
A Mortality-Based Description of EHDV and BTV Prevalence in Farmed White-Tailed Deer (Odocoileus virginianus) in Florida, USA


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Sample Collection
2.3. Laboratory Analysis
2.4. Statistical Analysis
3. Results
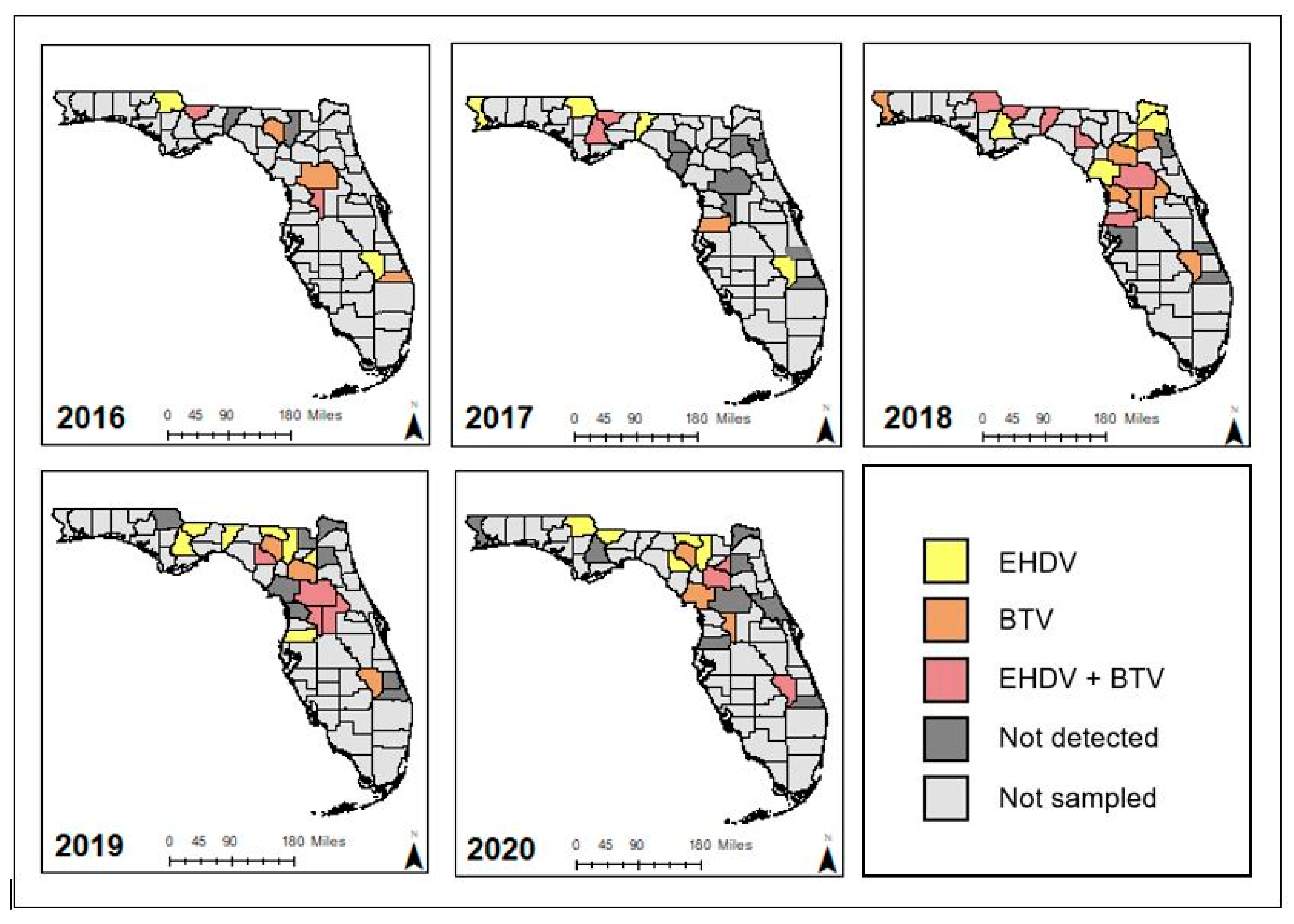
3.1. Descriptive Sampling Summary
3.2. Prevalence of EHDV and BTV in Dead Farmed Deer
3.3. Predominant Circulating EHDV Serotypes
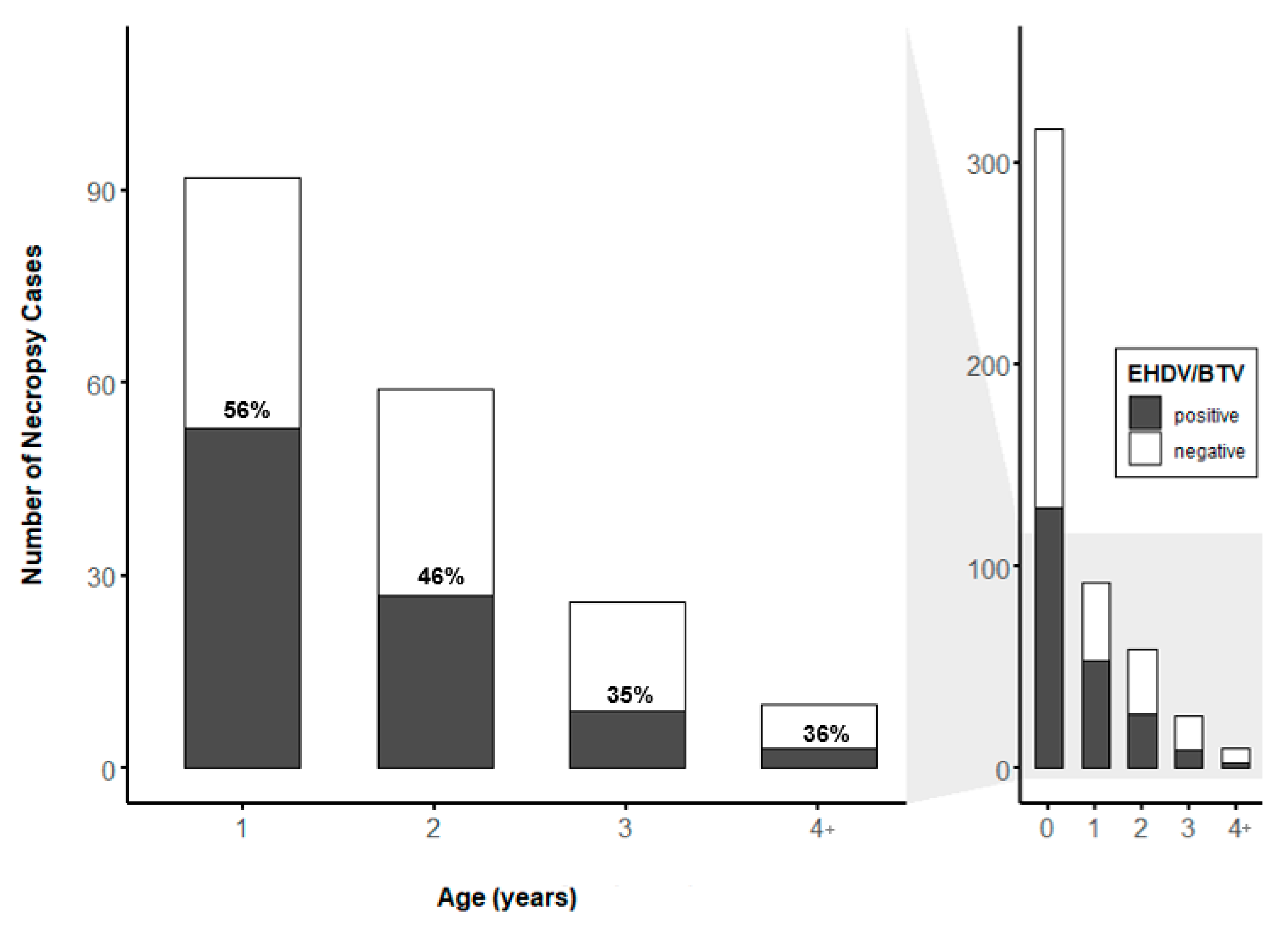
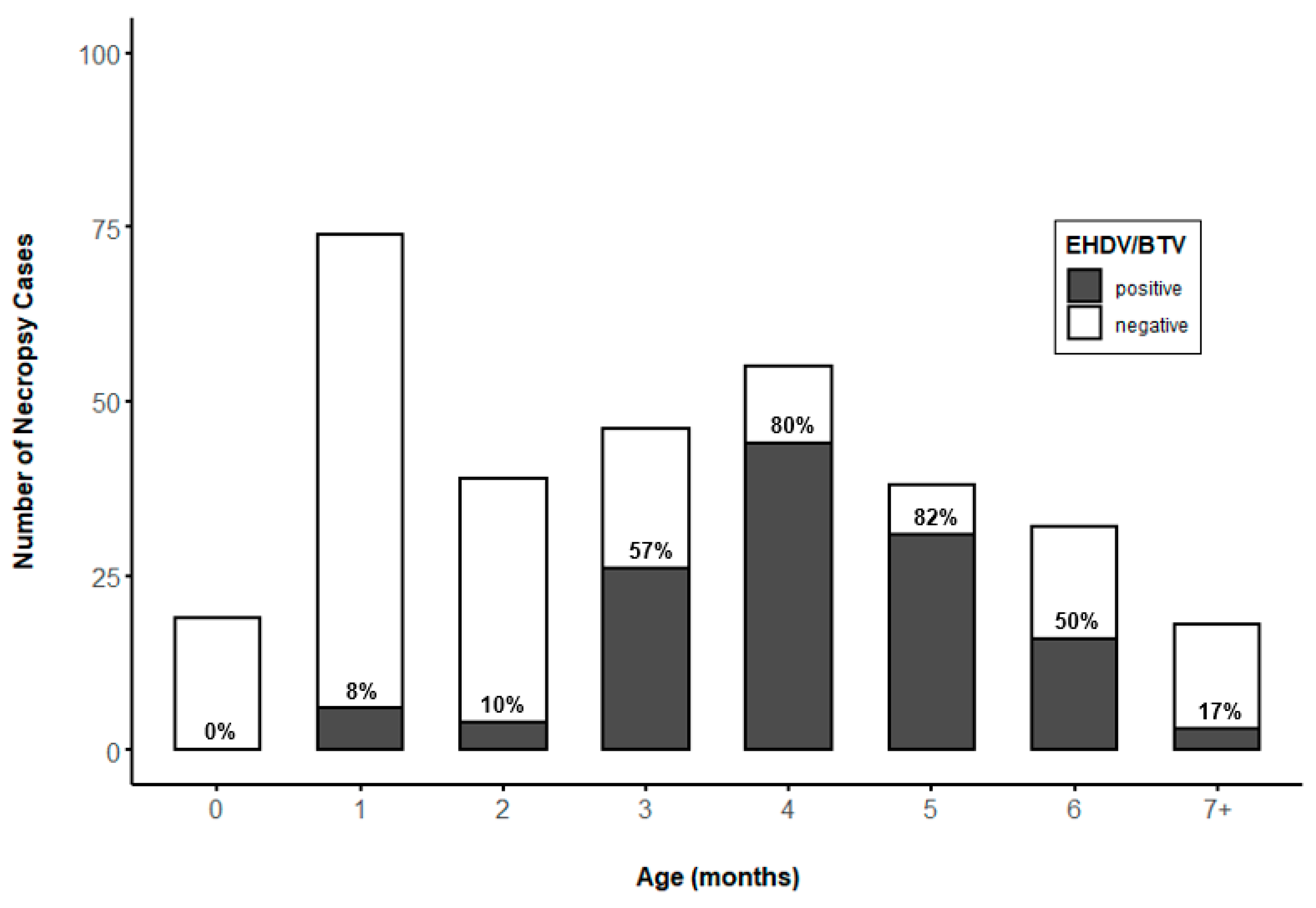
3.4. Prevalence of EHDV and BTV by WTD Age
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, F.C. Hemorrhagic Disease. In Diseases and Parasites of White-Tailed Deer; Davidson, W.A., Hayes, F.A., Nettles, V.F., Kellogg, F.E., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 1981; ISSN 0496-764X. [Google Scholar]
- Gibbs, E.P.; Greiner, E.C. Bluetongue and Epizootic Hemorrhagic Disease. In The Arboviruses: Epidemiology and Ecology; CRC Press: Boca Raton, FL, USA, 1988; Volume 2, ISBN 0-8493-4386-0. [Google Scholar]
- Gibbs, E.P.J.; Greiner, E.C. The Epidemiology of Bluetongue. Comp. Immunol. Microbiol. Infect. Dis. 1994, 17, 207–220. [Google Scholar] [CrossRef]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P.J. Transmission and Epidemiology of Bluetongue and Epizootic Hemorrhagic Disease in North America: Current Perspectives, Research Gaps, and Future Directions. Vector Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Prestwood, A.K.; Kistner, T.P.; Kellogg, F.E.; Hayes, F.A. The 1971 Outbreak of hemorrhagic disease among white-tailed deer of the southeastern United States. J. Wildl. Dis. 1974, 10, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Stallknecht, D.E.; Nettles, V.F.; Rollor, E.A., III; Howerth, E.W. Epizootic hemorrhagic disease virus and bluetongue virus serotype distribution in white-tailed deer in Georgia. J. Wildl. Dis. 1995, 31, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruder, M.G.; Johnson, D.; Ostlund, E.; Allison, A.B.; Kienzle, C.; Phillips, J.E.; Poulson, R.L.; Stallknecht, D.E. The First 10 Years (2006–15) of Epizootic Hemorrhagic Disease Virus Serotype 6 in the USA. J. Wildl. Diss. 2017, 53, 901–905. [Google Scholar] [CrossRef]
- Nettles, V.F.; Davidson, W.R.; Stallknecht, D.E. Surveillance for Hemorrhagic Disease in White-Tailed Deer and Other Wild Ruminants, 1980–1989. In Proceedings of the Annual Conference of the Southeastern Association of Fish and Wildlife Agencies, Corpus Christie, TX, USA, 25–28 October 1992; Volume 46, pp. 138–146. [Google Scholar]
- Stallknecht, D.E.; Blue, J.L.; Rollor, E.A., III; Nettles, V.F.; Davidson, W.R.; Pearson, J.E. Precipitating antibodies to epizootic hemorrhagic disease and bluetongue viruses in white-tailed deer in the southeastern United States. J. Wildl. Dis. 1991, 27, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Stallknecht, D.E.; Luttrell, M.P.; Smith, K.E.; Nettles, V.F. Hemorrhagic Disease in White-Tailed Deer in Texas: A Case for Enzootic Stability. J. Wildl. Dis. 1996, 32, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Stilwell, N.K.; Clarke, L.L.; Howerth, E.W.; Kienzle-Dean, C.; Fojtik, A.; Hollander, L.P.; Carter, D.; Osborn, D.A.; D’Angelo, G.J.; Miller, K.V.; et al. The Effect of Maternal Antibodies on Clinical Response to Infection with Epizootic Hemorrhagic Disease Virus in White-Tailed Deer (Odocoileus Virginianus) Fawns. J. Wildl. Dis. 2020, 57, 189–193. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Howerth, E.W. Epidemiology of Bluetongue and Epizootic Haemorrhagic Disease in Wildlife: Surveillance Methods. Vet. Ital. 2004, 40, 203–207. [Google Scholar]
- Florida Fish and Wildlife Conservation Commission. Rule 68A-4.0051. Importation of Deer, Elk and Other Wildlife Species in the Family of Cervidae; Rulemaking Authority Art. IV, Sec. 9, Fla. Const. Law Implemented Art. IV, Sec. 9, Fla. Const. History-New 10-16-02, Amended 2-27-03, 11-12-13; Florida Department of State: Tallahassee, FL, USA, 2013. [Google Scholar]
- Couvillion, C.E.; Nettles, V.F.; Davidson, W.R.; Pearson, J.E.; Gustafson, G.A. Hemorrhagic Disease among White-Tailed Deer in the Southeast from 1971 through 1980. In Proceedings of the US Animal Health Association, St. Louis, MO, USA, 18–20 October 1981; Volume 85, pp. 522–537. [Google Scholar]
- Stallknecht, D.E.; Howerth, E.W.; Gaydos, J.K. Hemorrhagic Disease in White-Tailed Deer: Our Current Understanding of Risk. In Proceedings of the Transactions of the North American Wildlife and Natural Resources Conference, Dallas, TX, USA, 3–7 April 2002; Volume 67, pp. 75–86. [Google Scholar]
- Nettles, V.F.; Stallknecht, D.E. History and Progress in the Study of Hemorrhagic Disease of Deer. In Proceedings of the Transactions of the North American Wildlife and Natural Resources Conference, Corpus Christie, TX, USA, 25–28 October 1992; Volume 57, pp. 499–516. [Google Scholar]
- Jones, R.H.; Roughton, R.D.; Foster, N.M.; Bando, B.M. Culicoides, the Vector of Epizootic Hemorrhagic Disease in White-Tailed Deer in Kentucky in 1971. J. Wildl. Dis. 1977, 13, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, N.M.; Breckon, R.D.; Luekde, A.J.; Jones, R.H.; Metcalf, H.E. T Transmission of two strains of epizootic hemorrhagic disease virus in deer by Culicoides variipennis. J. Wildl. Dis. 1977, 13, 9–16. [Google Scholar] [CrossRef] [PubMed]
- McGregor, B.L.; Sloyer, K.E.; Sayler, K.A.; Goodfriend, O.; Krauer, J.M.C.; Acevedo, C.; Zhang, X.; Mathias, D.; Wisely, S.M.; Burkett-Cadena, N.D. Field Data Implicating Culicoides stellifer and Culicoides venustus (Diptera: Ceratopogonidae) as Vectors of Epizootic Hemorrhagic Disease Virus. Parasit. Vectors 2019, 12, 258. [Google Scholar] [CrossRef] [PubMed]
- Cauvin, A.; Dinh, E.T.N.; Orange, J.P.; Shuman, R.M.; Blackburn, J.K.; Wisely, S.M. Antibodies to Epizootic Hemorrhagic Disease Virus (EHDV) in Farmed and Wild Florida White-Tailed Deer (Odocoileus Virginianus). J. Wildl. Dis. 2020, 56, 208. [Google Scholar] [CrossRef]
- Anderson, D.P.; Frosch, B.J.; Outlaw, J.L. Economic Impact of the Texas Deer Breeding Industry; APFC Research Report; Agricultural & Food Policy Center: College Station, TX, USA, 2007. [Google Scholar]
- McGregor, B.L.; Stenn, T.; Sayler, K.A.; Blosser, E.M.; Blackburn, J.K.; Wisely, S.M.; Burkett-Cadena, N.D. Host Use Patterns of Culicoides Spp. Biting Midges at a Big Game Preserve in Florida, U.S.A., and Implications for the Transmission of Orbiviruses. Med. Vet. Entomol. 2019, 33, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisely, S.M.; Sayler, K. Autogenous Vaccine Field Trial for Epizootic Hemorrhagic Disease Virus and Bluetongue Virus Does Not Result in High Titer to Homologous Virus Serotypes. Int. J. Infect. Dis. 2016, 53, 149–150. [Google Scholar] [CrossRef]
- Wernike, K.; Hoffmann, B.; Beer, M. Simultaneous Detection of Five Notifiable Viral Diseases of Cattle by Single-Tube Multiplex Real-Time RT-PCR. J. Virol. Methods 2015, 217, 28–35. [Google Scholar] [CrossRef]
- Sun, F.; Cochran, M.; Beckham, T.; Clavijo, A. Molecular Typing of Epizootic Hemorrhagic Disease Virus Serotypes by One-Step Multiplex RT-PCR. J. Wildl. Dis. 2014, 50, 639–644. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Fisher, R.A. On the Interpretation of Χ2 from Contingency Tables, and the Calculation of P. J. R. Stat. Soc. 1922, 85, 87–94. [Google Scholar] [CrossRef]
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation, 2021.
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Environmental Systems Research Institute (ESRI). ArcGIS Desktop Geostatistical Analyst; Environmental Systems Research Institute (ESRI): Redlands, CA, USA, 2019. [Google Scholar]
- Gaydos, J.K.; Davidson, W.R.; Elvinger, F.; Howerth, E.W.; Murphy, M.; Stallknecht, D.E. Cross-protection between epizootic hemorrhagic disease virus serotypes 1 and 2 in white-tailed deer. J. Wildl. Dis. 2002, 38, 720–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, A.B.; Goekjian, V.H.; Potgieter, A.C.; Wilson, W.C.; Johnson, D.J.; Mertens, P.P.C.; Stallknecht, D.E. Detection of a Novel Reassortant Epizootic Hemorrhagic Disease Virus (EHDV) in the USA Containing RNA Segments Derived from Both Exotic (EHDV-6) and Endemic (EHDV-2) Serotypes. J. Gen. Virol. 2010, 91, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Wobeser, G.A. How Disease is Detected, Described, and Measured. In Essentials of Disease in Wild Animals; Blackwell Publishing Professional: Ames, IA, USA, 2006; pp. 45–59. ISBN 0-8138-0589-9. [Google Scholar]
- Quist, C.F.; Howerth, E.W.; Stallknecht, D.E.; Brown, J.; Pisell, T.; Nettles, V.F. Host defense responses associated with experimental hemorrhagic disease in white-tailed deer. J. Wildl. Dis. 1997, 33, 584–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sloyer, K.E.; Burkett-Cadena, N.D.; Yang, A.; Corn, J.L.; Vigil, S.L.; McGregor, B.L.; Wisely, S.M.; Blackburn, J.K. Ecological Niche Modeling the Potential Geographic Distribution of Four Culicoides Species of Veterinary Significance in Florida, USA. PLoS ONE 2019, 14, e0206648. [Google Scholar] [CrossRef] [Green Version]
- Elbers, A.R.W.; Koenraadt, C.; Meiswinkel, R. Mosquitoes and Culicoides Biting Midges: Vector Range and the Influence of Climate Change. Rev. Sci. Tech. 2015, 34, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, J.C.; Stevens, D.R. Some Production Outcomes When Management Practices and Deer Behavior Interact. NZGA Res. Pract. Ser. 2002, 9, 73–77. [Google Scholar] [CrossRef]
- Grimstad, P.R.; Williams, D.G.; Schmitt, S.M. Infection of white-tailed deer (Odocoileus virginianus) in Michigan with Jamestown Canyon Virus (California serogroup) and the importance of maternal antibody in viral maintenance. J. Wildl. Dis. 1987, 23, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Brenton-Rule, T. Investigations of Apparent Vaccine Breakdown. Deer Branch Course 2001, 18, 19–21. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Name | Sequence 5′-3′ | Genomic Target | Concentration (pmol/reaction) | Reference |
|---|---|---|---|---|
| BTV-NS3-183F | AAA TMT TGG AYA AAG CRA TGT CAA A | NS3 | 10 | Wernicke et al. [24] |
| BTV-NS3-288R | CTY ACR TCA TCA CGA AAC GCT | 10 | ||
| BTV-NS3-242FAM | FAM-AAR GCT GCA TTC GCA TCG TAC GC-BHQ1 | 2 | ||
| EHD NS1 5F | AAA AAG TTC YTC GTC GAC TGC | NS1 | 15 | Wernicke et al. [24] |
| EHD NS1 80R | ATT GGC RTA RTA ACT GTT CAT GTT | 15 | ||
| EHD NS1 TAMRA | TAMRA-ATC GAG ATG GAR CGC TTY TTG AGA AAA T-BHQ2 | 2.5 | This study |
| Positive RT-qPCR Test Results to a: | |||||
|---|---|---|---|---|---|
| Sample Period * | n | EHDV Only | BTV Only | EHDV + BTV b | Total Positive |
| 2016 | 54 | 17 (31%) | 12 (22%) | 4 (7%) | 33 (61%) |
| 2017 | 118 | 38 (32%) | 14 (12%) | 1 (0.8%) | 53 (45%) |
| 2018 | 152 | 27 (18%) | 42 (27%) | 3 (2%) | 72 (47%) |
| 2019 | 126 | 45 (36%) | 9 (7%) | 2 (2%) | 56 (44%) |
| 2020 | 89 | 13 (15%) | 10 (11%) | 0 (0%) | 23 (26%) |
| Total | 539 | 140 (26%) | 87 (16%) | 10 (2%) | 237 (44%) |
| Serotype | |||||
|---|---|---|---|---|---|
| Sample Period * | n | EHDV-1 | EHDV-2 | EHDV-6 | EHDV-2 + -6 |
| 2016 | 18 | 0 (0%) | 17 (94%) | 1 (6%) | 0 (0%) |
| 2017 | 31 | 0 (0%) | 8 (26%) | 22 (71%) | 1 (3%) |
| 2018 | 23 | 0 (0%) | 23 (100%) | 0 (0%) | 0 (0%) |
| 2019 | 41 | 19 (46%) | 11 (27%) | 10 (24%) | 1 (2%) |
| 2020 | 8 | 0 (0%) | 8 (100%) | 0 (0%) | 0 (0%) |
| Total | 121 | 19 (16%) | 67 (55%) | 33 (27%) | 2 (2%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cottingham, S.L.; White, Z.S.; Wisely, S.M.; Campos-Krauer, J.M. A Mortality-Based Description of EHDV and BTV Prevalence in Farmed White-Tailed Deer (Odocoileus virginianus) in Florida, USA. Viruses 2021, 13, 1443. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081443
Cottingham SL, White ZS, Wisely SM, Campos-Krauer JM. A Mortality-Based Description of EHDV and BTV Prevalence in Farmed White-Tailed Deer (Odocoileus virginianus) in Florida, USA. Viruses. 2021; 13(8):1443. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081443
Chicago/Turabian StyleCottingham, Sydney L., Zoe S. White, Samantha M. Wisely, and Juan M. Campos-Krauer. 2021. "A Mortality-Based Description of EHDV and BTV Prevalence in Farmed White-Tailed Deer (Odocoileus virginianus) in Florida, USA" Viruses 13, no. 8: 1443. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081443