
Characterization of Anti-Bacterial Effect of the Two New Phages against Uropathogenic Escherichia coli

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Molecular Characterization of New Bacteriophages against Uropathogenic Escherichia coli Strains
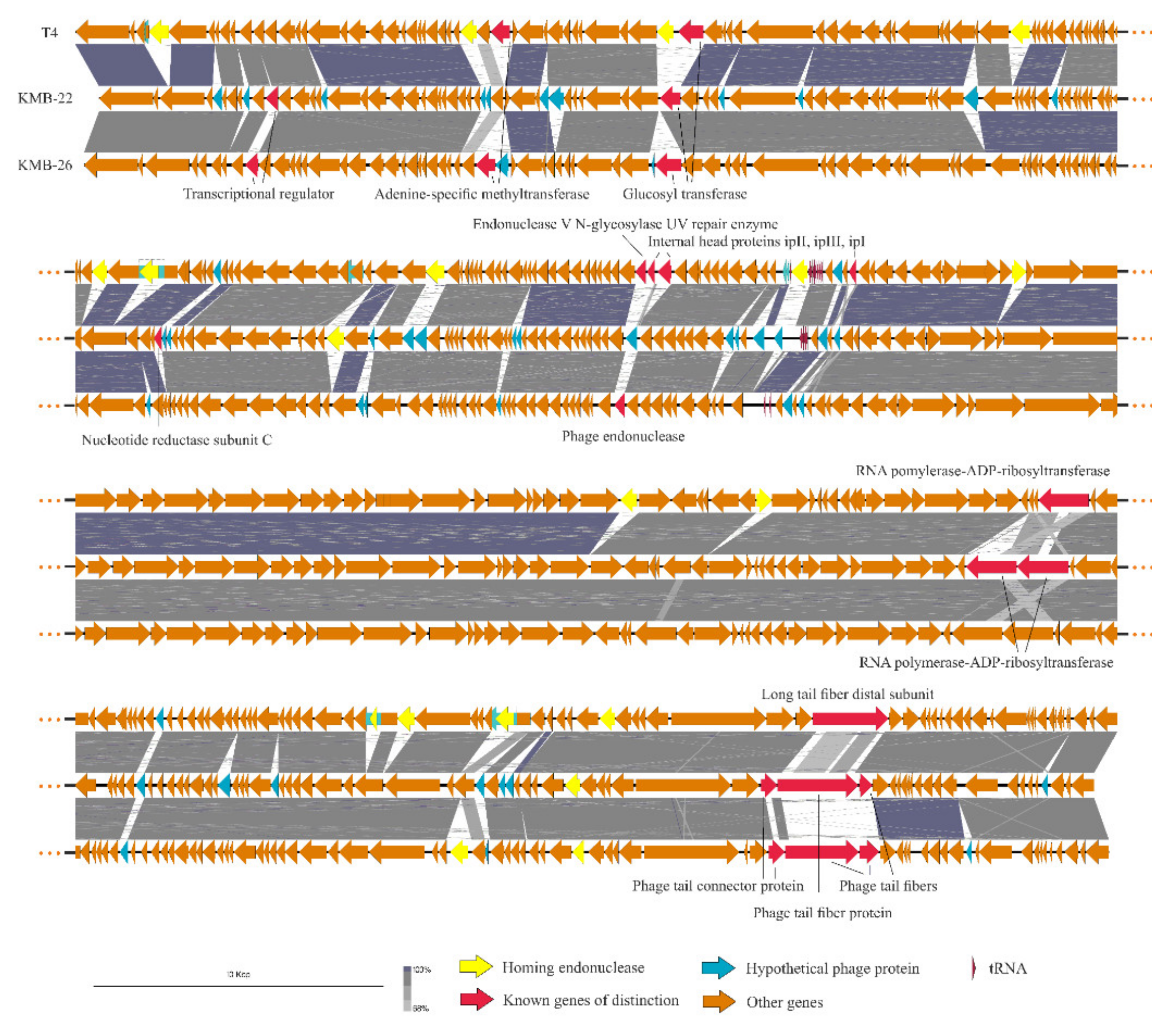
2.2. Isolation of Phage DNA, Sequencing and Bioinformatics
2.3. Clinical Escherichia coli Strains
2.4. Detection of Phage Antibacterial Activity
3. Results
3.1. Newly Isolated Anti-Escherichia coli Phages
3.2. Characteristics of Clinical Escherichia coli Strains Selected for Phage-Susceptibility Testing
3.3. Antibacterial Activity of the Phages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary Tract Infections: Epidemiology, Mechanisms of Infection and Treatment Options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Jhang, J.-F.; Kuo, H.-C. Recent Advances in Recurrent Urinary Tract Infection from Pathogenesis and Biomarkers to Prevention. Ci Ji Yi Xue Za Zhi 2017, 29, 131–137. [Google Scholar]
- Koreň, J. Urinary tract infections. In Medical Microbiology, 1st ed.; Liptáková, A., Ed.; Herba: Bratislava, Slovak Republic, 2019; pp. 799–805. (In Slovak) [Google Scholar]
- Lee, D.S.; Lee, S.-J.; Choe, H.-S. Community-Acquired Urinary Tract Infection by Escherichia coli in the Era of Antibiotic Resistance. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage Treatment of Human Infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCallin, S.; Alam Sarker, S.; Barretto, C.; Sultana, S.; Berger, B.; Huq, S.; Krause, L.; Bibiloni, R.; Schmitt, B.; Reuteler, G.; et al. Safety Analysis of a Russian Phage Cocktail: From Metagenomic Analysis to Oral Application in Healthy Human Subjects. Virology 2013, 443, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Llorente, C.; Lang, S.; Brandl, K.; Chu, H.; Jiang, L.; White, R.C.; Clarke, T.H.; Nguyen, K.; Torralba, M.; et al. Bacteriophage Targeting of Gut Bacterium Attenuates Alcoholic Liver Disease. Nature 2019, 575, 505–511. [Google Scholar] [CrossRef]
- Bao, J.; Wu, N.; Zeng, Y.; Chen, L.; Li, L.; Yang, L.; Zhang, Y.; Guo, M.; Li, L.; Li, J.; et al. Non-Active Antibiotic and Bacteriophage Synergism to Successfully Treat Recurrent Urinary Tract Infection Caused by Extensively Drug-Resistant Klebsiella pneumoniae. Emerg. Microbes Infect. 2020, 9, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Barr, J.J.; Auro, R.; Furlan, M.; Whiteson, K.L.; Erb, M.L.; Pogliano, J.; Stotland, A.; Wolkowicz, R.; Cutting, A.S.; Doran, K.S.; et al. Bacteriophage Adhering to Mucus Provide a Non-Host-Derived Immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 10771–10776. [Google Scholar] [CrossRef] [Green Version]
- Garretto, A.; Miller-Ensminger, T.; Wolfe, A.J.; Putonti, C. Bacteriophages of the Lower Urinary Tract. Nat. Rev. Urol. 2019, 16, 422–432. [Google Scholar] [CrossRef]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S. Phage Therapy in Clinical Practice: Treatment of Human Infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the All-Bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, J.H.; Carroll, K.C.; Funke, G.; Pfaller, M.A.; Landry, M.L.; Richter, S.S.; Warnock, D.W. (Eds.) Manual of Clinical Microbiology, 11th ed.; ASM Press: Washington, DC, USA, 2015. [Google Scholar]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli Phylo-Typing Method Revisited: Improvement of Specificity and Detection of New Phylo-Groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Besser, T.E.; Call, D.R.; Weissman, S.J.; Jones, L.P.; Davis, M.A. Evaluation of Two Multi-Locus Sequence Typing Schemes for Commensal Escherichia coli from Dairy Cattle in Washington State. J. Microbiol. Methods 2016, 124, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grose, J.H.; Casjens, S.R. Understanding the Enormous Diversity of Bacteriophages: The Tailed Phages That Infect the Bacterial Family Enterobacteriaceae. Virology 2014, 468–470, 421–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, E.S.; Kutter, E.; Mosig, G.; Arisaka, F.; Kunisawa, T.; Rüger, W. Bacteriophage T4 Genome. Microbiol. Mol. Biol. Rev. 2003, 67, 86–156. [Google Scholar] [CrossRef] [Green Version]
- Trojet, S.N.; Caumont-Sarcos, A.; Perrody, E.; Comeau, A.M.; Krisch, H.M. The gp38 Adhesins of the T4 Superfamily: A Complex Modular Determinant of the Phage’s Host Specificity. Genome Biol. Evol. 2011, 3, 674–686. [Google Scholar] [CrossRef] [Green Version]
- Petrov, V.M.; Ratnayaka, S.; Nolan, J.M.; Miller, E.S.; Karam, J.D. Genomes of the T4-Related Bacteriophages as Windows on Microbial Genome Evolution. Virol. J. 2010, 7, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Roer, L.; Johannesen, T.B.; Hansen, F.; Stegger, M.; Tchesnokova, V.; Sokurenko, E.; Garibay, N.; Allesøe, R.; Thomsen, M.C.F.; Lund, O.; et al. CHTyper, a Web Tool for Subtyping of Extraintestinal Pathogenic Escherichia Coli Based on the and Alleles. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [Green Version]
- Pham-Khanh, N.H.; Sunahara, H.; Yamadeya, H.; Sakai, M.; Nakayama, T.; Yamamoto, H.; Truong Thi Bich, V.; Miyanaga, K.; Kamei, K. Isolation, Characterisation and Complete Genome Sequence of a Tequatrovirus Phage, Escherichia Phage KIT03, Which Simultaneously Infects Escherichia Coli O157:H7 and Salmonella Enterica. Curr. Microbiol. 2019, 76, 1130–1137. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, Y.; Liu, Y.; Pei, J.; Yao, S.; Cheng, C. Bacteriophage Therapy against Enterobacteriaceae. Virol. Sin. 2015, 30, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Brüssow, H. Phage Therapy: The Escherichia coli Experience. Microbiology 2005, 151, 2133–2140. [Google Scholar] [CrossRef] [Green Version]
- Hertz, F.B.; Schønning, K.; Rasmussen, S.C.; Littauer, P.; Knudsen, J.D.; Løbner-Olesen, A.; Frimodt-Møller, N. Epidemiological Factors Associated with ESBL- and Non ESBL-Producing E. coli Causing Urinary Tract Infection in General Practice. Infect. Dis. 2016, 48, 241–245. [Google Scholar] [CrossRef]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The Population Genetics of Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Finn, T.J. The Evolutionary Puzzle of Escherichia coli ST131. Infect. Genet. Evol. 2020, 81, 104265. [Google Scholar] [CrossRef]
- Mirzaei, M.K.; Nilsson, A.S. Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar] [CrossRef] [Green Version]
- Villarroel, J.; Larsen, M.V.; Kilstrup, M.; Nielsen, M. Metagenomic Analysis of Therapeutic PYO Phage Cocktails from 1997 to 2014. Viruses 2017, 9, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitner, L.; Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Chkonia, I.; Rigvava, S.; Chkhotua, A.; Changashvili, G.; McCallin, S.; Schneider, M.P.; et al. Intravesical bacteriophages for treating urinary tract infections in patients undergoing transurethral resection of the prostate: A randomised, placebo-controlled, double-blind clinical trial. Lancet Infect. Dis. 2021, 21, 427–436. [Google Scholar] [CrossRef]
- Alteri, C.J.; Smith, S.N.; Mobley, H.L.T. Fitness of Escherichia coli during Urinary Tract Infection Requires Gluconeogenesis and the TCA Cycle. PLoS Pathog. 2009, 5, e1000448. [Google Scholar] [CrossRef] [Green Version]
- Lacerda Mariano, L.; Ingersoll, M.A. The immune response to infection in the bladder. Nat. Rev. Urol. 2020, 17, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Grygorcewicz, B.; Wojciuk, B.; Roszak, M.; Łubowska, N.; Błażejczak, P.; Jursa-Kulesza, J.; Rakoczy, R.; Masiuk, H.; Dołęgowska, B. Environmental Phage-Based Cocktail and Antibiotic Combination Effects on Acinetobacter baumannii Biofilm in a Human Urine Model. Microb. Drug Resist. 2021, 27, 25–35. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| No. of Sensitive Strains 2 | ||||||
|---|---|---|---|---|---|---|
| Genomic Group | CH Type 1 | No. of Strains | No. of AMR Strains (ESBL) | vKMB22 | vKMB26 | Cocktail |
| A | 11–54 | 5 | 5 (1) | 0/0 | 0/0 | 0/0 |
| 11–23 | 1 | 0 (0) | 0/0 | 0/0 | 0/0 | |
| B1 | 6–31 | 2 | 1 (0) | 0/0 | 1/2 | 1/2 |
| 4–54 | 1 | 0 (0) | 0/0 | 0/1 | 0/1 | |
| B2 | 13–5 | 1 | 0 (0) | 0/0 | 1/1 | 1/1 |
| 13–233 | 1 | 1 (0) | 0/0 | 1/1 | 1/1 | |
| 14–27 | 1 | 0 (0) | 0/0 | 1/1 | 1/1 | |
| 24–10 | 3 | 2 (2) | 1/1 | 1/3 | 3/3 | |
| 24–103 | 1 | 0 (0) | 0/1 | 1/1 | 1/1 | |
| 24–30 | 4 | 0 (0) | 0/0 | 4/4 | 4/4 | |
| 38–30 | 1 | 1 (1) | 0/1 | 1/1 | 1/1 | |
| 40–30 | 5 | 5 (4) | 2/5 | 1/5 | 4/5 | |
| 40–41 | 6 | 5 (5) | 3/6 | 5/6 | 4/6 | |
| 52–5 | 1 | 0 (0) | 0/0 | 0/1 | 0/1 | |
| D | 26–0 | 2 | 2 (0) | 2/2 | 2/2 | 2/2 |
| 35–27 | 1 | 1 (0) | 0/0 | 0/1 | 0/1 | |
| 35–47 | 1 | 0 (0) | 0/1 | 1/1 | 0/1 | |
| 35–483 | 1 | 0 (0) | 0/0 | 0/0 | 0/0 | |
| 37–56 | 2 | 2 (1) | 1/1 | 2/2 | 0/2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slobodníková, L.; Markusková, B.; Kajsík, M.; Andrezál, M.; Straka, M.; Liptáková, A.; Drahovská, H. Characterization of Anti-Bacterial Effect of the Two New Phages against Uropathogenic Escherichia coli. Viruses 2021, 13, 1348. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071348
Slobodníková L, Markusková B, Kajsík M, Andrezál M, Straka M, Liptáková A, Drahovská H. Characterization of Anti-Bacterial Effect of the Two New Phages against Uropathogenic Escherichia coli. Viruses. 2021; 13(7):1348. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071348
Chicago/Turabian StyleSlobodníková, Lívia, Barbora Markusková, Michal Kajsík, Michal Andrezál, Marek Straka, Adriána Liptáková, and Hana Drahovská. 2021. "Characterization of Anti-Bacterial Effect of the Two New Phages against Uropathogenic Escherichia coli" Viruses 13, no. 7: 1348. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071348