
Characterisation and Distribution of Karaka Ōkahu Purepure Virus—A Novel Emaravirus Likely to Be Endemic to New Zealand



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Virus Discovery and Genome Characterisation
2.1.1. Plant Material, Nucleic Acid Isolation, and Sequencing Platform
2.1.2. Genome and Protein Analysis
2.1.3. Phylogenetic Analysis
2.2. Survey for KŌPV
2.2.1. Geospatial Location for Plant Collection
2.2.2. Plant Material and RNA Isolation
2.2.3. Diagnostic Method
3. Results
3.1. Virus Discovery and Genome Analysis
3.2. Phylogenetic Analysis
3.3. Detection and Distribution
4. Discussion
4.1. Genome Analysis
4.2. Phylogeny
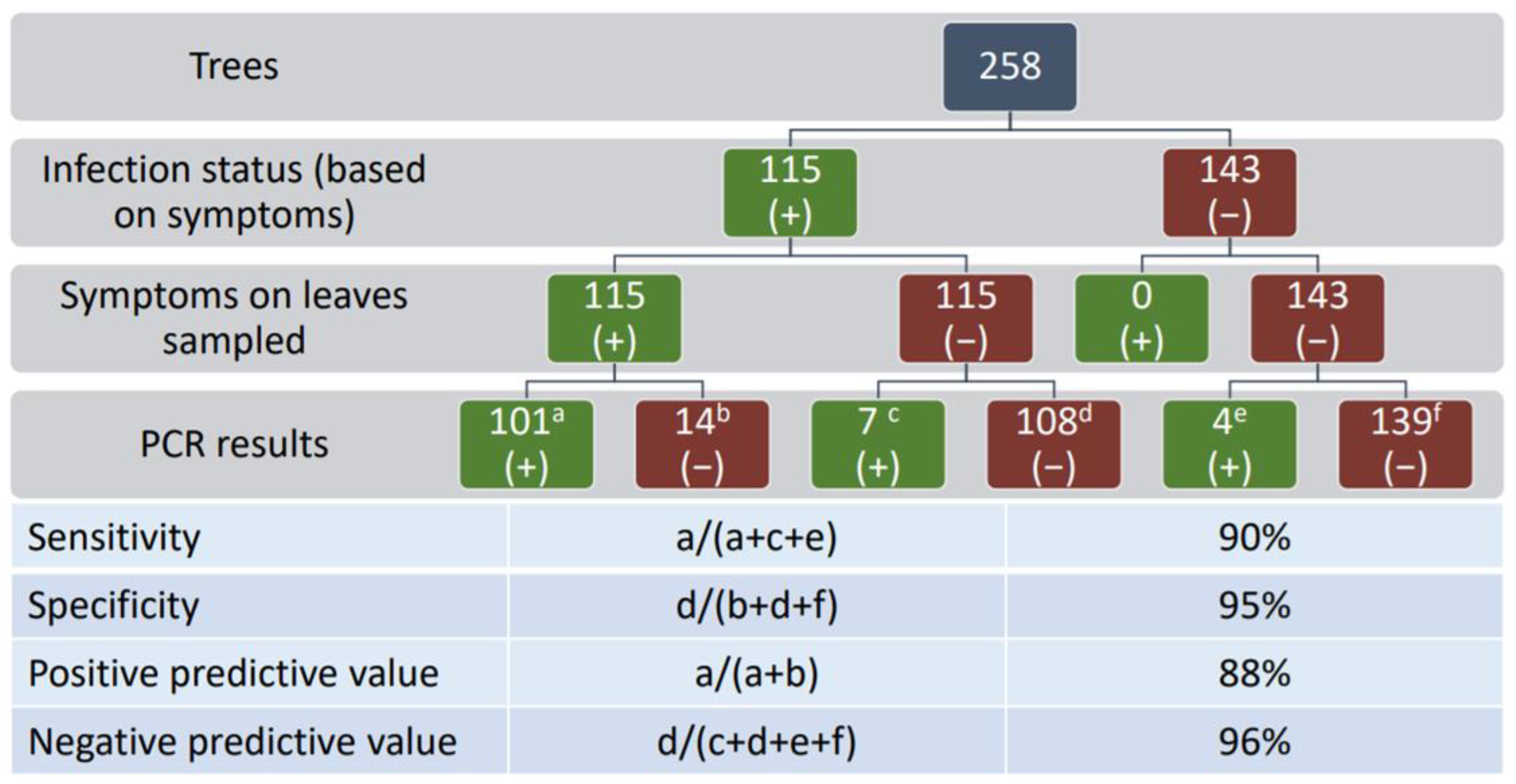
4.3. Diagnostics and Distribution
4.4. Cultural Importance and Potential to be Endemic
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Lange, P.J. Corynocarpus Laevigatus Fact Sheet (Content Continuously Updated). New Zealand Plant Conservation Network. Available online: https://www.nzpcn.org.nz/flora/species/corynocarpus-laevigatus/ (accessed on 25 May 2021).
- Campbell, D.J.; Atkinson, I.A. Depression of tree recruitment by the Pacific rat (Rattus exulans Peale) on New Zealand’s northern offshore islands. Biol. Conserv. 2002, 107, 19–35. [Google Scholar] [CrossRef]
- Costall, J.A.; Carter, R.J.; Shimada, Y.; Anthony, D.; Rapson, G.L. The endemic tree Corynocarpus laevigatus (karaka) as a weedy invader in forest remnants of southern North Island, New Zealand. N. Z. J. Bot. 2006, 44, 5–22. [Google Scholar] [CrossRef]
- Macaskill, J.J.; Manley-Harris, M.; Field, R.J. Quantification of nitropropanoyl glucosides in karaka nuts before and after treatment. Food Chem. 2015, 175, 543–548. [Google Scholar] [CrossRef]
- Sawyer, S.; McFadgen, B.; Hughes, P. Karaka (Corynocarpus laevigatus J.R. et G. Forst.) in Wellington Conservancy (Excluding Chatham Islands); DOC Science Internal Series 101; Department of Conservation: Wellington, New Zealand, 2003; Volume 2, pp. 1–27.
- Van Essen, R.; Rapson, G. Fruit size of karaka (Corynocarpus laevigatus) in relation to potential selection by Maori. N. Z. Bot. Soc. Newsl. 2005, 81, 13–16. [Google Scholar]
- Brown, P.; Newstrom-Lloyd, L.E.; Foster, B.J.; Badger, P.H.; McLean, J.A. Winter 2016 honey bee colony losses in New Zealand. J. Apic. Res. 2018, 57, 278–291. [Google Scholar] [CrossRef] [Green Version]
- Hunt, H.; Cave, N.; Gartrell, B.; Cogger, N.; Petersen, J.; Roe, W. An epidemiological investigation of an idiopathic myopathy in hunting dogs in New Zealand. N. Z. Vet. J. 2018, 66, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Leach, H.; Stowe, C. Oceanic arboriculture at the margins-The case of the Karaka (Corynocarpus laevigatus) in Aotearoa. J. Polyn. Soc. 2005, 114, 7–27. [Google Scholar]
- Ridley, G.S.; Bain, J.; Bulman, L.S.; Dick, M.A.; Kay, M.K. Threats to New Zealand’s indigenous forests from exotic pathogens and pests. Sci. Conserv. 2000, 142, 1–67. [Google Scholar]
- Ashby, J. Infection of karaka (Corynocarpus laevigatus JR & G. Forst.) by cucumber mosaic virus. N. Z. J. Agric. Res. 1977, 20, 533–534. [Google Scholar]
- Veerakone, S.; Tang, J.Z.; Ward, L.I.; Liefting, L.W.; Perez-Egusquiza, Z.; Lebas, B.S.M.; Delmiglio, C.; Fletcher, J.D.; Guy, P.L. A review of the plant virus, viroid, liberibacter and phytoplasma records for New Zealand. Australas. Plant Pathol. 2015, 44, 463–514. [Google Scholar] [CrossRef]
- Xue, X.F.; Zhang, Z.Q. New Zealand Eriophyoidea (Acari: Prostigmata): An update with descriptions of one new genus and six new species. Zootaxa 2008, 1962, 1–32. [Google Scholar] [CrossRef]
- Kormelink, R.; Verchot, J.; Tao, X.; Desbiez, C. The Bunyavirales: The Plant-Infecting Counterparts. Viruses 2021, 13, 842. [Google Scholar] [CrossRef]
- Kulkarni, N.K.; Kumar, P.L.; Muniyappa, V.; Jones, A.T.; Reddy, D.V.R. Transmission of pigeonpea sterility mosaic virus by the eriophyid mite, Aceria cajani Acari, Arthropoda. Plant Dis. 2002, 86, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.L.; Jones, A.T.; Reddy, D.V.R. A novel mite-transmitted virus with a divided RNA genome closely associated with pigeonpea sterility mosaic disease. Phytopathology 2003, 93, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Di Bello, P.L.; Thekke-Veetil, T.; Druciarek, T.; Tzanetakis, I.E. Transmission attributes and resistance to rose rosette virus. Plant Pathol. 2018, 67, 499–504. [Google Scholar] [CrossRef]
- Hassan, M.; Di Bello, P.L.; Keller, K.E.; Martin, R.R.; Sabanadzovic, S.; Tzanetakis, I.E. A new, widespread emaravirus discovered in blackberry. Virus Res. 2017, 235, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, K.; Usugi, T.; Tomitaka, Y.; Shimomoto, Y.; Takeuchi, S.; Kadono, F.; Yanagisawa, H.; Chiaki, Y.; Tsuda, S. Perilla mosaic virus is a highly divergent emaravirus transmitted by Shevtchenkella sp. (acari: Eriophyidae). Phytopathology 2020, 110, 1352–1361. [Google Scholar] [CrossRef]
- Martin, N.A. Karaka gall mite-Aculus corynocarpi. In Interesting Insects and Other Invertebrates; New Zealand Arthropod Factsheet Series 2017, Number 87; Manaaki Whenua-Landcare Research and Plant & Food Research: Lincoln, New Zealand, 2017. [Google Scholar]
- Mielke-Ehret, N.; Mühlbach, H.P. Emaravirus: A novel genus of multipartite, negative strand RNA plant viruses. Viruses 2012, 4, 1515–1536. [Google Scholar] [CrossRef] [Green Version]
- Mielke, N.; Muehlbach, H.-P. A novel, multipartite, negative-strand RNA virus is associated with the ringspot disease of European mountain ash (Sorbus aucuparia L.). J. Gen. Virol. 2007, 88, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, Y.; Wang, G.; Hong, J.; Yang, Z.; Bai, J.; Hong, N. Molecular characteristics of Jujube yellow mottle-associated virus infecting Jujube (Ziziphus jujube Mill.) Grown at Aksu in Xinjiang of China. Viruses 2020, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Peracchio, C.; Forgia, M.; Chiapello, M.; Vallino, M.; Turina, M.; Ciuffo, M. A complex virome including two distinct emaraviruses associated with virus-like symptoms in Camellia japonica. Virus Res. 2020, 286, 197964. [Google Scholar] [CrossRef]
- ICTV. International Committee on Taxonomy Virus (ICTV): 2019 Release. 2019. Retrieved from Washington, DC, USA. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 31 March 2021).
- Blouin, A.G.; Ross, H.A.; Hobson-Peters, J.; O’Brien, C.A.; Warren, B.; MacDiarmid, R. A new virus discovered by immunocapture of double-stranded RNA, a rapid method for virus enrichment in metagenomic studies. Mol. Ecol. Resour. 2016, 16, 1255–1263. [Google Scholar] [CrossRef] [Green Version]
- Podolyan, A.; Blouin, A.G.; Dhami, M.K.; Veerakone, S.; MacDiarmid, R. First report of Ageratum latent virus in Veronica sp. Australas. Plant Dis. Notes 2020, 15, 39. [Google Scholar] [CrossRef]
- White, E.J.; Venter, M.; Hiten, N.F.; Burger, J.T. Modified cetyltrimethylammonium bromide method improves robustness and versatility: The benchmark for plant RNA extraction. Biotech. J. 2008, 3, 1424–1428. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Jung, E.; Brunak, S. Prediction of N-glycosylation sites in human proteins. NetNGlyc 1.0. 2004. Available online: http://www.cbs.dtu.dk/services/NetNGlyc/ (accessed on 25 May 2021).
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K. TMbase-A database of membrane spanning proteins segments. Biol. Chem. Hoppe-Seyler 1993, 374, 166. [Google Scholar]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef]
- Lupas, A.; Van Dyke, M.; Stock, J. Predicting coiled coils from protein sequences. Science 1991, 252, 1162–1164. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 3, 1547–1549. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- QGIS. QGIS Geographic Information System. 2019. Available online: http://qgis.org (accessed on 25 May 2021).
- Chooi, K.M.; Cohen, D.; Pearson, M.N. Generic and sequence-variant specific molecular assays for the detection of the highly variable Grapevine leafroll-associated virus 3. J. Virol. Methods 2013, 189, 20–29. [Google Scholar] [CrossRef]
- Elbeaino, T.; Whitfield, A.; Sharma, M.; Digiaro, M. Emaravirus-specific degenerate PCR primers allowed the identification of partial RNA-dependent RNA polymerase sequences of Maize red stripe virus and Pigeonpea sterility mosaic virus. J. Virol. Methods 2013, 188, 37–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Bello, P.L.; Laney, A.G.; Druciarek, T.; Ho, T.; Gergerich, R.C.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. A novel emaravirus is associated with redbud yellow ringspot disease. Virus Res. 2016, 222, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbeaino, T.; Digiaro, M.; Martelli, G.P. Complete nucleotide sequence of four RNA segments of fig mosaic virus. Arch. Virol. 2009, 154, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Navarro, B.; Wang, G.; Wang, Y.; Yang, Z.; Xu, W.; Zhu, C.; Wang, L.; Serio, F.D.; Hong, N. Actinidia chlorotic ringspot-associated virus: A novel emaravirus infecting kiwifruit plants. Mol. Plant Pathol. 2017, 18, 569–581. [Google Scholar] [CrossRef] [PubMed]
- McGavin, W.J.; Mitchell, C.; Cock, P.J.; Wright, K.M.; MacFarlane, S.A. Raspberry leaf blotch virus, a putative new member of the genus Emaravirus, encodes a novel genomic RNA. J. Gen. Virol. 2012, 93, 430–437. [Google Scholar] [CrossRef]
- Tatineni, S.; McMechan, A.J.; Wosula, E.N.; Wegulo, S.N.; Graybosch, R.A.; French, R.; Hein, G.L. An eriophyid mite-transmitted plant virus contains eight genomic RNA segments with unusual heterogeneity in the nucleocapsid protein. J. Virol. 2014, 88, 11834–11845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Zhang, S.; Han, T.; Fu, J.; Di Serio, F.; Cao, M. Identification and characterization of a novel emaravirus associated with jujube (Ziziphus jujuba Mill.) yellow mottle disease. Front. Microbiol. 2019, 10, 1417. [Google Scholar] [CrossRef] [Green Version]
- Laney, A.G.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. A discovery 70 years in the making: Characterization of the Rose rosette virus. J. Gen. Virol. 2011, 92, 1727–1732. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Yanagisawa, H.; Chiaki, Y.; Yamasaki, J.; Horikawa, H.; Tsunekawa, K.; Morita, Y.; Kadono, F. Complete nucleotide sequence of chrysanthemum mosaic-associated virus, a novel emaravirus infecting chrysanthemum. Arch. Virol. 2021, 166, 1241–1245. [Google Scholar] [CrossRef]
- Rehanek, M.; von Bargen, S.; Bandte, M.; Karlin, D.G.; Büttner, C. A novel emaravirus comprising five RNA segments is associated with ringspot disease in oak. Arch. Virol. 2021, 166, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Rumbou, A.; Candresse, T.; von Bargen, S.; Büttner, C. Next-generation sequencing reveals a novel emaravirus in diseased maple trees from a German urban forest. Front. Microbiol. 2021, 11, 621179. [Google Scholar] [CrossRef]
- Wang, Y.; Zhai, L.; Wen, S.; Yang, Z.; Wang, G.; Hong, N. Molecular characterization of a novel emaravirus infecting Actinidia spp. in China. Virus Res. 2020, 275, 197736. [Google Scholar] [CrossRef]
- von Bargen, S.; Dieckmann, H.L.; Candresse, T.; Mühlbach, H.P.; Roßbach, J.; Büttner, C. Determination of the complete genome sequence of European mountain ash ringspot-associated emaravirus from Sorbus intermedia reveals two additional genome segments. Arch. Virol. 2019, 164, 1937–1941. [Google Scholar] [CrossRef]
- Di Bello, P.L.; Ho, T.; Tzanetakis, I.E. The evolution of emaraviruses is becoming more complex: Seven segments identified in the causal agent of Rose rosette disease. Virus Res. 2015, 210, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Digiaro, M.; Mielke-Ehret, N.; Mühlbach, H.-P.; Martelli, G.P. ICTV virus taxonomy profile: Fimoviridae. J. Gen. Virol. 2018, 99, 1478–1479. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Hein, G.L.; Graybosch, R.A.; Tatineni, S. Octapartite negative-sense RNA genome of High Plains wheat mosaic virus encodes two suppressors of RNA silencing. Virology 2018, 518, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Hein, G.L.; Tatineni, S. P7 and P8 proteins of High Plains wheat mosaic virus, a negative-strand RNA virus, employ distinct mechanisms of RNA silencing suppression. Virology 2019, 535, 20–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormelink, R.; Garcia, M.L.; Goodin, M.; Sasaya, T.; Haenni, A.L. Negative-strand RNA viruses: The plant-infecting counterparts. Virus Res. 2011, 162, 184–202. [Google Scholar] [CrossRef]
- Liu, H.; Wang, G.; Yang, Z.; Wang, Y.; Zhang, Z.; Li, L.; Waqas, M.; Hong, N.; Liu, H.; Wang, G.; et al. Identification and characterization of a pear chlorotic leaf spot-associated virus, a novel Emaravirus associated with a severe disease of pear trees in China. Plant Dis. 2020, 104, 2786–2798. [Google Scholar] [CrossRef]
- Atherton, R.; Lockhart, P.; McLenachan, P.; de Lange, P.; Wagstaff, S.; Shepherd, L.A. A molecular investigation into the origin and relationships of karaka/kōpi (Corynocarpus laevigatus) in New Zealand. J. R. Soc. N. Z. 2015, 45, 212–220. [Google Scholar] [CrossRef]
- Kubota, K.; Chiaki, Y.; Yanagisawa, H.; Takeyama, S.; Suzuki, R.; Kohyama, M.; Horikawa, T.; Toda, S.; Kadono, F. First report of pear chlorotic leaf spot-associated virus on Japanese and European pears in Japan and its detection from an eriophyid mite. Plant Dis. 2021, 105, 1234. [Google Scholar] [CrossRef]
- Dolja, V.V.; Krupovic, M.; Koonin, E.V. Deep roots and splendid boughs of the global plant virome. Annu. Rev. Phytopathol. 2020, 58, 23–53. [Google Scholar] [CrossRef]
- Zhou, X.; Lin, W.; Sun, K.; Wang, S.; Zhou, X.; Jackson, A.O.; Li, Z. Specificity of plant rhabdovirus cell-to-cell movement. J. Virol. 2019, 93, e00296-19. [Google Scholar] [CrossRef] [Green Version]
- Guterres, A.; de Oliveira, R.C.; Fernandes, J.; de Lemos, E.R.S.; Schrago, C.G. New bunya-like viruses: Highlighting their relations. Infect. Genet. Evol. 2017, 49, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C. Molecular clocks and the puzzle of RNA virus origins. J. Virol. 2003, 77, 3893–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabbidge, L.O.; Blouin, A.G.; Chooi, K.M.; Higgins, C.M.; MacDiarmid, R.M. Characterisation and Distribution of Karaka Ōkahu Purepure Virus—A Novel Emaravirus Likely to Be Endemic to New Zealand. Viruses 2021, 13, 1611. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081611
Rabbidge LO, Blouin AG, Chooi KM, Higgins CM, MacDiarmid RM. Characterisation and Distribution of Karaka Ōkahu Purepure Virus—A Novel Emaravirus Likely to Be Endemic to New Zealand. Viruses. 2021; 13(8):1611. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081611
Chicago/Turabian StyleRabbidge, Lee O., Arnaud G. Blouin, Kar Mun Chooi, Colleen M. Higgins, and Robin M. MacDiarmid. 2021. "Characterisation and Distribution of Karaka Ōkahu Purepure Virus—A Novel Emaravirus Likely to Be Endemic to New Zealand" Viruses 13, no. 8: 1611. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081611