
Virological and Epidemiological Features of Norovirus Infections in Brazil, 2017–2018

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Stool Collection and Ethics Statements
2.2. Viral RNA Extraction
2.3. Norovirus Detection and Quantification
2.4. Genotyping and Sequencing
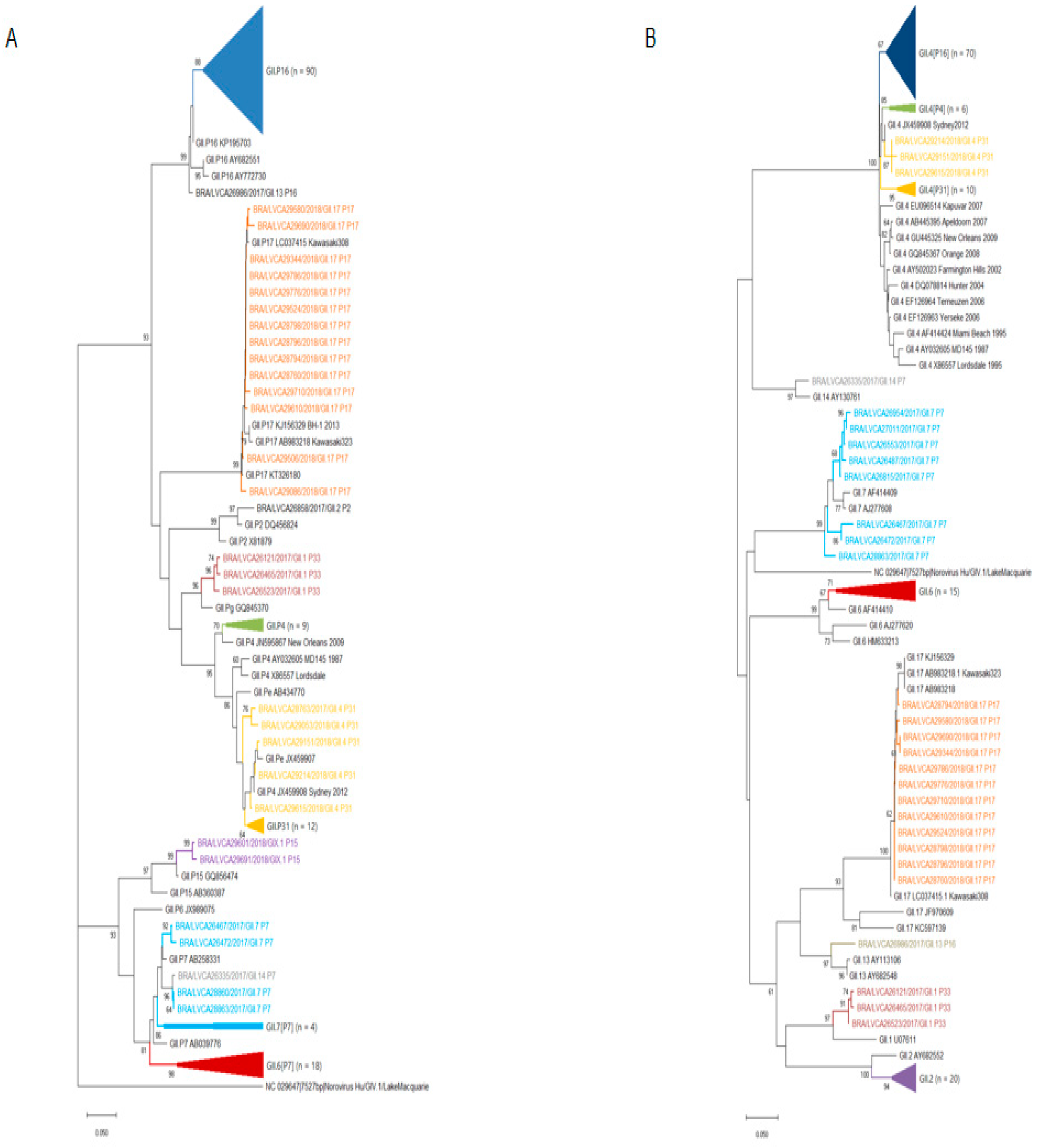
2.5. Phylogenetic Analysis
2.6. Statistical Analysis
3. Results
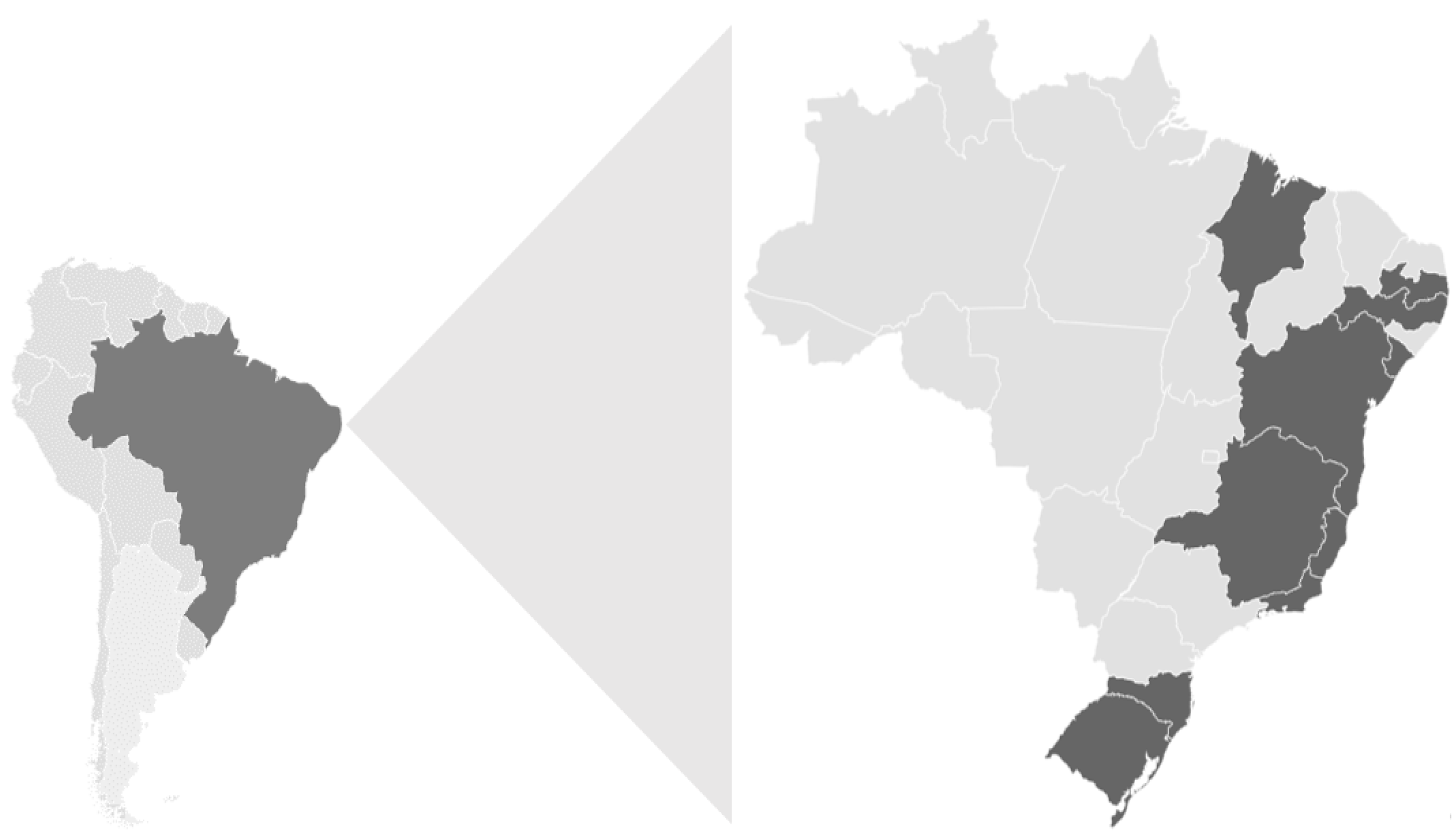
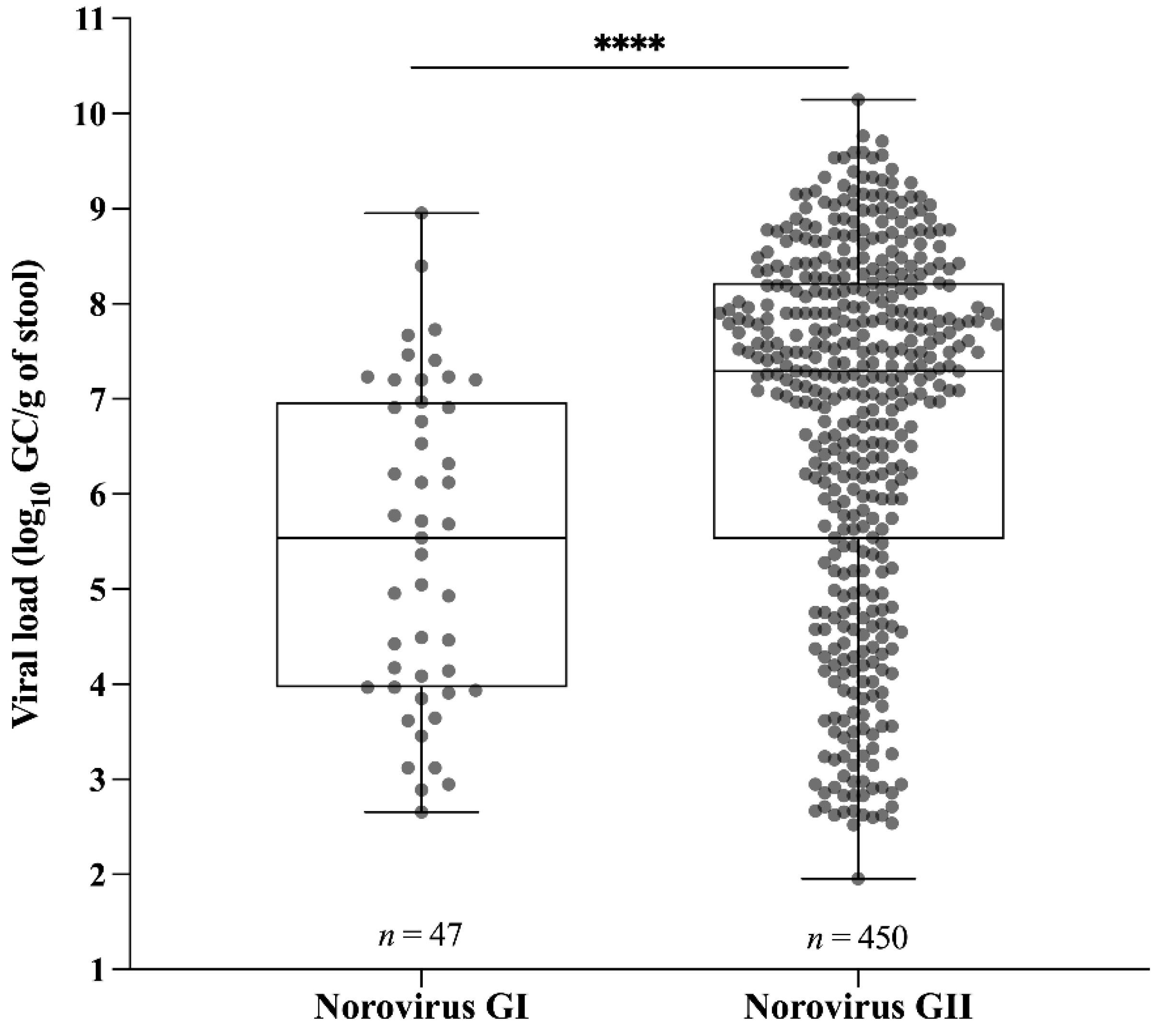
3.1. Norovirus Epidemiology
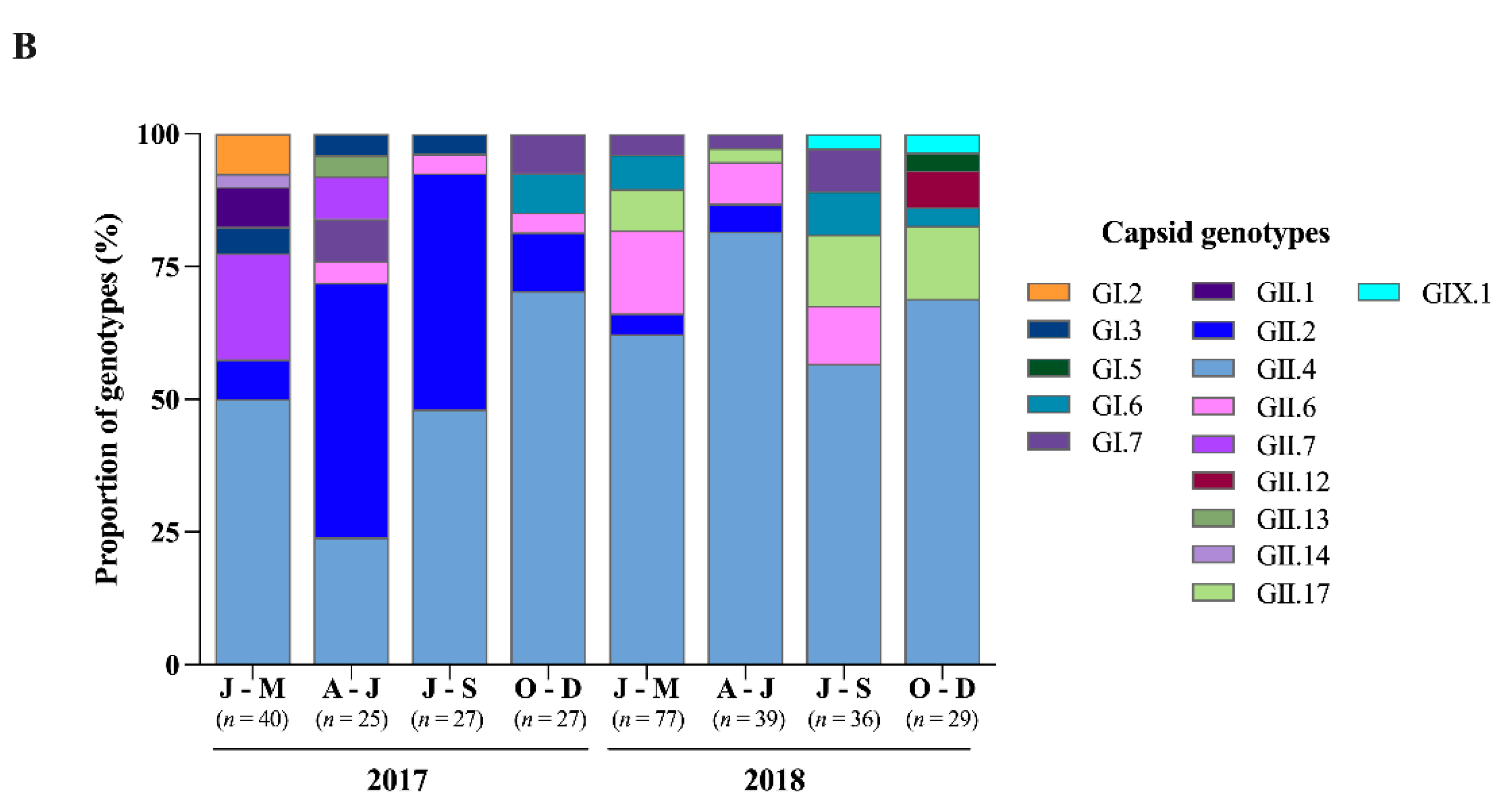
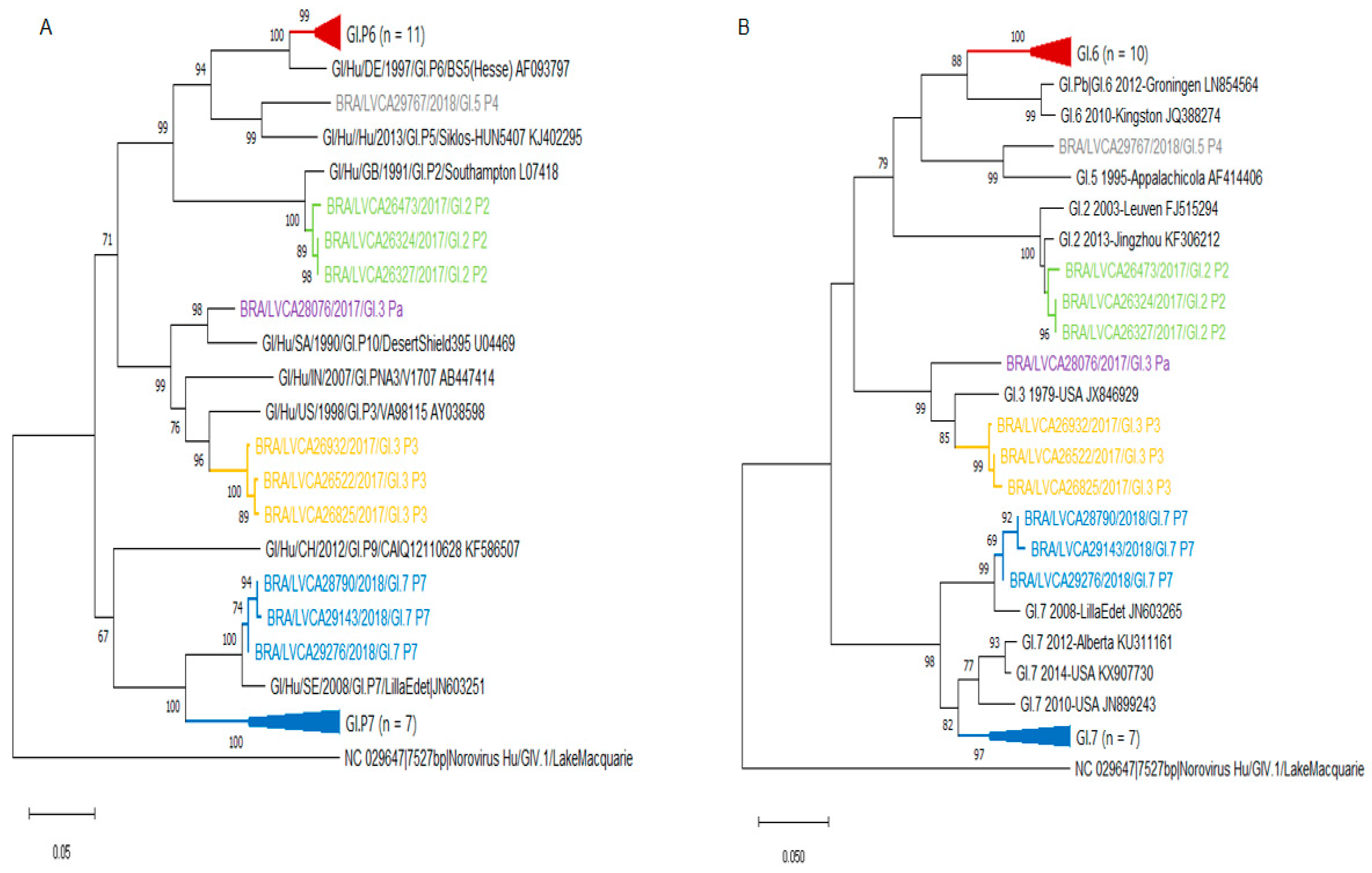
3.2. Norovirus Genotyping
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, C.L.F.; Aryee, M.J.; Boschi-Pinto, C.; Black, R.E. Estimating Diarrhea Mortality among Young Children in Low and Middle Income Countries. PLoS ONE 2012, 7, e29151. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Hall, A.J.; E. Robinson, A.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Koopmans, M.; A. Lopman, B. Global prevalence of norovirus in cases of gastroenteritis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Lopman, B.A.; Steele, D.; Kirkwood, C.D.; Parashar, U.D. The Vast and Varied Global Burden of Norovirus: Prospects for Prevention and Control. PLoS Med. 2016, 13, e1001999. [Google Scholar] [CrossRef] [PubMed]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Bucardo, F.; Reyes, Y.; Svensson, L.; Nordgren, J. Predominance of Norovirus and Sapovirus in Nicaragua after Implementation of Universal Rotavirus Vaccination. PLoS ONE 2014, 9, e98201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAtee, C.; Challappa, R.; Saito, M.; Mejia, C.; Bern, C.; Webman, R.; Pajuelo, M.; Gilman, R.H.; Soria, R.; Ribera, J.P.; et al. Burden of Norovirus and Rotavirus in Children After Rotavirus Vaccine Introduction, Cochabamba, Bolivia. Am. J. Trop. Med. Hyg. 2016, 94, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A.J.; Eisenbart, V.G.; Etingüe, A.L.; Gould, L.H.; Lopman, B.A.; Parashar, U.D. Epidemiology of Foodborne Norovirus Outbreaks, United States, 2001–2008. Emerg. Infect. Dis. 2012, 18, 1566–1573. [Google Scholar] [CrossRef]
- Green, K. Caliciviridae: The noroviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Racaniello, V.R., Roizman, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1, pp. 586–608. [Google Scholar]
- Chhabra, P.; De Graaf, M.; Parra, G.I.; Chan, M.C.-W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef]
- Siebenga, J.J.; Vennema, H.; Zheng, D.; Vinje, J.; Lee, B.E.; Pang, X.; Ho, E.C.M.; Lim, W.; Choudekar, A.; Broor, S.; et al. Norovirus Illness Is a Global Problem: Emergence and Spread of Norovirus GII.4 Variants, 2001–2007. J. Infect. Dis. 2009, 200, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Parra, G.I.; Squires, R.B.; Karangwa, C.K.; Johnson, J.A.; Lepore, C.J.; Sosnovtsev, S.V.; Green, K.Y. Static and Evolving Norovirus Genotypes: Implications for Epidemiology and Immunity. PLoS Pathog. 2017, 13, e1006136. [Google Scholar] [CrossRef] [PubMed]
- Eden, J.-S.; Hewitt, J.; Lim, K.L.; Boni, M.F.; Merif, J.; Greening, G.; Ratcliff, R.M.; Holmes, E.C.; Tanaka, M.M.; Rawlinson, W.D.; et al. The emergence and evolution of the novel epidemic norovirus GII.4 variant Sydney 2012. Virology 2013, 450–451, 106–113. [Google Scholar] [CrossRef]
- van Beek, J.; de Graaf, M.; Al-Hello, H.; Allen, D.J.; Ambert-Balay, K.; Botteldoorn, N.; Brytting, M.; Buesa, J.; Cabrerizo, M.; Chan, M.; et al. Molecular surveillance of norovirus, 2005–2016: An epidemiological analysis of data collected from the NoroNet network. Lancet Infect. Dis. 2018, 18, 545–553. [Google Scholar] [CrossRef]
- Ruis, C.; Lindesmith, L.C.; Mallory, M.L.; Brewer-Jensen, P.D.; Bryant, J.M.; Costantini, V.; Monit, C.; Vinjé, J.; Baric, R.S.; Goldstein, R.A.; et al. Preadaptation of pandemic GII.4 noroviruses in unsampled virus reservoirs years before emergence. Virus Evol. 2020, 6, veaa067. [Google Scholar] [CrossRef]
- Lu, J.; Sun, L.; Fang, L.; Yang, F.; Mo, Y.; Lao, J.; Zheng, H.; Tan, X.; Lin, H.; Rutherford, S.; et al. Gastroenteritis Outbreaks Caused by Norovirus GII.17, Guangdong Province, China, 2014–2015. Emerg. Infect. Dis. 2015, 21, 1240–1242. [Google Scholar] [CrossRef] [PubMed]
- Koo, E.S.; Kim, M.S.; Choi, Y.S.; Park, K.-S.; Jeong, Y.S. Occurrence of novel GII.17 and GII.21 norovirus variants in the coastal environment of South Korea in 2015. PLoS ONE 2017, 12, e0172237. [Google Scholar] [CrossRef]
- Bidalot, M.; Théry, L.; Kaplon, J.; De Rougemont, A.; Ambert-Balay, K. Emergence of new recombinant noroviruses GII.p16-GII.4 and GII.p16-GII.2, France, winter 2016 to 2017. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, J.L.; Barclay, L.; Collins, N.R.; Wikswo, M.E.; Castro, C.J.; Magaña, L.C.; Gregoricus, N.; Marine, R.; Chhabra, P.; Vinjé, J. Genetic and Epidemiologic Trends of Norovirus Outbreaks in the United States from 2013 to 2016 Demonstrated Emergence of Novel GII.4 Recombinant Viruses. J. Clin. Microbiol. 2017, 55, 2208–2221. [Google Scholar] [CrossRef] [Green Version]
- Medici, M.C.; Tummolo, F.; Martella, V.; De Conto, F.; Arcangeletti, M.C.; Pinardi, F.; Ferraglia, F.; Chezzi, C.; Calderaro, A. Emergence of novel recombinant GII.P16_GII.2 and GII. P16_GII.4 Sydney 2012 norovirus strains in Italy, winter 2016/2017. New Microbiol. 2018, 41, 71–72. [Google Scholar]
- Chan, M.C.-W.; Roy, S.; Bonifacio, J.; Zhang, L.-Y.; Chhabra, P.; Chan, J.C.; Celma, C.; Igoy, M.A.; Lau, S.-L.; Mohammad, K.N.; et al. Detection of Norovirus Variant GII.4 Hong Kong in Asia and Europe, 2017−2019. Emerg. Infect. Dis. 2021, 27, 289–293. [Google Scholar] [CrossRef]
- Cantelli, C.P.; da Silva, M.F.M.; Fumian, T.M.; da Cunha, D.C.; Andrade, J.D.S.R.; Malta, F.C.; da Silva, E.; Mouta Junior, S.; Fialho, A.M.; de Moraes, M.T.B.; et al. High genetic diversity of noroviruses in children from a community-based study in Rio de Janeiro, Brazil, 2014–2018. Arch Virol. 2019, 164, 1427–1432. [Google Scholar] [CrossRef]
- Hernandez, J.M.; Silva, L.D.; Junior, E.C.S.; Bandeira, R.S.; Rodrigues, E.A.M.; Lucena, M.S.S.; Costa, S.T.P.; Gabbay, Y.B. Molecular epidemiology and temporal evolution of norovirus associated with acute gastroenteritis in Amazonas state, Brazil. BMC Infect. Dis. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gondim, R.D.; Pankov, R.C.; Prata, M.M.; Medeiros, P.H.; Veras, H.N.; Santos, A.K.; Magalhães, L.M.; Havt, A.; Fumian, T.M.; Miagostovich, M.P.; et al. Genetic Diversity of Norovirus Infections, Coinfections, and Undernutrition in Children From Brazilian Semiarid Region. J. Pediatr. Gastroenterol. Nutr. 2018, 67, e117–e122. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, T.; Kojima, S.; Shinohara, M.; Uchida, K.; Fukushi, S.; Hoshino, F.B.; Takeda, N.; Katayama, K. Broadly Reactive and Highly Sensitive Assay for Norwalk-Like Viruses Based on Real-Time Quantitative Reverse Transcription-PCR. J. Clin. Microbiol. 2003, 41, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Kojima, S.; Kageyama, T.; Fukushi, S.; Hoshino, F.B.; Shinohara, M.; Uchida, K.; Natori, K.; Takeda, N.; Katayama, K. Genogroup-specific PCR primers for detection of Norwalk-like viruses. J. Virol. Methods 2001, 100, 107–114. [Google Scholar] [CrossRef]
- Beuret, C.; Kohler, D.; Baumgartner, A.; Lüthi, T.M. Norwalk-Like Virus Sequences in Mineral Waters: One-Year Monitoring of Three Brands. Appl. Environ. Microbiol. 2002, 68, 1925–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Battistuzzi, F.U. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Safadi, M.A.; Riera-Montes, M.; Bravo, L.; Tangsathapornpong, A.; Lagos, R.; Thisyakorn, U.; Linhares, A.C.; Capeding, R.; Prommalikit, O.; Verstraeten, T.; et al. The burden of norovirus disease in children: A multi-country study in Chile, Brazil, Thailand and the Philippines. Int. J. Infect. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Reymão, T.K.A.; Fumian, T.M.; Justino, M.C.A.; Hernandez, J.M.; Bandeira, R.S.; Lucena, M.S.S.; Teixeira, D.M.; Farias, F.P.; Silva, L.D.; Linhares, A.C.; et al. Norovirus RNA in serum associated with increased fecal viral load in children: Detection, quantification and molecular analysis. PLoS ONE 2018, 13, e0199763. [Google Scholar] [CrossRef]
- Da Costa, S.T.P.; Fumian, T.M.; De Lima, I.C.G.; Siqueira, J.A.M.; Da Silva, L.D.; Hernández, J.D.M.; De Lucena, M.S.S.; Reymão, T.K.A.; Soares, L.; Mascarenhas, J.D.P.; et al. High prevalence of norovirus in children with sporadic acute gastroenteritis in Manaus, Amazon Region, northern Brazil. Memórias Inst. Oswaldo Cruz 2017, 112, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamioka, G.A.; Madalosso, G.; Pavanello, E.I.; Sousa, S.C.Z.; Bassit, N.P.; Sato, A.P.S. Norovirus in Sao Paulo city, Brazil, 2010–2016: A cross-sectional study of the leading cause of gastroenteritis in children. Epidemiol. Serv. Saude 2019, 28, e2018290. [Google Scholar] [CrossRef] [Green Version]
- Hemming, M.; Räsänen, S.; Huhti, L.; Paloniemi, M.; Salminen, M.; Vesikari, T. Major reduction of rotavirus, but not norovirus, gastroenteritis in children seen in hospital after the introduction of RotaTeq vaccine into the National Immunization Programme in Finland. Eur. J. Nucl. Med. Mol. Imaging 2013, 172, 739–746. [Google Scholar] [CrossRef]
- Payne, D.C.; Vinje, J.; Szilagyi, P.G.; Edwards, K.M.; Staat, M.A.; Weinberg, G.; Hall, C.B.; Chappell, J.; Bernstein, D.I.; Curns, A.T.; et al. Norovirus and Medically Attended Gastroenteritis in U.S. Children. N. Engl. J. Med. 2013, 368, 1121–1130. [Google Scholar] [CrossRef] [Green Version]
- Becker-Dreps, S.; Bucardo, F.; Vilchez, S.; Zambrana, L.E.; Liu, L.; Weber, D.J.; Peña, R.; Barclay, L.; Vinjé, J.; Hudgens, M.G.; et al. Etiology of childhood diarrhea after rotavirus vaccine introduction: A prospective, population-based study in Nicaragua. Pediatr. Infect. Dis. J. 2014, 33, 1156–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.P.; Zhou, S.X.; Wang, X.; Lu, Q.B.; Shi, L.S.; Ren, X.; Zhang, H.Y.; Wang, Y.F.; Lin, S.H.; Zhang, C.H. Chinese Centers for Disease Control and Prevention (CDC) Etiology of Diarrhea Surveillance Study Team. Etiological, epidemiological, and clinical features of acute diarrhea in China. Nat. Commun. 2021, 12, 2464. [Google Scholar] [CrossRef]
- Bucardo, F.; Reyes, Y.; Becker-Dreps, S.; Bowman, N.; Gruber, J.F.; Vinjé, J.; Espinoza, F.; Paniagua, M.; Balmaseda, A.; Svensson, L.; et al. Pediatric norovirus GII.4 infections in Nicaragua, 1999–2015. Infect. Genet. Evol. 2017, 55, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.; Guo, J.; Wei, Z.; Huang, Z.; Wang, C.; Qiu, Y.; Xu, X.; Zeng, M. Aetiology of acute diarrhoea in children in Shanghai, 2015–2018. PLoS ONE 2021, 16, e0249888. [Google Scholar] [CrossRef]
- Chen, C.; Yan, J.B.; Wang, H.L.; Li, P.; Li, K.F.; Wu, B.; Zhang, H. Molecular epidemiology and spatiotemporal dynamics of norovirus associated with sporadic acute gastroenteritis during 2013–2017, Zhoushan Islands, China. PLoS ONE 2018, 13, e0200911. [Google Scholar] [CrossRef] [PubMed]
- Howard, L.M.; Mwape, I.; Siwingwa, M.; Simuyandi, M.; Guffey, M.B.; Stringer, J.S.A.; Chi, B.H.; Edwards, K.M.; Chilengi, R. Norovirus infections in young children in Lusaka Province, Zambia: Clinical characteristics and molecular epidemiology. BMC Infect. Dis. 2017, 17, 92. [Google Scholar] [CrossRef] [Green Version]
- Siafakas, N.; Zerva, L.; Hatzaki, D.; Lebessi, E.; Chronopoulou, G.; Paraskakis, I.; Pournaras, S. Molecular epidemiology of noroviruses in children in South Greece, 2013–2015. J. Med. Virol. 2018, 90, 1703–1711. [Google Scholar] [CrossRef]
- Lucero, Y.; Lagomarcino, A.J.; Espinoza, M.; Kawakami, N.; Mamani, N.; Huerta, N.; Del Canto, F.; Farfán, M.; Sawaguchi, Y.; George, S.; et al. Norovirus compared to other relevant etiologies of acute gastroenteritis among families from a semirural county in Chile. Int. J. Infect. Dis. 2020, 101, 353–360. [Google Scholar] [CrossRef]
- Santos, V.S.; Gurgel, R.Q.; Cavalcante, S.M.; Kirby, A.; Café, L.P.; Souto, M.J.; Dolabella, S.S.; de Assis, M.R.; Fumian, T.M.; Miagostovich, M.P.; et al. Acute norovirus gastroenteritis in children in a highly rotavirus-vaccinated population in Northeast Brazil. J. Clin. Virol. 2017, 88, 33–38. [Google Scholar] [CrossRef]
- Fioretti, J.M.; Ferreira, M.S.R.; Victoria, M.; Vieira, C.; Xavier, M.; Leite, J.P.G.; Miagostovich, M.P. Genetic diversity of noroviruses in Brazil. Memórias do Instituto Oswaldo Cruz 2011, 106, 942–947. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, J.d.S.; Rocha, M.S.; Carvalho-Costa, F.A.; Fioretti, J.M.; Xavier, M.d.P.; Nunes, Z.M.; Cardoso, J.; Fialho, A.M.; Leite, J.P.; Miagostovich, M.P. Noroviruses associated with outbreaks of acute gastroenteritis in the State of Rio Grande do Sul, Brazil, 2004–2011. J. Clin. Virol. 2014, 61, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karst, S.M. Pathogenesis of noroviruses, emerging RNA viruses. Viruses 2010, 2, 748–781. [Google Scholar] [CrossRef]
- Chan, M.C.; Sung, J.J.; Lam, R.K.; Chan, P.K.; Lee, N.L.; Lai, R.W.; Leung, W.K. Fecal viral load and norovirus-associated gastroenteritis. Emerg. Infect. Dis. 2006, 12, 1278–1280. [Google Scholar] [CrossRef]
- Shioda, K.; Barclay, L.; Becker-Dreps, S.; Bucardo-Rivera, F.; Cooper, P.J.; Payne, D.C.; Vinjé, J.; Lopman, B.A. Can Use of Viral Load Improve Norovirus Clinical Diagnosis and Disease Attribution? Open Forum Infect. Dis. 2017, 4, ofx131. [Google Scholar] [CrossRef]
- Lopman, B.; Armstrong, B.; Atchison, C.; Gray, J.J. Host, Weather and Virological Factors Drive Norovirus Epidemiology: Time-Series Analysis of Laboratory Surveillance Data in England and Wales. PLoS ONE 2009, 4, e6671. [Google Scholar] [CrossRef]
- Wollants, E.; De Coster, S.; Van Ranst, M.; Maes, P. A decade of norovirus genetic diversity in Belgium. Infect. Genet. Evol. 2015, 30, 37–44. [Google Scholar] [CrossRef]
- Gómez-Santiago, F.; Ribas-Aparicio, R.M.; García-Lozano, H. Molecular characterization of human calicivirus associated with acute diarrheal disease in mexican children. Virol. J. 2012, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Esteves, A.; Nordgren, J.; Tavares, C.; Fortes, F.; Dimbu, R.; Saraiva, N.; Istrate, C. Genetic diversity of norovirus in children under 5 years of age with acute gastroenteritis from Angola. Epidemiol. Infect. 2018, 146, 551–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirwati, H.; Donato, C.M.; Mawarti, Y.; Mulyani, N.S.; Ikram, A.; Aman, A.T.; Peppelenbosch, M.P.; Soenarto, Y.; Pan, Q.; Hakim, M.S. Norovirus and rotavirus infections in children less than five years of age hospitalized with acute gastroenteritis in Indonesia. Arch. Virol. 2019, 164, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Lartey, B.L.; Quaye, O.; Damanka, S.A.; Agbemabiese, C.A.; Armachie, J.; Dennis, F.E.; Enweronu-Laryea, C.; Armah, G.E. Understanding Pediatric Norovirus Epidemiology: A Decade of Study among Ghanaian Children. Viruses 2020, 12, 1321. [Google Scholar] [CrossRef]
- Niendorf, S.; Faber, M.; Tröger, A.; Hackler, J.; Jacobsen, S. Diversity of Noroviruses throughout Outbreaks in Germany 2018. Viruses 2020, 12, 1157. [Google Scholar] [CrossRef]
- Shioda, K.; Kambhampati, A.; Hall, A.J.; Lopman, B.A. Global age distribution of pediatric norovirus cases. Vaccine 2015, 33, 4065–4068. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Jung, J.; Oh, S.; Jung, H.; Oh, Y.; Cho, S.; Cho, S.; Cho, S.; Park, H.; Jo, N.; et al. Characterization of norovirus infections in Seoul, Korea. Microbiol. Immunol. 2012, 56, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Bruggink, L.D.; Dunbar, N.L.; Marshall, J.A. Norovirus genotype diversity associated with gastroenteritis outbreaks in aged-care facilities. Epidemiol. Infect. 2015, 143, 3064–3068. [Google Scholar] [CrossRef]
- Mugyia, A.E.; Ndze, V.; Akoachere, J.-F.T.K.; Browne, H.; Boula, A.; Ndombo, P.K.; Cannon, J.L.; Vinjé, J.; Ndip, L.M. Molecular epidemiology of noroviruses in children under 5 years of age with acute gastroenteritis in Yaoundé, Cameroon. J. Med. Virol. 2018, 91, 738–743. [Google Scholar] [CrossRef]
- Alam, A.C.; Qureshi, S.A.; Vinjé, J.; Zaidi, A.; Quresh, S.A. Genetic characterization of norovirus strains in hospitalized children from Pakistan. J. Med. Virol. 2015, 88, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Marques Mendanha de Oliveira, D.; Souza, M.; Souza Fiaccadori, F.; César Pereira Santos, H.; das Dôres de Paula Cardoso, D. Monitoring of Calicivirus among day-care children: Evidence of asymptomatic viral excretion and first report of GI.7 Norovirus and GI.3 Sapovirus in Brazil. J. Med. Virol. 2014, 86, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, Y.; Shimizu, T.; Ishikawa, M.; Komane, A.; Okabe, N.; Ryo, A.; Kimura, H.; Katayama, K.; Shimizu, H. Complete Genome Sequence of a Recombinant GII.P16-GII.4 Norovirus Detected in Kawasaki City, Japan, in 2016. Genome Announc. 2016, 4, e01099-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.S.; Koo, E.S.; Kim, M.S.; Choi, J.D.; Shin, Y.; Jeong, Y.S. Re-emergence of a GII.4 Norovirus Sydney 2012 Variant Equipped with GII.P16 RdRp and Its Predominance over Novel Variants of GII.17 in South Korea in 2016. Food Environ. Virol. 2017, 9, 168–178. [Google Scholar] [CrossRef]
- Niendorf, S.; Jacobsen, S.; Faber, M.; Eis-Hübinger, A.M.; Hofmann, J.; Zimmermann, O.; Höhne, M.; Bock, C.-T. Steep rise in norovirus cases and emergence of a new recombinant strain GII.P16-GII.2, Germany, winter 2016. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef]
- Lun, J.H.; Hewitt, J.; Sitabkhan, A.; Eden, J.S.; Enosi Tuipulotu, D.; Netzler, N.E.; Morrell, L.; Merif, J.; Jones, R.; Huang, B.; et al. Emerging recombinant noroviruses identified by clinical and waste water screening. Emerg. Microbes Infect. 2018, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Kuang, X.; Teng, Z.; Zhang, X. Genotypic prevalence of norovirus GII in gastroenteritis outpatients in Shanghai from 2016 to 2018. Gut Pathog. 2019, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cheung, S.K.; Kwok, K.; Zhang, L.-Y.; Mohammad, K.N.; Lui, C.Y.G.; Lee, N.; Nelson, E.A.S.; Lai, R.W.; Leung, T.F.; Chan, P.K.; et al. Higher Viral Load of Emerging Norovirus GII.P16-GII.2 than Pandemic GII.4 and Epidemic GII.17, Hong Kong, China. Emerg. Infect. Dis. 2018, 25, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohma, K.; Lepore, C.J.; Siltz, L.; Parra, G.I. Phylogenetic Analyses Suggest that Factors Other Than the Capsid Protein Play a Role in the Epidemic Potential of GII.2 Norovirus. mSphere 2017, 2, e00187-17. [Google Scholar] [CrossRef] [Green Version]
- Bull, R.A.; Eden, J.S.; Rawlinson, W.D.; White, P.A. Rapid evolution of pandemic noroviruses of the GII.4 lineage. PLoS Pathog. 2010, 6, e1000831. [Google Scholar] [CrossRef]
- Tran, T.H.; Trainor, E.; Nakagomi, T.; Cunliffe, N.; Nakagomi, O. Molecular epidemiology of noroviruses associated with acute sporadic gastroenteritis in children: Global distribution of genogroups, genotypes and GII.4 variants. J. Clin. Virol. 2013, 56, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Bruggink, L.D.; Moselen, J.M.; Marshall, J.A. Genotype analysis of noroviruses associated with gastroenteritis outbreaks in childcare centres, Victoria, Australia, 2012–2015. Epidemiol. Infect. 2017, 145, 1933–1941. [Google Scholar] [CrossRef] [Green Version]
- Diakoudi, G.; Lanave, G.; Catella, C.; Medici, M.C.; De Conto, F.; Calderaro, A.; Loconsole, D.; Chironna, M.; Bonura, F.; Giammanco, G.M.; et al. Analysis of GII.P7 and GII.6 noroviruses circulating in Italy during 2011–2016 reveals a replacement of lineages and complex recombination history. Infect. Genet. Evol. 2019, 75, 103991. [Google Scholar] [CrossRef] [PubMed]
- Fumian, T.M.; Andrade, J.D.S.R.D.; Leite, J.P.G.; Miagostovich, M.P. Norovirus Recombinant Strains Isolated from Gastroenteritis Outbreaks in Southern Brazil, 2004–2011. PLoS ONE 2016, 11, e0145391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, M.C.-W.; Lee, N.; Hung, T.-N.; Kwok, K.; Cheung, K.; Tin, E.K.Y.; Lai, R.W.M.; Nelson, E.A.S.; Leung, T.F.; Chan, P.K.S. Rapid emergence and predominance of a broadly recognizing and fast-evolving norovirus GII.17 variant in late 2014. Nat. Commun. 2015, 6, 10061. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, Y.; Ishikawa, M.; Shimizu, T.; Komane, A.; Kasuo, S.; Shinohara, M.; Nagasawa, K.; Kimura, H.; Ryo, A.; Okabe, N.; et al. Genetic analyses of GII.17 norovirus strains in diarrheal disease outbreaks from December 2014 to March 2015 in Japan reveal a novel polymerase sequence and amino acid substitutions in the capsid region. Eurosurveillance 2015, 20, 21173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Graaf, M.; van Beek, J.; Vennema, H.; Podkolzin, A.T.; Hewitt, J.; Bucardo, F.; Templeton, K.; Mans, J.; Nordgren, J.; Reuter, G.; et al. Emergence of a novel GII.17 norovirus – End of the GII.4 era? Eurosurveillance 2015, 20, 21178. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.C.W.; Hu, Y.; Chen, H.; Podkolzin, A.T.; Zaytseva, E.V.; Komano, J.; Sakon, N.; Poovorawan, Y.; Vongpunsawad, S.; Thanusuwannasak, T.; et al. Global Spread of Norovirus GII.17 Kawasaki 308, 2014–2016. Emerg. Infect. Dis. 2017, 23, 1350–1354. [Google Scholar] [CrossRef] [PubMed]
- das Neves Costa, L.C.P.; Teixeira, D.M.; Portela, A.C.R.; de Lima, I.C.G.; da Silva Bandeira, R.; Sousa Júnior, E.C.; Siqueira, J.A.M.; Resque, H.R.; da Silva, L.D.; Gabbay, Y.B. Molecular and evolutionary characterization of norovirus GII.17 in the northern region of Brazil. BMC Infect. Dis. 2019, 19, 1021. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.S.; Fumian, T.M.; Leite, J.P.G.; de Assis, M.R.; Bello, G.; Mir, D.; Miagostovich, M.P. Detection and molecular characterization of emergent GII.P17/GII.17 Norovirus in Brazil, 2015. Infect. Genet. Evol. 2017, 51, 28–32. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, Z.; Li, S.; Wu, Q.; Tian, P.; Zhang, Z.; Wang, D. Fingerprinting of human noroviruses co-infections in a possible foodborne outbreak by metagenomics. Int. J. Food Microbiol. 2020, 333, 108787. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region/ State | N° of Fecal Samples–Positive/Tested (%) | p-Value 1 (Chi-Square Test) | ||
|---|---|---|---|---|
| Total % of GI–GII | 2017 | 2018 | ||
| Southeastern | 94/274 (34.3) 12.8–87.2 | 27/106 (25.5) | 67/168 (39.8) | 0.0144 |
| Espírito Santo | 6/24 | 22/56 | ||
| Minas Gerais | 11/52 | 38/75 | ||
| Rio de Janeiro | 10/30 | 7/37 | ||
| Northeastern | 149/613(24.3) 7.4–92.6 | 64/361 (17.7) | 85/252 (33.7) | <0.0001 |
| Bahia | 22/74 | 26/98 | ||
| Maranhão | 17/55 | 3/8 | ||
| Paraíba | 3/25 | 0/37 | ||
| Pernambuco | 8/150 | 31/68 | ||
| Sergipe | 14/57 | 25/41 | ||
| Southern | 254/659 (38.5) 12.2–87.8 | 99/278 (35.6) | 155/381 (40.6) | 0.1865 |
| Rio Grande do Sul | 33/131 | 50/168 | ||
| Santa Catarina | 66/147 | 105/213 | ||
| Total | 497/1,546 (32.1) 10.9–89.1 | 190/745 (25.5) | 307/801(38.3) | <0.0001 |
| Age Group (Months) | N° of Fecal Samples–Positive/Tested (%) | p-Value 1 (Chi-Square Test) | ||
|---|---|---|---|---|
| 2017 | 2018 | Total | ||
| 0–6 | 28/118 (23.7) | 24/123 (19.5) | 52/241 (21.5) | <0.0001 |
| >6–12 | 47/122 (38.5) | 66/133 (49.6) | 113/255 (44.3) | - |
| >12–24 | 54/174 (31.0) | 104/203 (51.2) | 158/377 (41.9) | 0.9742 |
| >24–60 | 30/164 (18.3) | 44/141 (31.2) | 74/305 (24.2) | <0.0001 |
| >60 | 31/167 (18.5) | 69/201 (34.3) | 100/368 (27.1) | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarmento, S.K.; de Andrade, J.d.S.R.; Miagostovich, M.P.; Fumian, T.M. Virological and Epidemiological Features of Norovirus Infections in Brazil, 2017–2018. Viruses 2021, 13, 1724. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091724
Sarmento SK, de Andrade JdSR, Miagostovich MP, Fumian TM. Virological and Epidemiological Features of Norovirus Infections in Brazil, 2017–2018. Viruses. 2021; 13(9):1724. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091724
Chicago/Turabian StyleSarmento, Sylvia Kahwage, Juliana da Silva Ribeiro de Andrade, Marize Pereira Miagostovich, and Tulio Machado Fumian. 2021. "Virological and Epidemiological Features of Norovirus Infections in Brazil, 2017–2018" Viruses 13, no. 9: 1724. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091724