
Intravenous, Intratracheal, and Intranasal Inoculation of Swine with SARS-CoV-2

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Animal Experiment
2.3. Nucleic Acid Extraction and Real-Time Reverse Transcriptase PCR (rRT-PCR)
2.4. Virus Isolation
2.5. Serological Analysis
2.6. In Situ Hybridization (ISH)
3. Results
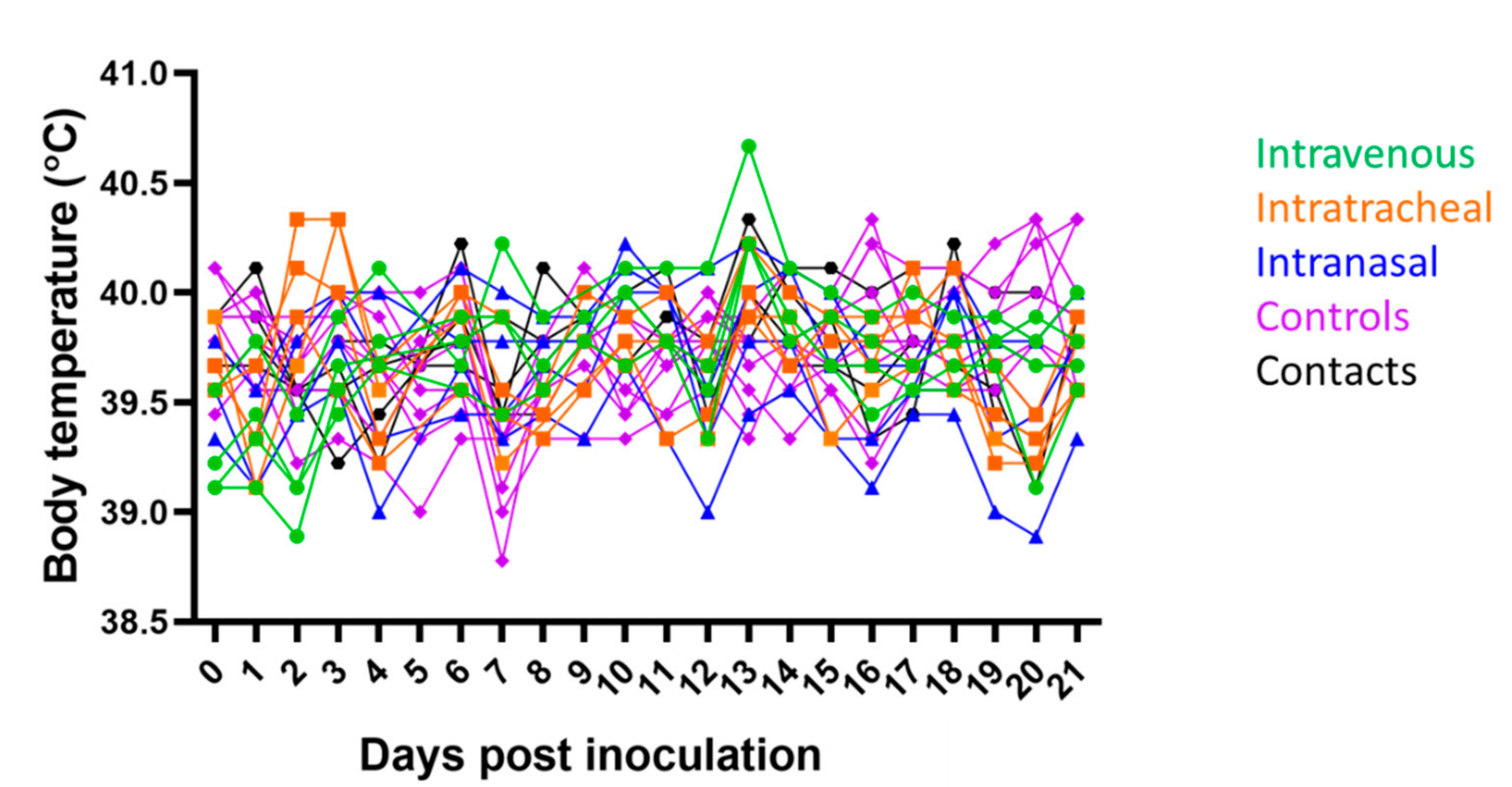
3.1. Clinical Signs
3.2. Nucleic Acid Detection of SARS-CoV-2 and Neutralizing Antibody Response
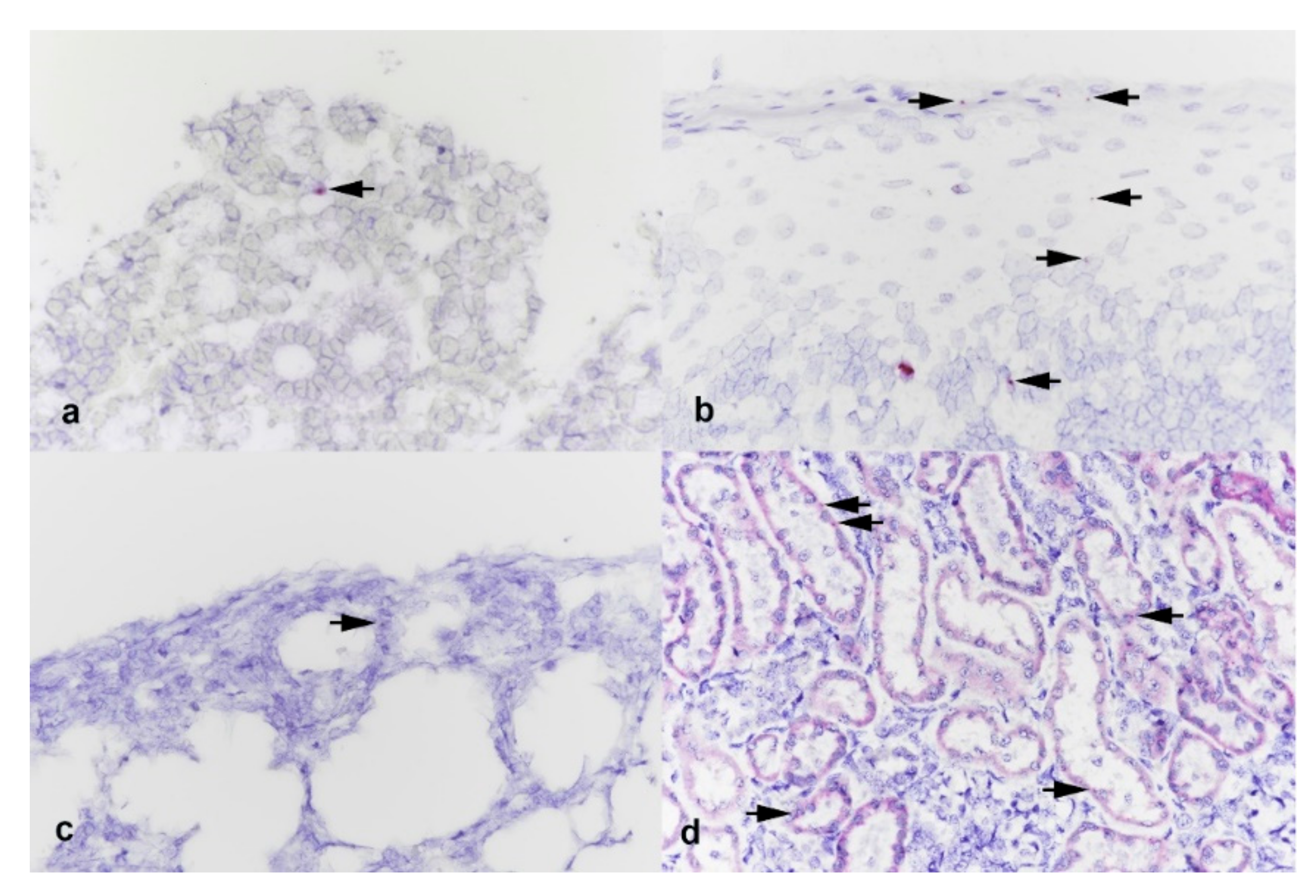
3.3. Histology and In Situ Hybridization
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Wu, T.; Liu, Q.; Yang, Z. The SARS-CoV-2 outbreak: What we know. Int. J. Infect. Dis. 2020, 94, 44–48. [Google Scholar] [CrossRef]
- Zhao, J.; Cui, W.; Tian, B.P. The potential intermediate hosts for SARS-CoV-2. Front. Microbiol. 2020, 11, 580137. [Google Scholar] [CrossRef]
- Guan, Y.J.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Butt, K.M.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Dudas, G.; Carvalho, L.M.; Rambaut, A.; Bedford, T. MERS-CoV spillover at the camel-human interface. eLife 2018, 7, e31257. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.L.; Anderson, T.; Lager, K.M. A brief introduction to influenza a virus in swine. Methods Mol. Biol. 2020, 2123, 249–271. [Google Scholar]
- Chua, K.B. Nipah virus outbreak in Malaysia. J. Clin. Virol. 2003, 26, 265–275. [Google Scholar] [CrossRef]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.-L.; Shi, W.-F.; Zhang, W.; Zhu, Y.; Zhang, Y.-W.; Xie, Q.-M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 2018, 556, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Vlasova, A.N.; Kenney, S.P.; Saif, L.J. Emerging and re-emerging coronaviruses in pigs. Curr. Opin. Virol. 2019, 34, 39–49. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Holmes, E.C. A genomic perspective on the origin and emergence of SARS-CoV-2. Cell 2020, 181, 223–227. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Mora-Díaz, J.C.; Piñeyro, P.E.; Houston, E.; Zimmerman, J.; Giménez-Lirola, L.G. Porcine hemagglutinating encephalomyelitis virus: A review. Front. Veter. Sci. 2019, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.; Pfenning, A.; Zhao, H.; et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor recognition by the novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS coronavirus. J. Virol. 2020, 94, e00127-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, X.; Sun, J.; Yan, Z.; Zhang, J.; Zhao, J.; Zhao, Z.; Gao, Q.; He, W.-T.; Veit, M.; Su, S. Comparison of severe acute respiratory syndrome coronavirus 2 spike protein binding to ACE2 receptors from human, pets, farm animals, and putative intermediate hosts. J. Virol. 2020, 94, e00831-20. [Google Scholar] [CrossRef]
- Meekins, D.A.; Morozov, I.; Trujillo, J.D.; Gaudreault, N.N.; Bold, D.; Carossino, M.; Artiaga, B.L.; Indran, S.V.; Kwon, T.; Balaraman, V.; et al. Susceptibility of swine cells and domestic pigs to SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 2278–2288. [Google Scholar] [CrossRef] [PubMed]
- Mossel, E.C.; Huang, C.; Narayanan, K.; Makino, S.; Tesh, R.B.; Peters, C.J. Exogenous ACE2 expression allows refractory cell lines to support severe acute respiratory syndrome coronavirus replication. J. Virol. 2005, 79, 3846–3850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, e218–e225. [Google Scholar] [CrossRef]
- Vergara-Alert, J.; Rodon, J.; Carrillo, J.; Te, N.; Izquierdo-Useros, N.; de la Concepción, M.L.R.; Ávila-Nieto, C.; Guallar, V.; Valencia, A.; Cantero, G.; et al. Pigs are not susceptible to SARS-CoV-2 infection but are a model for viral immunogenicity studies. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Pickering, B.S.; Smith, G.; Pinette, M.M.; Embury-Hyatt, C.; Moffat, E.; Marszal, P.; Lewis, C.E. Susceptibility of domestic swine to experimental infection with severe acute respiratory syndrome coronavirus 2. Emerg. Infect. Dis. 2021, 27, 104–112. [Google Scholar] [CrossRef]
- Falkenberg, S.; Buckley, A.; Laverack, M.; Martins, M.; Palmer, M.V.; Lager, K.; Diel, D.G. Experimental inoculation of young calves with SARS-CoV-2. Viruses 2021, 13, 441. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Martins, M.; Falkenberg, S.; Buckley, A.; Caserta, L.C.; Mitchell, P.K.; Cassmann, E.D.; Rollins, A.; Zylich, N.C.; Renshaw, R.W. Susceptibility of white-tailed deer (Odocoileus virginianus) to SARS-CoV-2. J. Virol. 2021, 95, e00083-21. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D.; et al. From people to panthera: Natural SARS-CoV-2 infection in tigers and lions at the Bronx zoo. mBio 2020, 11, e02220-20. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Zhang, A.J.; Yuan, S.; Poon, V.K.-M.; Chan, C.C.-S.; Lee, A.C.-Y.; Chan, W.-M.; Fan, Z.; Tsoi, H.-W.; Wen, L.; et al. Simulation of the clinical and pathological manifestations of coronavirus disease 2019 (COVID-19) in a golden syrian hamster model: Implications for disease pathogenesis and transmissibility. Clin. Infect. Dis. 2020, 71, 2428–2446. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.W.; Fung, K.; Choy, K.T.; Wong, A.Y.; Kaewpreedee, P.; Perera, R.A.M.; Poon, L.; Nicholls, J.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and rapid transmission of SARS-CoV-2 in ferrets. Cell Host. Microbe 2020, 27, 704–709. [Google Scholar] [CrossRef]
- Shan, C.; Yao, Y.F.; Yang, X.L.; Zhou, Y.W.; Gao, G.; Peng, Y.; Yang, L.; Hu, X.; Xiong, J.; Jiang, R.D.; et al. Infection with novel coronavirus (SARS-CoV-2) causes pneumonia in Rhesus macaques. Cell Res. 2020, 30, 670–677. [Google Scholar] [CrossRef]
- Munster, V.J.; Feldmann, F.; Williamson, B.N.; van Doremalen, N.; Pérez-Pérez, L.; Schulz, J.; Meade-White, K.; Okumura, A.; Callison, J.; Brumbaugh, B.; et al. Respiratory disease in rhesus macaques inoculated with SARS-CoV-2. Nature 2020, 585, 268–272. [Google Scholar] [CrossRef]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental infection of cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979. [Google Scholar] [CrossRef]
- Sun, K.; Gu, L.; Ma, L.; Duan, Y. Atlas of ACE2 gene expression reveals novel insights into transmission of SARS-CoV-2. Heliyon 2021, 7, e05850. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, J.; Izquierdo-Useros, N.; Ávila-Nieto, C.; Pradenas, E.; Clotet, B.; Blanco, J. Humoral immune responses and neutralizing antibodies against SARS-CoV-2; implications in pathogenesis and protective immunity. Biochem. Biophys. Res. Commun. 2021, 538, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Legros, V.; Denolly, S.; Vogrig, M.; Boson, B.; Siret, E.; Rigaill, J.; Pillet, S.; Grattard, F.; Gonzalo, S.; Verhoeven, P. A longitudinal study of SARS-CoV-2-infected patients reveals a high correlation between neutralizing antibodies and COVID-19 severity. Cell Mol. Immunol. 2021, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Cervia, C.; Nilsson, J.; Zurbuchen, Y.; Valaperti, A.; Schreiner, J.; Wolfensberger, A.; Raeber, M.E.; Adamo, S.; Weigang, S.; Emmenegger, M.; et al. Systemic and mucosal antibody responses specific to SARS-CoV-2 during mild versus severe COVID-19. J. Allergy Clin. Immunol. 2021, 147, 545–557. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | Intravenous | Intratracheal | Intranasal | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample | N/O | R | BC | OF | N/O | R | BC | OF | N/O | R | BC | OF |
| DPI | ||||||||||||
| 0 | 0/4 | 0/4 | 0/4 | NS | 0/4 | 0/4 | 0/4 | NS | 0/4 | 0/4 | 0/4 | NS |
| 1 | 0/4 | 0/4 | NS | NS | 4/4 | 2/4 | NS | NS | 4/4 | 3/4 | NS | NS |
| 2 | 0/4 | 0/4 | NS | NS | 2/4 | 2/4 | NS | NS | 2/4 | 2/4 | NS | NS |
| 3 | 0/4 | 0/4 | 2/4 | NS | 0/4 | 1/4 | 0/4 | NS | 1/4 | 1/4 | 0/4 | NS |
| 4 | 1/4 | 0/4 | NS | 0/1 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 1/4 | NS | 1/1 |
| 5 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 1/4 | NS | 0/1 |
| 6 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 0/4 | NS | 1/1 | 0/4 | 0/4 | NS | 0/1 |
| 7 | 0/4 | 0/4 | 0/4 | 0/1 | 1/4 | 1/4 | 0/4 | 0/1 | 0/4 | 0/4 | 0/4 | 0/1 |
| 10 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 1/4 | NS | 0/1 |
| 12 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 1/4 | NS | 0/1 |
| 14 | 0/4 | 0/4 | 0/4 | 0/1 | 0/4 | 0/4 | 0/4 | 0/1 | 0/4 | 0/4 | 0/4 | 0/1 |
| 18 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 0/4 | NS | 0/1 | 0/4 | 0/4 | NS | 0/1 |
| 21 | 0/4 | 0/4 | 0/4 | 0/1 | 0/4 | 0/4 | 0/4 | 0/1 | 0/4 | 0/4 | 0/4 | 0/1 |
| Pig # | Group | 3 dpi | 7 dpi | 14 dpi | 21 dpi |
|---|---|---|---|---|---|
| 344 | IV | 8 | 64 | 8 | <8 |
| 345 | IV | <8 | 32 | 32 | 8 |
| 346 | IV | <8 | 64 | 16 | 8 |
| 347 | IV | <8 | 32 | 8 | 8 |
| 348 | IT | <8 | <8 | <8 | <8 |
| 349 | IT | <8 | <8 | <8 | <8 |
| 350 | IT | <8 | <8 | 8 | 32 |
| 351 | IT | <8 | <8 | 16 | 8 |
| 352 | IN | <8 | <8 | 32 | 16 |
| 353 | IN | <8 | <8 | <8 | <8 |
| 354 | IN | <8 | <8 | 8 | <8 |
| 355 | IN | <8 | <8 | <8 | <8 |
| 364 | Contact | <8 | <8 | <8 | <8 |
| 365 | Contact | <8 | <8 | <8 | <8 |
| 366 | Contact | <8 | <8 | <8 | <8 |
| Pig # | Group | Nasal Turbinates | Tonsil | Mandibular LN | Tracheobronchial LN | Lung |
|---|---|---|---|---|---|---|
| 344 | IV | - | 41.62 | 39.94 | 38.25 | - |
| 345 | IV | - | - | - | - | - |
| 346 | IV | - | - | - | - | - |
| 347 | IV | - | - | - | - | - |
| 348 | IT | - | - | 39.98 | - | - |
| 349 | IT | - | - | - | - | - |
| 350 | IT | - | - | - | 34.67 | - |
| 351 | IT | - | - | - | - | - |
| 352 | IN | - | - | - | 39.52 | - |
| 353 | IN | - | - | - | - | - |
| 354 | IN | - | 39.91 | - | - | - |
| 355 | IN | - | - | - | - | - |
| 364 | Contact | - | - | 39.55 | - | - |
| 365 | Contact | - | - | - | - | - |
| 366 | Contact | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buckley, A.; Falkenberg, S.; Martins, M.; Laverack, M.; Palmer, M.V.; Lager, K.; Diel, D.G. Intravenous, Intratracheal, and Intranasal Inoculation of Swine with SARS-CoV-2. Viruses 2021, 13, 1506. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081506
Buckley A, Falkenberg S, Martins M, Laverack M, Palmer MV, Lager K, Diel DG. Intravenous, Intratracheal, and Intranasal Inoculation of Swine with SARS-CoV-2. Viruses. 2021; 13(8):1506. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081506
Chicago/Turabian StyleBuckley, Alexandra, Shollie Falkenberg, Mathias Martins, Melissa Laverack, Mitchell V. Palmer, Kelly Lager, and Diego G. Diel. 2021. "Intravenous, Intratracheal, and Intranasal Inoculation of Swine with SARS-CoV-2" Viruses 13, no. 8: 1506. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081506