific antibody responses with cross-protective potential. One possible explanation for these discrepant results is that the cHA-based LAIVs replicated poorly in the ferrets’ upper respiratory tract, whereas the M2e-based LAIV candidates were highly infective and animals shed the vaccine viruses up to day 4 post inoculation, and such enhanced replicative activity could promote robust M2e-specific antibody responses. Another possibility is that the stalk-HA-targeted and the M2e antibodies differ by their avidity and Fc-mediated functional activities, however a side-by-side experiment is required to confirm this assumption.
Besides the induction of balanced adaptive immune responses, the M2e-based universal vaccine based on a licensed LAIV backbone has more advantages over other M2e-based approaches, such as an adjuvant-free technology and an intranasal route of administration, whereas the majority of other M2e-based vaccine prototypes are injectable and require adjuvants to boost potent M2e-specific antibody responses [
5]. Recent advances in influenza reverse genetics technology allow rapid generation of recombinant LAIV reassortant viruses with updated major antigenic determinants, including the design of the chimeric HA+4M2e gene to match currently circulating seasonal influenza viruses. However, if vaccine-induced broad immunity is to be sustained, regular antigen update will not be required. Although the current study did not assess the duration of the induced immune responses, a recent cHA-based study of LAIV candidates revealed the persistence of HA stem-reactive antibodies in immunized ferrets for at least one year [
45].


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










