
Recent Advances in Developing Treatments of Kaposi’s Sarcoma Herpesvirus-Related Diseases


1
Institute of Virology, Hannover Medical School, 30625 Hannover, Germany
2
German Centre for Infection Research, Hannover-Braunschweig Site, 38023 Braunschweig, Germany
3
Cluster of Excellence 2155 RESIST, Institute of Virology, Hannover Medical School, 30625 Hannover, Germany
*
Author to whom correspondence should be addressed.
Viruses 2021, 13(9), 1797; https://0-doi-org.brum.beds.ac.uk/10.3390/v13091797
Submission received: 30 July 2021
/
Revised: 30 August 2021
/
Accepted: 1 September 2021
/
Published: 9 September 2021
(This article belongs to the Special Issue Anti-herpesvirus Drugs and Vaccines)
Abstract
:Kaposi-sarcoma-associated herpesvirus (KSHV) or human herpesvirus 8 (HHV-8) is the causative agent of several malignancies, including Kaposi’s sarcoma (KS), primary effusion lymphoma (PEL), and multicentric Castleman’s disease (MCD). Active KSHV replication has also been associated with a pathological condition called KSHV inflammatory cytokine syndrome (KICS), and KSHV may play a role in rare cases of post-transplant polyclonal lymphoproliferative disorders. Several commonly used herpesviral DNA polymerase inhibitors are active against KSHV in tissue culture. Unfortunately, they are not always efficacious against KSHV-induced diseases. To improve the outcome for the patients, new therapeutics need to be developed, including treatment strategies that target either viral proteins or cellular pathways involved in tumor growth and/or supporting the viral life cycle. In this review, we summarize the most commonly established treatments against KSHV-related diseases and review recent developments and promising new compounds that are currently under investigation or on the way to clinical use.
{kind=link}
1. Introduction
Kaposi’s sarcoma-associated herpesvirus (KSHV) is a double-stranded DNA virus, discovered in 1994 by Patrick Moore and Yuan Chang and classified as a human gamma2-herpesvirus [1]. KSHV is associated with three neoplastic diseases: Kaposi’s sarcoma (KS) [1], multicentric Castleman’s disease (MCD) [2] and primary effusion lymphoma (PEL) [3]; As reviewed elsewhere, it meets the epidemiological and basic research requirements for recognition as an oncogenic agent and has thus been classified as a class I human carcinogen by the International Agency for Research against Cancer (IARC) [4,5]. Additionally, KSHV has also been associated with a pathological condition called KSHV inflammatory cytokine syndrome (KICS) [6] and rare cases of post-transplant polyclonal lymphoproliferative disorders [7,8] as well as with cases of plasmablastic lymphoma emerging from MCD [9].
The histopathological features of KS include atypical vascularization and neoangiogenesis with extensive infiltrates of inflammatory cells and the proliferation of atypical endothelial spindle cells [10]. Based on epidemiological criteria, KS has been classified into four different forms that are histologically similar [11]. Moritz Kaposi described KS for the first time in 1872 as a rare tumor endemic in the Mediterranean area that mostly affected middle-aged and older men. This “classic” KS is characterized by indolent skin lesions and viscera are only rarely involved [12].
In East and Central Africa, a more aggressive form of KS (the “endemic African” KS) in HIV-negative patients involves internal organs and lymph nodes in children and adults [13,14]. The “iatrogenic” KS affects up to 25% (in KSHV endemic regions) of transplant recipients under immunosuppressive therapy who were previously infected with KSHV [15,16]. This form of KS often recedes after the interruption of the immunosuppressive therapy [17,18].
In the 1980s, following the emergence of HIV/AIDS, the AIDS-associated or epidemic KS became a common manifestation among HIV-infected people [19]. AIDS-KS is the most aggressive form of KS. It can involve the lungs and the gastrointestinal tract [20]. After the introduction of the antiretroviral combination therapy (ART) against HIV, the incidence of this variant of KS was significantly reduced [19,21,22,23,24].
Primary effusion lymphoma (PEL) or body cavity-based lymphoma (BCBL) is a B cell non-Hodgkin lymphoma characterized by lymphomatous effusions in the pleural, pericardial and peritoneal body cavities [25]. PEL often arises in HIV-infected individuals and it is estimated to account for 2–5% of HIV-associated lymphomas [26]. PEL can also occur in transplant recipients [27]. KSHV DNA detection in the tumor cells is a diagnostic criterion for PEL [28,29].
Multicentric Castleman’s disease is characterized by systemic inflammation, increased levels of cytokines IL-6, IL-10 and vIL6 and by clinical symptoms like lymphadenopathy, fever, diarrhea and weight loss. The patients with MCD are more prone to non-Hodgkin lymphomas and organ failure [30,31,32,33].
KSHV replication in KSHV-HIV positive patients may induce a systemic inflammation characterized by high levels of IL-6 and IL-10 and high KSHV viral load in the blood. This non-malignant clinical manifestation is called KSHV associated inflammatory cytokine syndrome (KICS). As the clinical symptoms and laboratory abnormalities overlap with those seen in MCD, the diagnosis of KICS is predicated on the absence of the lymphadenopathy seen in MCD. KICS can also arise in KS or PEL patients and is often associated with a more severe disease course [6,34].
Current antiviral treatments against KSHV mainly rely on herpesviral DNA polymerase inhibitors. Although some of them efficiently inhibit KSHV replication in tissue culture, their efficacy against KSHV-associated disease is limited. Surgery, radiotherapy, and chemotherapy combined with antiviral agents and immunomodulatory molecules are used to obtain the best outcome for these patients. In trying to develop novel approaches to an effective pharmacological treatment of KSHV-associated diseases, either the combination of antiviral drugs directed against different viral targets, or the targeting of cellular proteins that are required for viral persistence, replication (‘dependency factors’) or the growth of tumor cells are being explored.
2. KSHV DNA Polymerase Inhibitors
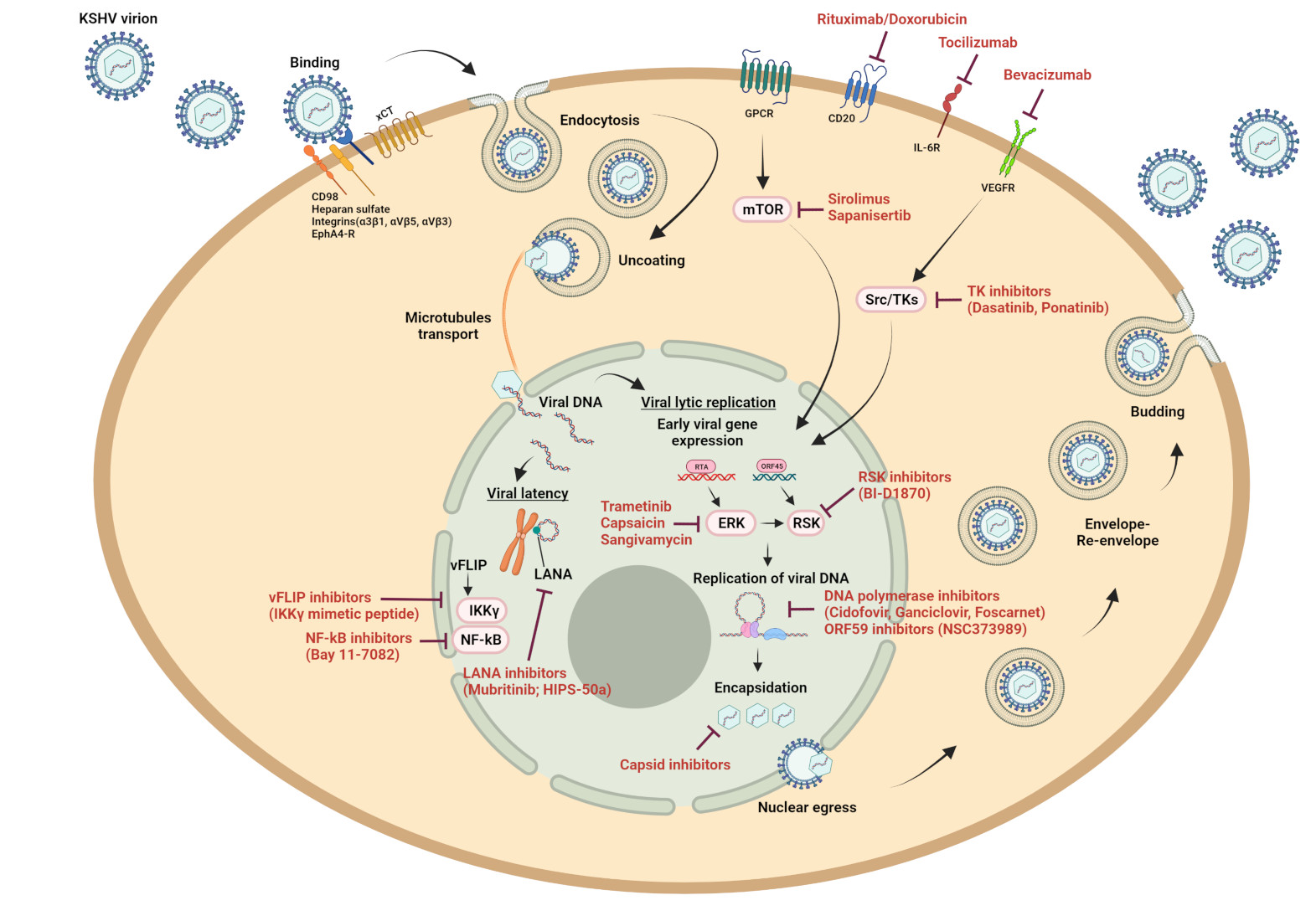
Since the discovery of KSHV, nucleoside inhibitors of the viral DNA polymerase such as ganciclovir, cidofovir, foscarnet, brivudine, and adefovir have provided the most potent inhibitors of KSHV replication in tissue culture [35,36,37,38,39,40]. In patients, some studies reported ganciclovir, valganciclovir, valacyclovir, famciclovir, cidofovir or foscarnet to reduce the shedding of KSHV in oral samples or KSHV viral load in peripheral blood, while others failed to notice pronounced effects of these drugs in treated patients [41,42,43]. With few exceptions [44], herpesviral DNA polymerase inhibitors (foscarnet, cidofovir, ganciclovir, valganciclovir) were found to be largely ineffective when used to treat established KS lesions [45,46,47,48]. In addition to these drugs, which are already approved for clinical use against other herpesviruses, several new promising nucleoside inhibitors have been identified in preclinical studies but are not yet approved or available for clinical treatments (for more details, see: [37,40,49,50,51].
The disappointing efficacy of herpesviral DNA polymerase inhibitors against KS is likely related to the fact that they are nucleoside analogs that, except for foscarnet, need to be phosphorylated to become active drugs. The first phosphorylation step is mediated by KSHV-encoded kinases, while generation of the nucleoside di- and triphosphates is due to the action of cellular kinases [40]. There are two KSHV kinases capable of activating nucleoside prodrugs by phosphorylation: a Ser/Thr kinase (vPK) encoded by open reading frame (ORF) 36 and a thymidine kinase (TK) encoded by ORF21. Both viral kinases are only expressed during the lytic phase of the viral cycle, and may thus only be able to exert their function in the relatively small number of infected cells in KS tumors that undergo lytic replication (see below).
Genes for herpesviral TKs occur in alpha- and gammaherpesviruses, but not in betaherpesviruses. In alpha- and gammaherpesviruses, the TK homologs are conserved: UL23 of herpes simplex virus 1 and 2 (HSV-1/2), ORF36 of varicella-zoster virus (VZV), BXLF1 of Epstein-Barr virus (EBV), and ORF21 of KSHV (52). Herpesviral TKs differ in their capacity to phosphorylate nucleoside analogs. Pyrimidine nucleosides such as brivudine and azidothymidine (zidovudine, the anti-HIV nucleoside reverse transcriptase inhibitor) are efficiently phosphorylated by KSHV TK [40,52]. Instead, PK/ORF36 efficiently phosphorylates purine analogs like valganciclovir [53].
As in the case of KS, the use of these viral DNA polymerase inhibitors against PEL and MCD has also in most cases only produced unsatisfactory clinical results [41,54,55,56,57,58]. The only notable exception is the combination of high dose ganciclovir and zidovudine in patients with MCD, which has shown promising clinical response rates [59]. This drug regimen is based on the activation of the prodrug zidovudine to a toxic moiety by KSHV TK, in combination with the antiviral effects of ganciclovir [59].
The limited efficacy of the DNA polymerase inhibitors pushed the scientific community to identify other suitable targets with different molecular mechanisms to treat KSHV-associated disease.
3. Antivirals Targeting Other Steps in the Viral Life Cycle
KSHV, like the other members of the Herpesviridae family, has a biphasic life cycle characterized by distinct patterns of viral gene expression [60]. KSHV establishes a permanent infection that lasts for the entire life of the infected host. During the KSHV latent phase, the viral DNA is maintained as a circular episome in the infected cells, replicated in dividing cells together with the cellular DNA [61,62], and a few latency-associated genes are transcribed: these encode the latency-associated nuclear antigen (LANA, encoded by ORF73), the viral homolog of cyclin D (vCyc-D/ORF72), the viral homolog of the fas-associated death domain-like interleukin-1-β-converting enzyme (FLICE-) inhibitory protein (vFLIP/ORF71), Kaposin (A, B and C encoded by K12) and 25 mature microRNAs [63,64,65,66]. In order to produce new viral progeny, KSHV has to periodically reactivate from latency and to switch into the productive (‘lytic’) phase of its life cycle. During the immediate-early and early stages of productive KSHV infection, only a subset of lytic viral genes is expressed. Following the replication of the viral DNA, viral genes encoding viral structural proteins are switched on (‘late’ phase of the productive replication cycle) in order to allow the production of new viral progeny [67].
In KS and PEL, the majority of KSHV infected cells adopt the latent program; therefore, a considerable effort has been made to target latent viral proteins or cellular pathways in which they interfere.
LANA (latency-associated nuclear antigen) is expressed in all the latently KSHV-infected cells [35,68]. In PEL cells, decreasing LANA expression with shRNA [69], by treatment with glycyrrhizic acid [70] or HSP90 inhibitors [71] induces cell death. The gene-editing technique, CRISPR-Cas9, has also been used successfully in two studies to target KSHV LANA and to target KSHV latency [72,73]. These findings suggest that LANA may be a promising viral target to disrupt KSHV latency. Its C-terminal DNA-binding domain (DBD) binds the latent KSHV replication origin in the terminal repeat (TRs) region of the viral genome; this interaction ensures the viral genome replication and segregation during cell mitosis [62,74,75,76,77]. The structure of the LANA DBD alone and in complex with the viral latent replication origin has been solved [76,78], allowing Kirsch and colleagues to discover and optimize new small compounds able to inhibit the binding of LANA to viral DNA in the low micromolar range [79,80,81]. In addition, Mubritinib (TAK165) was identified as a potent inhibitor of LANA-DNA binding and strongly reduced living KSHV PEL cells in vitro and in vivo [82].
Another potential target of the latent KSHV cycle is the viral FLICE-inhibitory protein (vFLIP). vFLIP is a potent activator of the NF-kB pathway and counteracts Fas-induced apoptosis [83,84]. Silencing vFLIP using siRNA [85] or using NF-kB inhibitors such as Bay 11-7082 [83,86,87,88] induces PEL cell apoptosis, suggesting that vFLIP may also represent an attractive therapeutic target. In order to activate the NF-kB pathway, vFLIP directly interacts with IKKγ/NEMO, a key player in the canonical NF-kB pathway [85,89]. The structure of a fragment of the coiled-coil domains of IKKγ/NEMO in complex with vFLIP has been solved [90], which provided the basis for a structure-guided development of vFLIP inhibitors. A conformationally constrained, stapled IKKγ peptide derived from the IKKγ–vFLIP interaction site interferes with the binding of IKKγ to vFLIP and enhances apoptosis in PEL cell lines [91]. Also, a tertiary protein structure mimic of the vFLIP-interaction site in the IKKγ/NEMO helix was able to induce cell death in PEL cell lines and to delay tumor growth in a PEL xenograft mouse model [88]. These findings indicate that it may be feasible to develop small molecule inhibitors targeting the vFLIP-IKKγ/NEMO interaction and showing a therapeutic effect against some KSHV-associated diseases.
New approaches to target the productive (‘lytic’) phase of the KSHV life cycle have also been developed but remain at a preclinical stage. Among the viral immediate early proteins to be expressed early after lytic reactivation are RTA, K-bZIP, and pORF45, crucial regulatory proteins and/or transcription factors [92,93]. RTA (encoded by ORF50) is necessary and sufficient to trigger the KSHV lytic phase, thus it is called “the master of KSHV lytic-switch” [94]. Long and colleagues recently described the efficiency of Gallic acid (GA) to inhibit RTA transcriptional activity by preventing its binding to target promoters. GA induces apoptosis in a PEL cell line in a dose-dependent manner [95].
Another novel viral target is the KSHV protein encoded by ORF59. pORF59 is a homologue of the DNA polymerase-associated processivity factor, which occurs in pro- as well as eukaryotes and in all herpesviruses. By acting in concert with the KSHV DNA polymerase encoded by ORF9, it facilitates the elongation of newly synthesized viral DNA. The compound NSC373989 was shown to target the pORF9/pORF59 complex and to inhibit viral DNA synthesis in vitro as well as KSHV lytic reactivation in PEL cells [96]. An inhibitor of pORF59 could provide an alternative approach to inhibiting viral DNA replication during the lytic phase of the replication and could potentially be used in combination with established competitive DNA polymerase inhibitors.
The structural similarity between the RNAse H-like nucleotidyltransferase domain contained in the HIV integrase and the two single-strand DNA (ssDNA) binding proteins essential for herpesviral DNA replication allowed the identification of XZ45, an HIV integrase inhibitor, as a compound that also inhibits the replication of KSHV and other herpesviruses [97]. Raltegravir and Dolutegravir, two HIV integrase inhibitors approved for clinical use against HIV, were successfully tested in vitro for their inhibition of the KSHV large terminase subunit encoded by pORF29. The C-terminal domain of KSHV pORF29 also shows a high degree of similarity with RNAse H-like nucleotidyltransferases and its inhibition impairs KSHV lytic reactivation in tissue culture [98].
In the case of herpes simplex virus (HSV), Varicella-Zoster virus (VZV), and human cytomegalovirus (HCMV), structure-based drug design has been employed to target capsid proteins, capsid assembly, and DNA encapsidation and this approach has already shown promising results [99,100,101,102]. One compound, letermovir, targets the HCMV terminase and the incorporation of viral DNA into newly formed viral capsids and has been approved for clinical use in stem cell transplant recipients [103]. However, it is not active against KSHV or other human herpesviruses. For KSHV, inhibitors of the pORF17 scaffold/protease polyprotein, which transiently fills the newly assembled capsid and is then released by autoproteolytic cleavage to allow packaging of capsids with viral DNA [104], have been developed and some have shown potency in tissue culture [105,106,107]. Nelfinavir, an HIV protease inhibitor, has been shown by Gantt and colleagues to inhibit KSHV, HSV and HCMV replication. Whether the KSHV pORF17 scaffold/protease is the key target of nelfinavir however still remains unclear [108,109].
Attempts were also made to target viral glycoproteins expressed on the surface of KSHV infected cells such as K8.1 and gH, by using immunotoxins that bind to these viral proteins. These immunotoxins could induce cell death in KSHV-infected cells in tissue culture, and a combination with ganciclovir increased their effect [110,111].
An opposite approach to inhibiting the viral lytic replication cycle involves its activation, with the aiming of taking advantage of the cell death occurring in lytically reactivated cells. Different treatments aimed to induce the viral lytic cycle in latently KSHV-infected cells have been attempted. Treatment with histone deacetylase (HADAC) inhibitors like sodium butyrate (NaB/SB), 12-O-tetradecanoylphorbol-13-acetate (TPA), and trichostatin (TSA), with the DNA methyltransferase inhibitor 5-Azacytidine (5-AZaC) or with some African autochthonous plant extracts can induce the lytic reactivation in vitro [112,113,114,115]. The proteasome inhibitor bortezomib induces KSHV and EBV lytic reactivation and it was successfully used in a clinical trial in combination with ganciclovir to treat MCD [57]. Liang and colleagues used the CRISPR-Cas9 system to inhibit the expression of KSHV miRNAs in latently KSHV positive PEL cells. This alters the expression of the mature miRNAs and induces upregulation of the viral lytic genes [116]. Recently, the suberoyl bis-hydroxamic acid (SAHA) was selected for its ability to induce KSHV lytic reactivation and apoptosis in a dose-dependent manner in PEL cells, indicating its possible therapeutic use [117].
Another possible strategy to counteract KSHV infection could be inhibition of virus entry into the target cell. This has so far proved difficult, because KSHV entry is mediated through diverse receptors, depending on the cell type that is to be infected. Binding of KSHV to the cell surface is achieved by heparan sulfate-proteoglycans (HSPGs) and DC-SIGN and entry is mediated through Ephrin receptors (EPHA2, 4, 5 and 7), integrins (α3β1, αVβ3, αVβ5, and α5β1) and xCT (reviewed in [118]).
4. Cellular Targets to Inhibit KSHV Replication
Instead of targeting the virus replication directly by inhibiting a viral protein, there are several ways of inhibiting the virus by targeting cellular processes that are essential for the virus to survive. As viruses hijack the host cells and exploit the cell machinery, there are several cellular proteins that play an important role in the viral life cycle and that can be targeted pharmacologically. A potential drawback of this approach is that these cellular processes are also important for cellular functions and that the inhibition of these targets may be accompanied by side effects.
4.1. Kinase Inhibitors
In KSHV infected cells, cellular tyrosine kinases play an essential role during the KSHV life cycle. Therefore, targeting cellular receptor tyrosine kinases such as c-kit, PDGFR, VEGFR, and Eph2A for antiviral treatment was investigated in several clinical studies. Treatment of KS patients with imatinib, a c-abl tyrosine kinase inhibitor in clinical use for the treatment of chronic myelogenous leukemia, resulted in clinical and histological regression of KS lesions in some patients [119,120]. Furthermore, sorafenib, which inhibits the VEGF receptor and is used to treat several malignancies like hepatocellular carcinoma or kidney carcinoma, was shown in a case report to achieve a complete remission of KS lesions in one patient [121]. Unfortunately, sorafenib showed only moderate effects when used to treat KS in a phase 1b clinical trial [122].
Of the more than 20 currently available inhibitors of cellular tyrosine kinases, five compounds (dasatinib, ponatinib, bosutinib, gefitinib, and nilotinib) were shown to also inhibit KSHV thymidine kinase (TK/pORF21), which, in contrast to its name, acts as an efficient protein tyrosine kinase [52,123]. Dasatinib and ponatinib also strongly inhibited KSHV early viral gene expression and the production of new viral progeny in B, endothelial and epithelial cells, most likely as a result of the inhibition of cellular tyrosine kinases, and dasatinib inhibited the growth of KSHV-driven endothelial tumors in a mouse xenograft model [52]. UNC3810A, a small molecule inhibitor of the receptor tyrosine kinase Tyro3, a member of the Tyro3/Axl/Mer (TAM) family of tyrosine kinases that promote the proliferation and survival of several cancers, was shown to be a potent inhibitor of PEL cell growth in a mouse xenograft model [124].
In KSHV infected cells, mTOR, a cellular kinase that belongs to the phosphatidylinositol 3-kinase related family of protein kinases, plays a key role in the life cycle of KSHV by promoting the activation of mTOR downstream of the viral G-protein coupled receptor (GPCR) homologue vGPCR. Blocking mTOR activity by rapamycin (sirolimus) inhibited cell growth in cell culture and tumor growth in a KSHV related mouse model, whereas overstimulation of the mTOR pathway resulted in the opposite, showing that this pathway is important for KSHV [125,126]. Several mTOR inhibitors reduce the growth of PEL cell lines in tissue culture and/or in mouse xenograft models, in particular when combined with an AKT inhibitor [127,128,129]. An ATP-competitive inhibitor of mTOR, MLN0128 (sapanisertib), induces apoptosis in PEL cells and reduces the growth of PEL in a xenograft model at nanomolar IC50 concentrations and is still effective against doxorubicin- or rapamycin-resistant PEL cell clones [129].
Similarly, the ERK/MAPK pathway was shown to be crucial in KSHV induced pathogenesis [130]. Several experimental compounds, e.g., sangivamycin and capsaicin, as well as trametinib, a MEK1/2 inhibitor approved for clinical use, can inhibit ERK activation and KSHV reactivation, and/or are able to induce apoptosis in PEL cell lines, suggesting that they could perhaps be used for the treatment of PEL [52,131,132]. BI-D1870, a RSK1/2 inhibitor, suppresses KSHV lytic gene expression and virus production. The RSK1/2 serine/threonine kinase is activated by its interaction with KSHV pORF45 and a small peptide blocking this interaction has been shown to inhibit viral lytic gene expression and viral progeny formation [133]. Furthermore, crizotinib, an inhibitor of ALK and c-Met, suppresses the growth of PEL cells in a mouse xenograft model [134].
4.2. HSP90 and HSP70 Inhibitors
HSP90 is a molecular chaperone required for the correct folding of cellular proteins. HSP90 inhibitors have found use as anti-cancer drugs to treat several malignancies, including lung or prostate cancer. Several groups have shown that HSP90 is also involved in essential steps in the KSHV life cycle: vFLIP, a viral latent protein, is found in a complex containing IKKγ/NEMO and HSP90 [135]. HSP90 serves as a co-factor for MAPK activation and latent viral gene expression of KSHV, and the KSHV K1 protein was shown to bind HSP90ᵦ [89,136,137].
Several HSP90 inhibitors have been used in cell culture studies and mouse models of KSHV malignancies. In particular treating KSHV positive cells with different HSP90 inhibitors (PU-H71, AUY922, BIIB021, NVP-BEP800, or 17-DMAG) leads to the proteasomal degradation of LANA and Eph2A and inhibited cell growth as well as induced apoptosis in PEL cells [71,138]. Furthermore, PU-H71, BIIB021, and AUY922 also repressed tumor progression in xenograft mouse models [138,139,140]. These are encouraging data, but HSP90 inhibitors have not yet been tested in clinical trials against KSHV malignancies.
Several HSP70 chaperone family members are involved in the formation of KSHV nuclear replication and transcription compartments (RTCs) during the early stages of the KSHV lytic cycle, and a small molecule HSP70 inhibitor, VER-155008, blocked KSHV RTC formation [141]. Formation of KSHV RTCs also involves neddylation, and the neddylation inhibitor MLN 4924 induces apoptosis in PEL cells [142].
4.3. Other Cellular Targets
Another promising drug candidate for treating patients with KSHV malignancies is bortezomib, a proteasome inhibitor, which has been shown to promote the KSHV and EBV lytic cycle. Bortezomib activates JNK and induces autophagy and apoptosis in PEL cell lines [143,144] and in a xenograft mouse model [145]. Further clinical studies confirmed this drug as a potential treatment against AIDS-associated KS in a pilot trial study, AMC-063 [146] and in combination with ganciclovir against MCD in a case report [55].
Recently, Chen et al. could show that pemetrexed, an anti-cancer drug already in clinical use, is able to inhibit the lytic replication of KSHV by blocking the dTMP synthesis in infected cells [147].
Other cellular factors that are important for the KSHV replication machinery and which could be targeted pharmacologically include topoisomerase II: (+)-Rutamarin, a topoisomerase II inhibitor that efficiently inhibits KSHV lytic DNA replication in BCBL-1 cells [148].
Hypoxia and hypoxia-mediated signaling are important factors that drive KSHV replication and may play a role in KSHV-associated malignancies. Thus, hypoxia-inducible factors (HIFs), the first mediators of the cellular response to hypoxia, play a crucial role in KSHV induced tumors and have been shown to activate KSHV lytic replication [149]. HIF1α can directly activate RTA, LANA, as well as the ORF34-37 cluster of lytic genes [150,151]. A small molecule inhibitor of HIF1α, PX-478, was shown to achieve a significant inhibition of PEL cell growth in tissue culture, suggesting that HIF1α could be a suitable target for treating this disease [152].
Other cellular metabolic targets include the metabolic sensor SIRT1 that is functionally required for sustaining the proliferation and survival of PEL cells. Inhibition of SIRT with the inhibitor tenovin-6 induced cell cycle arrest and apoptosis in PEL cell culture and significantly extended the survival of mice in a murine PEL model [153].
Heme oxygenase 1 (HO-1) is highly expressed in KSHV-infected HUVECs cells. Targeting HO-1 by siRNA or by the chemical inhibitor SnPP induces cell death in KSHV-infected endothelial cells and inhibits their growth as tumors in nude mice [154].
However, with the exceptions mentioned above, most of these inhibitors directed at cellular targets have so far not been studied in clinical trials. Nevertheless, these findings show the potential of targeting cellular mechanisms for the treatment of KSHV-associated diseases.
5. Monoclonal Antibodies and Immunomodulatory Therapies
Several monoclonal antibodies have been used to treat KSHV-related diseases. One of the targets to have been explored in this manner is the vascular endothelial growth factor (VEGF). Bevacizumab, a humanized monoclonal antibody against VEGF-A, showed an overall response rate of 31% in patients with HIV-associated KS [155,156]. Furthermore, rituximab, a monoclonal antibody against CD20 that is widely used to treat several types of B-cell lymphoma and autoimmune diseases, is effective in clinical trials against multicentric Castleman’s disease, either alone or in combination with liposomal doxorubicin [157]. However, treatment with rituximab can also cause the progression of KS in these patients [158,159].
Tocilizumab, a monoclonal antibody directed against the inteleukin-6 receptor that is currently used to treat rheumatoid arthritis and other autoimmune diseases, has also been shown to be beneficial in treating MCD [160,161,162].
The combination of lenalidomide, an immunomodulatory drug, with arsenic trioxide (ATO), which is normally used to treat acute promyelocytic leukemia, has been reported to produce encouraging results when treating PEL in a xenograft mouse model. In this study, lenalidomide/ATO treatment decreased the proliferation of PEL cells and downregulated the expression of KSHV latent viral proteins. This was associated with less NF-kB expression and downregulation of IL-6 and IL-10 as well as the inhibition of VEGF and the induction of apoptosis [163].
Pomalidomide, another immunomodulatory drug, has been shown to act against KS in HIV-negative and HIV-positive people in a clinical Phase I/II trial. Pomalidomide induced an increase in ICAM-1 and B7-2 expression in PEL cells, thereby leading to T cell activation and NK cell-mediated killing of PEL cells, which makes pomalidomide a promising candidate for the treatment of KSHV related malignancies [164,165].
6. KSHV Tropism and Models to Study the Virus
KSHV shows a relatively strict species tropism for humans. In vivo, viral DNA has been detected in human B cells [166], macrophages [167], keratinocytes, endothelial cells [168,169] and epithelial cells [170]. In vitro, KSHV can infect a broad spectrum of different cells including epithelial cells, endothelial cells, keratinocytes, fibroblasts, B- and T-lymphocytes, monocytes, macrophages and dendritic cells [167,168,171,172]. Besides, KSHV can infect non-human cell lines such as owl monkey kidney cells, baby hamster kidney fibroblasts cells, Chinese hamster ovary cells and mouse fibroblasts [173].
The B-cell lines derived from PEL patients are used to study KSHV pathogenesis in vitro as they are able to maintain the viral genome in a latent state. In contrast, the endothelial spindle cells lose the virus after a few passages in cell culture [174].
KSHV-infected lymphatic endothelial cells (LECs) exploit a unique transcription program with the expression of latent and lytic genes, which differs from the latency program described in stably infected blood endothelial cells (HUVECs) [175,176], both models are used to characterize KSHV molecular mechanisms of action.
Because of its restricted species tropism, studying KSHV infection in vivo is not straightforward. There are mainly three ways of how KSHV infection can be studied in vivo (nicely reviewed in [177]): the first is to infect non-human primates like common marmosets with KSHV [178]. The second approach involves the use of KSHV related viruses, like murine herpesvirus 68 (MHV-68), rhesus rhadinovirus (RRV) [179] or herpesvirus saimiri (HVS) [180].
MHV-68 infects mice and is used as a model to mimic KSHV infection, because it has been shown that 90% of the MHV-68 genes have homologs to KSHV [181]. MHV-68 behaves like a natural persisting pathogen in mice but without showing the disease. Therefore, MHV-68 is often used to study immunological topics like immunevasion and infection control by T cells. Nevertheless, this model is not suitable to study host colonialization and viral reactivation nor can it be used to study KSHV related malignancies [182].
The third approach, which is becoming increasingly more important, is to use humanized mice. In a xenograft mouse model, human PEL derived cell lines are implanted into immunodeficient mice to establish a PEL like phenotype [182,183]. McHugh et al. described an animal model, showing that coinfection with EBV establishes persistent KSHV infection in B cells, resulting in a PEL like phenotype in these mice [184].
7. Conclusions
Immunocompromised patients and people living in KSHV-endemic areas are most likely to be affected by KSHV-induced diseases. Despite more than 25 years of research on KSHV, we still lack effective therapies to counteract KSHV infection, reactivation and pathogenicity (Table S1). However, progress has been made over recent years and both new viral as well as cellular ‘druggable’ targets have emerged. Some of the insights into KSHV pathogenesis that have been made over the last two decades have also laid the ground for the development of active compounds that may find use in other malignancies. As KSHV-related malignancies are not among the most common cancers and often occur in economically disadvantaged countries, the development of effective drugs against KSHV and KSHV-associated diseases faces the obstacle of a lack of interest on the part of the pharmaceutical industry. Focusing on mechanisms of pathogenicity that are shared between KSHV-related and other, more common, malignancies may open up a way forward to overcome this obstacle.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/v13091797/s1, Table S1: Therapies in clinical trials.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Acknowledgments
Work in TFS’ group is supported by by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under Germany’s Excellence Strategy—EXC 2155 “RESIST”—Project ID 39087428 and the Collaborative Research Center 900 ‘Chronic Infections’ (Project C1), as well as the German Center for Infection Research (TTU IICH) and the Lower Saxony Ministry of Science and Culture (project 14-76103-184).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chang, Y.; Cesarman, E.; Pessin, M.; Lee, F.; Culpepper, J.; Knowles, D.; Moore, P. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soulier, J.; Grollet, L.; Oksenhendler, E.; Cacoub, P.; Cazals-Hatem, D.; Babinet, P.; D’Agay, M.F.; Clauvel, J.P.; Raphael, M.; Degos, L. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman’s disease. Blood 1995, 86, 1276–1280. [Google Scholar] [CrossRef] [Green Version]
- Cesarman, E.; Chang, Y.; Moore, P.; Said, J.W.; Knowles, D.M. Kaposi’s Sarcoma–Associated Herpesvirus-Like DNA Sequences in AIDS-Related Body-Cavity–Based Lymphomas. N. Engl. J. Med. 1995, 332, 1186–1191. [Google Scholar] [CrossRef]
- Bouvard, V.; Baan, R.; Straif, K.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens—Part B: Biological agents. Lancet Oncol. 2009, 10, 321–322. [Google Scholar] [CrossRef]
- Longnecker, R.; Neipel, F. Introduction to the human γ-herpesviruses. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Eds.; Cambridge Ubiversity Press: Cambridge, UK, 2007. [Google Scholar]
- Uldrick, T.S.; Wang, V.; O’Mahony, D.; Aleman, K.; Wyvill, K.M.; Marshall, V.; Steinberg, S.M.; Pittaluga, S.; Maric, I.; Whitby, D.; et al. An Interleukin-6–Related Systemic Inflammatory Syndrome in Patients Co-Infected with Kaposi Sarcoma–Associated Herpesvirus and HIV but without Multicentric Castleman Disease. Clin. Infect. Dis. 2010, 51, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, A.Y.; Strauchen, J.A.; Lee, G.; Scigliano, E.; Hale, E.E.; Weisse, M.T.; Burstein, D.; Kamel, O.; Moore, P.S.; Chang, Y. Posttransplantation Plasmacytic Proliferations Related to Kaposi’s Sarcoma–Associated Herpesvirus. Am. J. Surg. Pathol. 1999, 23, 1393. [Google Scholar] [CrossRef] [PubMed]
- Kapelushnik, J.; Ariad, S.; Benharroch, D.; Landau, D.; Moser, A.; Delsol, G.; Brousset, P. Post renal transplantation human herpesvirus 8-associated lymphoproliferative disorder and Kaposi’s sarcoma. Br. J. Haematol. 2001, 113, 425–428. [Google Scholar] [CrossRef]
- Dupin, N.; Diss, T.L.; Kellam, P.; Tulliez, M.; Du, M.-Q.; Sicard, D.; Weiss, R.A.; Isaacson, P.G.; Boshoff, C. HHV-8 is associated with a plasmablastic variant of Castleman disease that is linked to HHV-8–positive plasmablastic lymphoma. Blood 2000, 95, 1406–1412. [Google Scholar] [CrossRef] [PubMed]
- Orenstein, J.M. Ultrastructure of Kaposi Sarcoma. Ultrastruct. Pathol. 2008, 32, 211–220. [Google Scholar] [CrossRef]
- Antman, K.; Chang, Y. Kaposi’s sarcoma. N. Engl. J. Med. 2000, 342, 1027–1038. [Google Scholar] [CrossRef]
- Kaposi, M. Idiopathisches multiples Pigmentsarkom der Haut. Arch. Dermatol. Syph. 1872, 4, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Plancoulaine, S.; Abel, L.; van Beveren, M.; Trégouët, D.A.; Joubert, M.; Tortevoye, P. Human herpesvirus 8 transmission from mother to child and between siblings in an endemic population. Lancet 2000, 356, 1062–1065. [Google Scholar] [CrossRef]
- Dedicoat, M.; Newton, R.; Alkharsah, K.R.; Sheldon, J.; Szabados, I.; Ndlovu, B. Mother-to-Child Transmission of Human Herpesvirus-8 in South Africa. J. Infect. Dis. 2004, 190, 1068–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebbé, C.; Legendre, C.; Francès, C. Kaposi sarcoma in transplantation. Transplant. Rev. 2008, 22, 252–261. [Google Scholar] [CrossRef]
- Cattani, P.; Capuano, M.; Graffeo, R.; Ricci, R.; Cerimele, F.; Cerimele, D.; Nanni, G.; Fadda, G. Kaposi’s Sarcoma Associated with Previous Human Herpesvirus 8 Infection in Kidney Transplant Recipients. J. Clin. Microbiol. 2001, 39, 506–508. [Google Scholar] [CrossRef] [Green Version]
- Grulich, E.A.; van Leeuwen, M.; Falster, M.; Vajdic, C. Incidence of cancers in people with HIV/AIDS compared with immunosuppressed transplant recipients: A meta-analysis. Lancet 2007, 370, 59–67. [Google Scholar] [CrossRef]
- Francès, C.; Marcelin, A.G.; Legendre, C.; Chevret, S.; Dussaix, E.; Lejeune, J.T. The impact of Preexisting or Acquired Kaposi Sarcoma Herpesvirus infection in Kidney Trasplant Recipiens on Morbidity and Survival. Am. J. Transpl. 2009, 9, 2580–2586. [Google Scholar] [CrossRef]
- Wabinga, H.R.; Nambooze, S.; Amulen, P.M.; Okello, C.; Mbus, L.; Parkin, D.M. Trends in the incidence of cancer in Kampala, Uganda 1991–2010. Int. J. Cancer. 2014, 135, 432–439. [Google Scholar] [CrossRef]
- Friedman-Kien, A.E. Disseminated Kaposi’s sarcoma syndrome in young homosexual men. J. Am. Acad. Dermatol. 1981, 5, 468–471. [Google Scholar] [CrossRef]
- Dupin, N.; De Cervens, V.R.; Gorin, I.; Calvez, V.; Pessis, E.; Grandadam, M.; Rabian, C.; Viard, J.P.; Huraux, J.M.; Escande, J.P. The influence of highly active antiretroviral therapy on AIDS-associated Kaposi’s sarcoma. Br. J. Dermatol. 1999, 140, 875–881. [Google Scholar] [CrossRef]
- Cattelan, A.; Calabro’, M.; Aversa, S.; Zanchetta, M.; Meneghetti, F.; De Rossi, A.; Chieco-Bianchi, L. Regression of AIDS-related Kaposi’s sarcoma following antiretroviral therapy with protease inhibitors: Biological correlates of clinical outcome. Eur. J. Cancer 1999, 35, 1809–1815. [Google Scholar] [CrossRef]
- Van Leeuwen, M.; Vajdic, C.; Middleton, M.G.; McDonald, A.M.; Law, M.; Kaldor, J.M.; Grulich, A.E. Continuing declines in some but not all HIV-associated cancers in Australia after widespread use of antiretroviral therapy. AIDS 2009, 23, 2183–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grulich, A.E.; Li, Y.; McDonald, A.M.; Correll, P.K.; Law, M.G.; Kaldor, J.M. Decreasing rates of Kaposi’s sarcoma and non-Hodgkin’s lymphoma in the era of potent combination anti-retroviral therapy. AIDS 2001, 15, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Knowles, D.M.; Inghirami, G.; Ubriaco, A.; Dalla-Favera, R. Molecular Genetic Analysis of Three AIDS-Associated Neoplasms of Uncertain Lineage Demonstrates Their B-Cell Derivation and the Possible Pathogenetic Role of the Epstein-Barr Virus. Blood 1989, 73, 792–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Z.G.; Zhang, Q.Y.; Lu, Z.B.; Quinto, T.; Rozenvald, I.B.; Liu, L.-T. Extracavitary KSHV-associated large B-Cell lymphoma: A distinct entity or a subtype of primary effusion lymphoma? Study of 9 cases and review of an additional 43 cases. Am. J. Surg. Pathol. 2012, 36, 1129–1140. [Google Scholar] [CrossRef]
- Boulanger, E.; Afonso, P.; Yahiaoui, Y.; Adle-Biassette, H.; Gabarre, J.; Agbalika, F. Human Herpesvirus-8 (HHV-8)-Associated Primary Effusion Lymphoma in two Renal Transplant Recipients Receiving Rapamycin. Arab. Archaeol. Epigr. 2008, 8, 707–710. [Google Scholar] [CrossRef]
- Nador, R.G.; Cesarman, E.; Chadburn, A.; Dawson, D.B.; Ansari, M.Q.; Sald, J.; Knowles, D.M. Primary effusion lymphoma: A distinct clinicopathologic entity associated with the Kaposi’s sarcoma-associated herpes virus. Blood 1996, 88, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Song, J.Y.; Jaffe, E.S. HHV-8–positive but EBV-negative primary effusion lymphoma. Blood 2013, 122, 3712. [Google Scholar] [CrossRef] [Green Version]
- Castleman, B.; Towne, V.W. Case Records of the Massachusetts General Hospital: Case No. 40231. N. Engl. J. Med. 1954, 250, 1001–1005. [Google Scholar] [CrossRef]
- Du, M.-Q.; Liu, H.; Diss, T.C.; Ye, H.; Hamoudi, R.A.; Dupin, N. Kaposi sarcoma-associated herpesvirus infects monotypic (IgM lambda) but polyclonal naive B cells in Castleman disease and associated lymphoproliferative disorders. Blood 2001, 97, 2130–2136. [Google Scholar] [CrossRef]
- Aoki, Y.; Tosato, G.; Fonville, T.W.; Pittaluga, S. Serum viral interleukin-6 in AIDS-related multicentric Castleman disease. Blood 2001, 97, 2526–2527. [Google Scholar] [CrossRef] [Green Version]
- Oksenhendler, E.; Boulanger, E.; Galicier, L.; Du, M.-Q.; Dupin, N.; Diss, T.C.; Hamoudi, R.; Daniel, M.-T.; Agbalika, F.; Boshoff, C.; et al. High incidence of Kaposi sarcoma–associated herpesvirus–related non-Hodgkin lymphoma in patients with HIV infection and multicentric Castleman disease. Blood 2002, 99, 2331–2336. [Google Scholar] [CrossRef]
- Polizzotto, M.N.; Uldrick, T.S.; Hu, D.; Yarchoan, R. Clinical Manifestations of Kaposi Sarcoma Herpesvirus Lytic Activation: Multicentric Castleman Disease (KSHV–MCD) and the KSHV Inflammatory Cytokine Syndrome. Front. Microbiol. 2012, 3, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kedes, D.H.; Ganem, D. Sensitivity of Kaposi’s sarcoma-associated herpesvirus replication to antiviral drugs. Implications for potential therapy. J. Clin. Investig. 1997, 99, 2082–2086. [Google Scholar] [CrossRef] [PubMed]
- Medveczky, M.M.; Horvath, E.; Lund, T.; Medveczky, P.G. In vitro antiviral drug sensitivity of the Kaposi’s sarcoma-associated herpesvirus. AIDS 1997, 11, 1327–1332. [Google Scholar] [CrossRef]
- Neyts, J.; De Clercq, E. Antiviral drug susceptibility of human herpesvirus 8. Antimicrob. Agents Chemother. 1997, 41, 2754–2756. [Google Scholar] [CrossRef] [Green Version]
- Sergerie, Y.; Boivin, G. Evaluation of Susceptibility of Human Herpesvirus 8 to Antiviral Drugs by Quantitative Real-Time PCR. J. Clin. Microbiol. 2003, 41, 3897–3900. [Google Scholar] [CrossRef] [Green Version]
- Coen, N.; Singh, U.; Vuyyuru, V.; Oord, J.J.V.D.; Balzarini, J.; Duraffour, S.; Snoeck, R.; Cheng, Y.C.; Chu, C.K.; Andrei, G. Activity and Mechanism of Action of HDVD, a Novel Pyrimidine Nucleoside Derivative with High Levels of Selectivity and Potency against Gammaherpesviruses. J. Virol. 2013, 87, 3839–3851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coen, N.; Duraffour, S.; Topalis, D.; Snoeck, R.; Andrei, G. Spectrum of Activity and Mechanisms of Resistance of Various Nucleoside Derivatives against Gammaherpesviruses. Antimicrob. Agents Chemother. 2014, 58, 7312–7323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luppi, M.; Trovato, R.; Barozzi, P.; Vallisa, D.; Rossi, G.; Re, A.; Ravazzini, L.; Potenza, L.; Riva, G.; Morselli, M.; et al. Treatment of herpesvirus associated primary effusion lymphoma with intracavity cidofovir. Leukemia 2005, 19, 473–476. [Google Scholar] [CrossRef]
- Casper, C.; Krantz, E.M.; Corey, L.; Kuntz, S.R.; Wang, J.; Selke, S.; Hamilton, S.; Huang, M.L.; Wald, A. Valganciclovir for suppression of human herpesvirus-8 replication: A randomized, Double-blind, Placebo-Controlled, Crossover trial. J. Infect. Dis. 2008, 198, 23–30. [Google Scholar] [CrossRef]
- Cattamanchi, A.; Saracino, M.; Selke, S.; Huang, M.-L.; Magaret, A.; Celum, C.; Corey, L.; Wald, A.; Casper, C. Treatment with valacyclovir, famciclovir, or antiretrovirals reduces human herpesvirus-8 replication in HIV-1 seropositive men. J. Med. Virol. 2011, 83, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzi, R.; Parisi, S.G.; Sarmati, L.; Uccella, I.; Nicastri, E.; Carolo, G.; Gatti, F.; Concia, E.; Andreoni, M. Efficacy of cidofovir on human herpesvirus 8 viraemia and Kaposi’s sarcoma progression in two patients with AIDS. AIDS 2001, 15, 2061–2062. [Google Scholar] [CrossRef]
- Simonart, T.; Noel, J.C.; De Dobbeleer, G.; Parent, D.; Van Vooren, J.P.; De Clercq, E. Treatment of Classical Kaposi’s Sarcoma With Intralesional Injections of Cidofovir: Report of a Case. 1998. J. Med. Virol. 1998, 55, 215–218. [Google Scholar] [CrossRef]
- Little, R.F.; Merced-Galindez, F.; Staskus, K.; Whitby, D.; Aoki, Y.; Humphrey, R.; Pluda, J.M.; Marshall, V.; Walters, M.; Welles, L.; et al. A Pilot Study of Cidofovir in Patients with Kaposi Sarcoma. J. Infect. Dis. 2003, 187, 149–153. [Google Scholar] [CrossRef]
- Krown, S.E.; Dittmer, D.P.; Cesarman, E. Pilot Study of Oral Valganciclovir Therapy in Patients With Classic Kaposi Sarcoma. J. Infect. Dis. 2011, 203, 1082–1086. [Google Scholar] [CrossRef] [PubMed]
- Plachouri, K.; Oikonomou, C.; Sarantopoulos, A.; Koumoundourou, D.; Georgiou, S.; Spiliopoulos, T. Successful treatment and durable remission of human herpesvirus-8-induced Kaposi sarcoma and multicentric Castleman’s disease under valganciclovir in an HIV -negative patient. Dermatol. Ther. 2020, 33, e13419. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Burnette, A.; Dorjsuren, D.; Roberts, P.E.; Huleihel, M.; Shoemaker, R.H.; Marquez, V.E.; Agbaria, R.; Sei, S. Potent Antiviral Activity of North-Methanocarbathymidine against Kaposi’s Sarcoma-Associated Herpesvirus. Antimicrob. Agents Chemother. 2005, 49, 4980–4988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coen, N.; Duraffour, S.; Snoeck, R.; Andrei, G. KSHV Targeted Therapy: An Update on Inhibitors of Viral Lytic Replication. Viruses 2014, 6, 4731–4759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prichard, M.N.; Williams, J.D.; Komazin-Meredith, G.; Khan, A.R.; Price, N.B.; Jefferson, G.M.; Harden, E.A.; Hartline, C.B.; Peet, N.P.; Bowlin, T.L. Synthesis and Antiviral Activities of Methylenecyclopropane Analogs with 6-Alkoxy and 6-Alkylthio Substitutions That Exhibit Broad-Spectrum Antiviral Activity against Human Herpesviruses. Antimicrob. Agents Chemother. 2013, 57, 3518–3527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauclair, G.; Naimo, E.; Dubich, T.; Rückert, J.; Koch, S.; Dhingra, A.; Wirth, D.; Schulz, T.F. Targeting Kaposi’s Sarcoma-Associated Herpesvirus ORF21 Tyrosine Kinase and Viral Lytic Reactivation by Tyrosine Kinase Inhibitors Approved for Clinical Use. J. Virol. 2020, 94, e01791-19. [Google Scholar] [CrossRef]
- Gustafson, E.A.; Schinazi, R.F.; Fingeroth, J.D. Human Herpesvirus 8 Open Reading Frame 21 Is a Thymidine and Thymidylate Kinase of Narrow Substrate Specificity That Efficiently Phosphorylates Zidovudine but Not Ganciclovir. J. Virol. 2000, 74, 684–692. [Google Scholar] [CrossRef] [Green Version]
- Pastore, R.D.; Chadburn, A.; Kripas, C.; Schattner, E.J. Novel association of haemophagocytic syndrome with Kaposi’s sarcoma-associated herpesvirus-related primary effusion lymphoma. Br. J. Haematol. 2000, 111, 1112–1115. [Google Scholar]
- Sbenghe, M.M.; Besa, E.; Mahipal, A.; Florea, A.D.; Bray, P.; Caro, J. HHV-8–Associated Multicentric Castleman’s Disease in HIV-Negative Patient: A Novel Therapy for an Orphan Disease. Oncologist. 2012, 17, 145–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozbalak, M.; Tokatli, I.; Özdemirli, M.; Tecimer, T.; Ar, M.C.; Ornek, S.; Koroglu, A.; Laleli, Y.; Ferhanoglu, B. Is valganciclovir really effective in primary effusion lymphoma: Case report of an HIV(−) EBV(−) HHV8(+) patient. Eur. J. Haematol. 2013, 91, 467–469. [Google Scholar] [CrossRef]
- Kantarci, F.E.N.; Eren, R.; Gündoğan, C.; Huq, G.E.; Doğu, M.H.; Suyanı, E.; Gündoğdu, C. A HHV-8 positive, HIV negative multicentric Castleman disease treated with R-CEOP chemotherapy and valganciclovir combination. J. Infect. Chemother. 2016, 22, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Hawkes, E.; Chionh, F.; Chong, G.; Fracp, C.M.M.B. Durable remission of both multicentric Castleman’s disease and Kaposi’s sarcoma with valganciclovir, rituximab and liposomal doxorubicin in an HHV-8-positive, HIV-negative patient. J. Clin. Pharm. Ther. 2016, 42, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Uldrick, T.S.; Polizzotto, M.; Aleman, K.; O’Mahony, D.; Wyvill, K.M.; Wang, V.; Marshall, V.; Pittaluga, S.; Steinberg, S.M.; Tosato, G.; et al. High-dose zidovudine plus valganciclovir for Kaposi sarcoma herpesvirus-associated multicentric Castleman disease: A pilot study of virus-activated cytotoxic therapy. Blood 2011, 117, 6977–6986. [Google Scholar] [CrossRef] [Green Version]
- Dittmer, D.; Damania, B. Kaposi sarcoma associated herpesvirus pathogenesis (KSHV)—an update. Curr. Opin. Virol. 2013, 3, 238–244. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.; Wang, H.; Herndier, B.; Ganem, D. Restricted expression of Kaposi sarcoma-associated herpesvirus (human herpesvirus 8) genes in Kaposi sarcoma. Proc. Natl. Acad. Sci. USA 1996, 93, 6641–6646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballestas, M.E.; Chatis, P.A.; Kaye, K.M. Efficient Persistence of Extrachromosomal KSHV DNA Mediated by Latency-Associated Nuclear Antigen. Science 1999, 284, 641–644. [Google Scholar] [CrossRef]
- Dittmer, D.; Lagunoff, M.; Renne, R.; Staskus, K.; Haase, A.; Ganem, D. A Cluster of Latently Expressed Genes in Kaposi’s Sarcoma-Associated Herpesvirus. J. Virol. 1998, 72, 8309–8315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, K.; Kuppers, D.A.; Verma, S.C.; Sharma, N.; Murakami, M.; Robertson, E.S.; Cheung, C.Y.; Poon, L.L.M.; Ng, I.H.Y.; Luk, W.; et al. Induction of Kaposi’s Sarcoma-Associated Herpesvirus Latency-Associated Nuclear Antigen by the Lytic Transactivator RTA: A Novel Mechanism for Establishment of Latency. J. Virol. 2005, 79, 7819–7826. [Google Scholar] [CrossRef] [Green Version]
- Lan, K.; Kuppers, D.A.; Verma, S.C.; Robertson, E.S. Kaposi’s Sarcoma-Associated Herpesvirus-Encoded Latency-Associated Nuclear Antigen Inhibits Lytic Replication by Targeting Rta: A Potential Mechanism for Virus-Mediated Control of Latency. J. Virol. 2004, 78, 6585–6594. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Lu, S.; Zhang, Z.; Gonzalez, C.M.; Damania, B.; Cullen, B.R. Kaposi’s sarcoma-associated herpesvirus expresses an array of viral microRNAs in latently infected cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5570–5575. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Veettil, M.V.; Chandran, B. Kaposi’s Sarcoma Associated Herpesvirus Entry into Target Cells. Front. Microbiol. 2012, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupin, N.; Fisher, C.; Kellam, P.; Ariad, S.; Tulliez, M.; Franck, N. Distribution of human herpesvirus-8 latently infected cells in Kaposi’s sarcoma, multicentric Castleman’s disease, and primary effusion lymphoma. Proc. Natl. Acad. Sci. USA 1999, 96, 4546–4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, A.; Anderson, J.; Papanastasiou, A.; Takeuchi, Y.; Boshoff, C. Inhibiting primary effusion lymphoma by lentiviral vectors encoding short hairpin RNA. Blood 2005, 105, 2510–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curreli, F.; Friedman-Kien, A.E.; Flore, O. Glycyrrhizic acid alters Kaposi sarcoma–associated herpesvirus latency, triggering p53-mediated apoptosis in transformed B lymphocytes. J. Clin. Invest. 2005, 115, 642–652. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Sin, S.-H.; Wen, K.W.; Damania, B.; Dittmer, D.P. Hsp90 Inhibitors Are Efficacious against Kaposi Sarcoma by Enhancing the Degradation of the Essential Viral Gene LANA, of the Viral Co-Receptor EphA2 as well as Other Client Proteins. PLoS Pathog. 2012, 8, e1003048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tso, F.Y.; West, J.T.; Wood, C. Reduction of Kaposi’s Sarcoma-Associated Herpesvirus Latency Using CRISPR-Cas9 To Edit the Latency-Associated Nuclear Antigen Gene. J. Virol. 2019, 93, e02183-18. [Google Scholar] [CrossRef] [Green Version]
- Haddad, C.O.; Kalt, I.; Shovman, Y.; Xia, L.; Schlesinger, Y.; Sarid, R.; Parnas, O. Targeting the Kaposi’s sarcoma-associated herpesvirus genome with the CRISPR-Cas9 platform in latently infected cells. Virol. J. 2021, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ballestas, M.; Kaye, K.M. Kaposi’s Sarcoma-Associated Herpesvirus Latency-Associated Nuclear Antigen 1 Mediates Episome Persistence through cis -Acting Terminal Repeat (TR) Sequence and Specifically Binds TR DNA. J. Virol. 2001, 75, 3250–3258. [Google Scholar] [CrossRef] [Green Version]
- Barbera, J.A.; Ballestas, M.E.; Kaye, K.M. The Kaposi’s Sarcoma-Associated Herpesvirus Latency-Associated Nuclear Antigen 1 N Terminus Is Essential for Chromosome Association, DNA Replication, and Episome Persistence. J. Virol. 2004, 78, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Hellert, J.; Weidner-Glunde, M.; Krausze, J.; Lünsdorf, H.; Ritter, C.; Schulz, T.F.; Lührs, T. The 3D structure of Kaposi sarcoma herpesvirus LANA C-terminal domain bound to DNA. Proc. Natl. Acad. Sci. USA 2015, 112, 6694–6699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidner-Glunde, M.; Mariggiò, G.; Schulz, T.F. Kaposi’s Sarcoma-Associated Herpesvirus Latency-Associated Nuclear Antigen: Replicating and Shielding Viral DNA during Viral Persistence. J. Virol. 2017, 91, e01083-16. [Google Scholar] [CrossRef] [Green Version]
- Domsic, J.F.; Chen, H.-S.; Lu, F.; Marmorstein, R.; Lieberman, P.M. Molecular Basis for Oligomeric-DNA Binding and Episome Maintenance by KSHV LANA. PLoS Pathog. 2013, 9, e1003672. [Google Scholar] [CrossRef] [Green Version]
- Kirsch, P.E.; Jakob, V.; Oberhausen, K.; Stein, S.C.; Cucarro, I.S.; Schulz, T.F.; Empting, M. Fragment-Based Discovery of a Qualified Hit Targeting the Latency-Associated Nuclear Antigen of the Oncogenic Kaposi’s Sarcoma-Associated Herpesvirus/Human Herpesvirus 8. J. Med. Chem. 2019, 62, 3924–3939. [Google Scholar] [CrossRef]
- Kirsch, P.; Stein, S.C.; Berwanger, A.; Rinkes, J.; Jakob, V.; Schulz, T.F.; Empting, M. Hit-to-lead optimization of a latency-associated nuclear antigen inhibitor against Kaposi’s sarcoma-associated herpesvirus infections. Eur. J. Med. Chem. 2020, 202, 112525. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, P.; Jakob, V.; Elgaher, W.A.M.; Walt, C.; Oberhausen, K.; Schulz, T.; Empting, M. Discovery of Novel Latency-Associated Nuclear Antigen Inhibitors as Antiviral Agents Against Kaposi’s Sarcoma-Associated Herpesvirus. ACS Chem. Biol. 2020, 15, 388–395. [Google Scholar] [CrossRef]
- Calderon, A.; Soldan, S.S.; De Leo, A.; Deng, Z.; Frase, D.M.; Anderson, E.M.; Zhang, Y.; Vladimirova, O.; Lu, F.; Leung, J.C.; et al. Identification of Mubritinib (TAK 165) as an inhibitor of KSHV driven primary effusion lymphoma via disruption of mitochondrial OXPHOS metabolism. Oncotarget 2020, 11, 4224–4242. [Google Scholar] [CrossRef]
- Keller, S.A.; Schattner, E.J.; Cesarman, E. Inhibition of NF-B Induces Apoptosis of KSHV-Infected Primary Effusion Lymphoma Cells. 2000. Available online: https://ashpublications.org/blood/article-pdf/96/7/2537/1668374/h8190002537.pdf (accessed on 30 August 2021).
- Chugh, P.; Matta, H.; Schamus, S.; Zachariah, S.; Kumar, A.; Richardson, J.A.; Smith, A.L.; Chaudhary, P.M. Constitutive NF- B activation, normal Fas-induced apoptosis, and increased incidence of lymphoma in human herpes virus 8 K13 transgenic mice. Proc. Natl. Acad. Sci. USA 2005, 102, 12885–12890. [Google Scholar] [CrossRef] [Green Version]
- Guasparri, I.; Keller, S.A.; Cesarman, E. KSHV vFLIP Is Essential for the Survival of Infected Lymphoma Cells. J. Exp. Med. 2004, 199, 993–1003. [Google Scholar] [CrossRef]
- Keller, S.A.; Hernandez-Hopkins, D.; Vider, J.; Ponomarev, V.; Hyjek, E.; Schattner, E.J.; Cesarman, E. NF-κB is essential for the progression of KSHV- and EBV-infected lymphomas in vivo. Blood 2006, 107, 3295–3302. [Google Scholar] [CrossRef] [PubMed]
- Baloon, G.; Chen, K.; Perez, R.; Tam, W.; Cesarman, E. Kaposi Sarcoma Herpesvirus (KSHV) VFLIP Oncoprotein Induces B Cell Transdifferentiation and Tumorigenesis in Mice. J. Clin. Investig. 2011, 121, 1141–1153. [Google Scholar] [CrossRef] [Green Version]
- Sadek, J.; Wuo, M.G.; Rooklin, D.; Hauenstein, A.; Hong, S.H.; Gautam, A. Modulation of virus-induced NF-κB signaling by NEMO coiled coil mimics. Nat Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Field, N.; Low, W.; Daniels, M.; Howell, S.; Daviet, L.; Boshoff, C.; Collins, M. KSHV vFLIP binds to IKK-γ to activate IKK. J. Cell Sci. 2003, 116, 3721–3728. [Google Scholar] [CrossRef] [Green Version]
- Bagnéris, C.; Ageichik, A.V.; Cronin, N.; Wallace, B.; Collins, M.; Boshoff, C.; Waksman, G.; Barrett, T. Crystal Structure of a VFlip-IKKγ Complex: Insights into Viral Activation of the IKK Signalosome. Mol. Cell 2008, 30, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Briggs, L.C.; Chan, A.W.E.; Davis, C.A.; Whitelock, N.; Hotiana, H.A.; Baratchian, M.; Bagnéris, C.; Selwood, D.L.; Collins, M.K.; Barrett, T.E. IKKγ-Mimetic Peptides Block the Resistance to Apoptosis Associated with Kaposi’s Sarcoma-Associated Herpesvirus Infection. J. Virol. 2017, 91, e01170-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.X.; Cusano, T.; Yuan, Y. Identification of the Immediate-Early Transcripts of Kaposi’s Sarcoma-Associated Herpesvirus. J. Virol. 1999, 73, 5556–5567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purushothaman, P.; Uppal, T.; Verma, S.C. Molecular Biology of KSHV Lytic Reactivation. Viruses 2015, 7, 116–153. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Lin, S.-F.; Gradoville, L.; Yuan, Y.; Zhu, F.; Miller, G. A viral gene that activates lytic cycle expression of Kaposi’s sarcoma-associated herpesvirus. Proc. Natl. Acad. Sci. USA 1998, 95, 10866–10871. [Google Scholar] [CrossRef] [Green Version]
- Long, W.; Zhao, G.; Wu, Y.; Liu, Y. Gallic acid inhibits Kaposi’s Sarcoma-associated herpesvirus lytic reactivation by suppressing RTA transcriptional activities. Food Sci. Nutr. 2020, 9, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Dorjsuren, D.; Burnette, A.; Gray, G.N.; Chen, X.; Zhu, W.; Roberts, P.E.; Currens, M.J.; Shoemaker, R.H.; Ricciardi, R.P.; Sei, S. Chemical library screen for novel inhibitors of Kaposi’s sarcoma-associated herpesvirus processive DNA synthesis. Antivir. Res. 2006, 69, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Bryant, K.F.; Gregory, S.M.; Angelova, M.; Dreyfus, D.H.; Zhao, X.Z.; Coen, D.M.; Burke, T.R.; Knipe, D.M. HIV Integrase Inhibitors Block Replication of Alpha-, Beta-, and Gammaherpesviruses. mBio 2014, 5, e01318-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.T.; Zhao, H.; Masaoka, T.; Varnado, B.; Castro, E.M.C.; Marshall, V.A.; Kouhestani, K.; Lynn, A.Y.; Aron, K.E.; Xia, A.; et al. Sensitivity of the C-Terminal Nuclease Domain of Kaposi’s Sarcoma-Associated Herpesvirus ORF29 to Two Classes of Active-Site Ligands. Antimicrob. Agents Chemother. 2018, 62, e00233-18. [Google Scholar] [CrossRef] [Green Version]
- Newcomb, W.W.; Brown, J.C. Inhibition of Herpes Simplex Virus Replication by WAY-150138: Assembly of Capsids Depleted of the Portal and Terminase Proteins Involved in DNA Encapsidation. J. Virol. 2002, 76, 10084–10088. [Google Scholar] [CrossRef] [Green Version]
- Visalli, R.J.; Fairhurst, J.; Srinivas, S.; Hu, W.; Feld, B.; DiGrandi, M.; Curran, K.; Ross, A.; Bloom, J.D.; van Zeijl, M.; et al. Identification of Small Molecule Compounds That Selectively Inhibit Varicella-Zoster Virus Replication. J. Virol. 2003, 77, 2349–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visalli, R.J.; Van Zeijl, M. DNA encapsidation as a target for anti-herpesvirus drug therapy. Antivir. Res. 2003, 59, 73–87. [Google Scholar] [CrossRef]
- Kornfeind, E.M.; Visalli, R.J. Human herpesvirus portal proteins: Structure, function, and antiviral prospects. Rev. Med. Virol. 2018, 28, e1972. [Google Scholar] [CrossRef]
- Goldner, T.; Hewlett, G.; Ettischer, N.; Ruebsamen-Schaeff, H.; Zimmermann, H.; Lischka, P. The Novel Anticytomegalovirus Compound AIC246 (Letermovir) Inhibits Human Cytomegalovirus Replication through a Specific Antiviral Mechanism That Involves the Viral Terminase. J. Virol. 2011, 85, 10884–10893. [Google Scholar] [CrossRef] [Green Version]
- Unal, A.; Pray, T.R.; Lagunoff, M.; Pennington, M.W.; Ganem, D.; Craik, C.S. The protease and the assembly protein of Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8). J. Virol. 1997, 71, 7030–7038. [Google Scholar] [CrossRef] [Green Version]
- Shahian, T.; Lee, G.M.; Lazic, A.; Arnold, L.A.; Velusamy, P.; Roels, C.M.; Guy, R.K.; Craik, C.S. Inhibition of a viral enzyme by a small-molecule dimer disruptor. Nat. Chem. Biol. 2009, 5, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-R.; Brulois, K.; Wong, L.; Jung, J.U. Modulation of Immune System by Kaposi’s Sarcoma-Associated Herpesvirus: Lessons from Viral Evasion Strategies. Front. Microbiol. 2012, 3, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acker, T.M.; Gable, J.E.; Bohn, M.F.; Jaishankar, P.; Thompson, M.C.; Fraser, J.S.; Renslo, A.R.; Craik, C.S. Allosteric Inhibitors, Crystallography, and Comparative Analysis Reveal Network of Coordinated Movement across Human Herpesvirus Proteases. J. Am. Chem. Soc. 2017, 139, 11650–11653. [Google Scholar] [CrossRef] [PubMed]
- Gantt, S.; Carlsson, J.; Ikoma, M.; Gachelet, E.; Gray, M.; Geballe, A.P.; Corey, L.; Casper, C.; Lagunoff, M.; Vieira, J. The HIV Protease Inhibitor Nelfinavir Inhibits Kaposi’s Sarcoma-Associated Herpesvirus Replication In Vitro. Antimicrob. Agents Chemother. 2011, 55, 2696–2703. [Google Scholar] [CrossRef] [Green Version]
- Gantt, S.; Cattamanchi, A.; Krantz, E.; Magaret, A.; Selke, S.; Kuntz, S.R.; Huang, M.-L.; Corey, L.; Wald, A.; Casper, C. Reduced human herpesvirus-8 oropharyngeal shedding associated with protease inhibitor-based antiretroviral therapy. J. Clin. Virol. 2014, 60, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Berger, E.A. An Immunotoxin Targeting the GH Glycoprotein of KSHV for Selective Killing of Cells in the Lytic Phase of Infection. Antivir. Res. 2011, 90, 143–1450. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, D.; Chandran, B.; Berger, E.A. Selective Killing of Kaposi’s Sarcoma-Associated Herpesvirus Lytically Infected Cells with a Recombinant Immunotoxin Targeting the Viral GpK8.1A Envelope Glycoprotein. MAbs 2012, 4, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Heston, L.; Grogan, E.; Gradoville, L.; Rigsby, M.; Sun, R.; Shedd, D.; Kushnaryov, V.M.; Grossberg, S.; Chang, Y. Selective switch between latency and lytic replication of Kaposi’s sarcoma herpesvirus and Epstein-Barr virus in dually infected body cavity lymphoma cells. J. Virol. 1997, 71, 314–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Ueda, K.; Sakakibara, S.; Okuno, T.; Parravicini, C.; Corbellino, M.; Yamanishi, K. Activation of latent Kaposi’s sarcoma-associated herpesvirus by demethylation of the promoter of the lytic transactivator. Proc. Natl. Acad. Sci. USA 2001, 98, 4119–4124. [Google Scholar] [CrossRef] [Green Version]
- Whitby, D.; Marshall, V.A.; Bagni, R.K.; Miley, W.J.; McCloud, T.G.; Hines-Boykin, R.; Goedert, J.J.; Conde, B.A.; Nagashima, K.; Mikovits, J.; et al. Reactivation of Kaposi’s sarcoma-associated herpesvirus by natural products from Kaposi’s sarcoma endemic regions. Int. J. Cancer 2006, 120, 321–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; He, M.; Zhou, F.; Ye, F.; Gao, S.-J. Activation of Kaposi’s Sarcoma-Associated Herpesvirus (KSHV) by Inhibitors of Class III Histone Deacetylases: Identification of Sirtuin 1 as a Regulator of the KSHV Life Cycle. J. Virol. 2014, 88, 6355–6367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Qin, Z.; Riker, A.I.; Xi, Y. CRISPR/Cas9 ablating viral microRNA promotes lytic reactivation of Kaposi’s sarcoma-associated herpesvirus. Biochem. Biophys. Res. Commun. 2020, 533, 1400–1405. [Google Scholar] [CrossRef]
- Iida, S.; Mine, S.; Ueda, K.; Suzuki, T.; Hasegawa, H.; Katano, H. Suberoyl Bis-Hydroxamic Acid Reactivates Kaposi’s Sarcoma-Associated Herpesvirus through Histone Acetylation and Induces Apoptosis in Lymphoma Cells. J. Virol. 2021, 95, e01785-20. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, E.; Anderton, M.; Blumenthal, M.; Schäfer, G. Cellular Receptors Involved in KSHV Infection. Viruses 2021, 13, 118. [Google Scholar] [CrossRef]
- Koon, H.B.; Bubley, G.J.; Pantanowitz, L.; Masiello, D.; Smith, B.; Crosby, K.; Proper, J.; Weeden, W.; Miller, T.E.; Chatis, P.; et al. Imatinib-Induced Regression of AIDS-Related Kaposi’s Sarcoma. J. Clin. Oncol. 2005, 23, 982–989. [Google Scholar] [CrossRef]
- Koon, H.B.; Krown, S.E.; Lee, J.Y.; Honda, K.; Rapisuwon, S.; Wang, Z.; Aboulafia, D.; Reid, E.G.; Rudek, M.A.; Dezube, B.J.; et al. Phase II Trial of Imatinib in AIDS-Associated Kaposi’s Sarcoma: AIDS Malignancy Consortium Protocol 042. J. Clin. Oncol. 2014, 32, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Ardavanis, A.; Doufexis, D.; Kountourakis, P.; Rigatos, G. A Kaposi’s Sarcoma Complete Clinical Response after Sorafenib Administration. Ann. Oncol. 2008, 19, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Uldrick, T.S.; Gonçalves, P.H.; Wyvill, K.M.; Peer, C.J.; Bernstein, W.; Aleman, K.; Polizzotto, M.; Venzon, D.; Steinberg, S.M.; Marshall, V.; et al. A Phase Ib Study of Sorafenib (BAY 43-9006) in Patients with Kaposi Sarcoma. Oncology 2017, 22, 505. [Google Scholar] [CrossRef] [Green Version]
- Gill, M.B.; Turner, R.; Stevenson, P.G.; Way, M. KSHV-TK is a tyrosine kinase that disrupts focal adhesions and induces Rho-mediated cell contraction. EMBO J. 2014, 34, 448–465. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.P.; Stuhlmiller, T.J.; Giffin, L.C.; Lin, C.; Bigi, R.; Zhao, J.; Zhang, W.; Cruz, A.G.B.; Park, S.I.; Earp, H.S.; et al. Kinome profiling of non-Hodgkin lymphoma identifies Tyro3 as a therapeutic target in primary effusion lymphoma. Proc. Natl. Acad. Sci. USA 2019, 116, 16541–16550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodhi, A.; Chaisuparat, R.; Hu, J.; Ramsdell, A.K.; Manning, B.D.; Sausville, E.A.; Sawai, E.T.; Molinolo, A.; Gutkind, J.S.; Montaner, S. The TSC2/mTOR pathway drives endothelial cell transformation induced by the Kaposi’s sarcoma-associated herpesvirus G protein-coupled receptor. Cancer Cell 2006, 10, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Nichols, L.A.; Adang, L.A.; Kedes, D.H. Rapamycin Blocks Production of KSHV/HHV8: Insights into the Anti-Tumor Activity of an Immunosuppressant Drug. PLoS ONE 2011, 6, e14535. [Google Scholar] [CrossRef] [PubMed]
- Sin, S.-H.; Roy, D.; Wang, L.; Staudt, M.R.; Fakhari, F.D.; Patel, D.D.; Henry, D.; Harrington, W.J.; Damania, B.A.; Dittmer, D.P. Rapamycin Is Efficacious against Primary Effusion Lymphoma (PEL) Cell Lines in Vivo by Inhibiting Autocrine Signaling. Blood 2007, 109, 2165–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, A.P.; Bhende, P.M.; Sin, S.H.; Roy, D.; Dittmer, D.P.; Damania, B. Dual Inhibition of PI3K and MTOR Inhibits Autocrine and Paracrine Proliferative Loops in PI3K/Akt/MTOR-Addicted Lymphomas. Blood 2010, 115, 4455–4463. [Google Scholar] [CrossRef]
- Caro-Vegas, C.; Bailey, A.; Bigi, R.; Damania, B.; Dittmer, D.P. Targeting MTOR with MLN0128 Overcomes Rapamycin and Chemoresistant Primary Effusion Lymphoma. MBio 2019, 10, e02871-18. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Xie, J.; Ye, F.; Gao, S.-J. Modulation of Kaposi’s Sarcoma-Associated Herpesvirus Infection and Replication by MEK/ERK, JNK, and p38 Multiple Mitogen-Activated Protein Kinase Pathways during Primary Infection. J. Virol. 2006, 80, 5371–5382. [Google Scholar] [CrossRef] [Green Version]
- Wakao, K.; Watanabe, T.; Takadama, T.; Ui, S.; Shigemi, Z.; Kagawa, H.; Higashi, C.; Ohga, R.; Taira, T.; Fujimuro, M. Sangivamycin induces apoptosis by suppressing Erk signaling in primary effusion lymphoma cells. Biochem. Biophys. Res. Commun. 2014, 444, 135–140. [Google Scholar] [CrossRef]
- Moriguchi, M.; Watanabe, T.; Kadota, A.; Fujimuro, M. Capsaicin Induces Apoptosis in KSHV-Positive Primary Effusion Lymphoma by Suppressing ERK and p38 MAPK Signaling and IL-6 Expression. Front. Oncol. 2019, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Huang, L.; Xiao, Y.; Yao, X.; Long, X.; Zhu, F.; Kuang, E. Development of an ORF45-Derived Peptide To Inhibit the Sustained RSK Activation and Lytic Replication of Kaposi’s Sarcoma-Associated Herpesvirus. J. Virol. 2019, 93, e02154-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.; Trillo-Tinoco, J.; Cao, Y.; Bonstaff, K.; Doyle, L.; del Valle, L.; Whitby, D.; Qin, Z. Targeting HGF/c-MET Induces Cell Cycle Arrest, DNA Damage, and Apoptosis for Primary Effusion Lymphoma. Blood J. Am. Soc. Hematol. 2015, 126, 2821–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayar, U.; Lu, P.; Vider, J.; Cerchietti, L.; Chiosis, G.; Wang, L.; Blasberg, R.; Cesarman, E. Hsp90 is a viable therapeutic target in the treatment of KSHV-associated primary effusion lymphoma. Infect. Agents Cancer 2010, 5, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Wen, K.W.; Damania, B. Kaposi sarcoma-associated herpesvirus (KSHV): Molecular biology and oncogenesis. Cancer Lett. 2010, 289, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Defee, M.; Isaacs, J.S.; Parsons, C. Extracellular Hsp90 serves as a co-factor for MAPK activation and latent viral gene expression during de novo infection by KSHV. Virology 2010, 403, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Higashi, C.; Saji, C.; Yamada, K.; Kagawa, H.; Ohga, R.; Taira, T.; Fujimuro, M. The Effects of Heat Shock Protein 90 Inhibitors on Apoptosis and Viral Replication in Primary Effusion Lymphoma Cells. Biol. Pharm. Bull. 2012, 35, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, R.; Matta, H.; Chaudhary, P.M. A Purine Scaffold HSP90 Inhibitor BIIB021 Has Selective Activity against KSHV-Associated Primary Effusion Lymphoma and Blocks vFLIP K13-Induced NF-κB. Clin. Cancer Res. 2013, 19, 5016–5026. [Google Scholar] [CrossRef] [Green Version]
- Qi, C.-F.; Kim, Y.-S.; Xiang, S.; Abdullaev, Z.; Torrey, T.A.; Janz, S.; Kovalchuk, A.L.; Sun, J.; Chen, D.; Cho, W.C.; et al. Characterization of ARF-BP1/HUWE1 Interactions with CTCF, MYC, ARF and p53 in MYC-Driven B Cell Neoplasms. Int. J. Mol. Sci. 2012, 13, 6204–6219. [Google Scholar] [CrossRef] [Green Version]
- Baquero-Pérez, B.; Whitehouse, A. Hsp70 Isoforms Are Essential for the Formation of Kaposi’s Sarcoma-Associated Herpesvirus Replication and Transcription Compartments. PLoS Pathog. 2015, 11, e1005274. [Google Scholar] [CrossRef] [Green Version]
- Hughes, D.; Wood, J.J.; Jackson, B.; Baquero-Perez, B.; Whitehouse, A. NEDDylation Is Essential for Kaposi’s Sarcoma-Associated Herpesvirus Latency and Lytic Reactivation and Represents a Novel Anti-KSHV Target. PLoS Pathog. 2015, 11, e1004771. [Google Scholar] [CrossRef] [Green Version]
- Matta, H.; Chaudhary, P.M. The proteasome inhibitor bortezomib (PS-341) inhibits growth and induces apoptosis in primary effusion lymphoma cells. Cancer Biol. Ther. 2005, 4, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Granato, M.; Romeo, M.A.; Tiano, M.S.; Santarelli, R.; Gonnella, R.; Montani, M.S.G.; Faggioni, A.; Cirone, M. Bortezomib promotes KHSV and EBV lytic cycle by activating JNK and autophagy. Sci. Rep. 2017, 7, 13052. [Google Scholar] [CrossRef] [Green Version]
- Sarosiek, K.; Cavallin, L.E.; Bhatt, S.; Toomey, N.L.; Natkunam, Y.; Blasini, W.; Gentles, A.J.; Ramos, J.C.; Mesri, E.A.; Lossos, I.S. Efficacy of bortezomib in a direct xenograft model of primary effusion lymphoma. Proc. Natl. Acad. Sci. USA 2010, 107, 13069–13074. [Google Scholar] [CrossRef] [Green Version]
- Reid, E.G.; Suazo, A.; Lensing, S.Y.; Dittmer, D.P.; Ambinder, R.F.; Maldarelli, F.; Gorelick, R.J.; Aboulafia, D.M.; Mitsuyasu, R.; Dickson, M.A.; et al. Pilot Trial AMC-063: Safety and Efficacy of Bortezomib in AIDS-associated Kaposi Sarcoma. Clin. Cancer Res. 2019, 26, 558–565. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, H.; Chen, X. Pemetrexed Inhibits Kaposi’s Sarcoma-Associated Herpesvirus Replication through Blocking DTMP Synthesis. Antivir. Res. 2020, 180, 104825. [Google Scholar] [CrossRef]
- Xu, B.; Wang, L.; González-Molleda, L.; Wang, Y.; Xu, J.; Yuan, Y. Antiviral Activity of (+)-Rutamarin against Kaposi’s Sarcoma-Associated Herpesvirus by Inhibition of the Catalytic Activity of Human Topoisomerase II. Antimicrob. Agents Chemother. 2013, 58, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Rinderknecht, A.S.; Zoeteweij, J.P.; Aoki, Y.; Read-Connole, E.L.; Tosato, G.; Blauvelt, A.; Yarchoan, R. Hypoxia Induces Lytic Replication of Kaposi Sarcoma–Associated Herpesvirus. Blood 2001, 97, 3244–3250. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.; Davis, D.A.; Wang, V.; Widmer, I.; Yarchoan, R. Kaposi’s Sarcoma-Associated Herpesvirus (Human Herpesvirus 8) Contains Hypoxia Response Elements: Relevance to Lytic Induction by Hypoxia. J. Virol. 2003, 77, 6761–6768. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.; Wang, V.; Davis, D.A.; Zheng, Z.-M.; Yarchoan, R. Genetic Organization and Hypoxic Activation of the Kaposi’s Sarcoma-Associated Herpesvirus ORF34-37 Gene Cluster. J. Virol. 2006, 80, 7037–7051. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, P.; Davis, D.A.; Veeranna, R.P.; Carey, R.F.; Viollet, C.; Yarchoan, R. Hypoxia-inducible factor-1 alpha as a therapeutic target for primary effusion lymphoma. PLoS Pathog. 2017, 13, e1006628. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Tan, B.; Vasan, K.; Yuan, H.; Cheng, F.; da Silva, S.R.; Lu, C.; Gao, S.-J. SIRT1 and AMPK pathways are essential for the proliferation and survival of primary effusion lymphoma cells. J. Pathol. 2017, 242, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Qiao, J.; Nguyen, D.; Struckhoff, A.P.; Doyle, L.; Bonstaff, K.; Del Valle, L.; Parsons, C.; Toole, B.P.; Renne, R.; et al. Role of heme oxygenase-1 in the pathogenesis and tumorigenicity of Kaposi’s sarcoma-associated herpesvirus. Oncotarget 2016, 7, 10459–10471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uldrick, T.S.; Wyvill, K.M.; Kumar, P.; O’Mahony, D.; Bernstein, W.; Aleman, K.; Polizzotto, M.; Steinberg, S.M.; Pittaluga, S.; Marshall, V.; et al. Phase II Study of Bevacizumab in Patients With HIV-Associated Kaposi’s Sarcoma Receiving Antiretroviral Therapy. J. Clin. Oncol. 2012, 30, 1476–1483. [Google Scholar] [CrossRef] [PubMed]
- Ramaswami, R.; Uldrick, T.S.; Polizzotto, M.; Wyvill, K.M.; Goncalves, P.; Widell, A.; Lurain, K.; Steinberg, S.M.; Figg, W.D.; Tosato, G.; et al. A Pilot Study of Liposomal Doxorubicin Combined with Bevacizumab followed by Bevacizumab Monotherapy in Patients with Advanced Kaposi Sarcoma. Clin. Cancer Res. 2019, 25, 4238–4247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uldrick, T.S.; Polizzotto, M.; Aleman, K.; Wyvill, K.M.; Marshall, V.; Whitby, D.; Wang, V.; Pittaluga, S.; O’Mahony, D.; Steinberg, S.M.; et al. Rituximab plus liposomal doxorubicin in HIV-infected patients with KSHV-associated multicentric Castleman disease. Blood 2014, 124, 3544–3552. [Google Scholar] [CrossRef] [Green Version]
- Marcelin, A.-G.; Aaron, L.; Mateus, C.; Gyan, E.; Gorin, I.; Viard, J.-P.; Calvez, V.; Dupin, N. Rituximab therapy for HIV-associated Castleman disease. Blood 2003, 102, 2786–2788. [Google Scholar] [CrossRef] [Green Version]
- Neuville, S.; Agbalika, F.; Rabian, C.; Brière, J.; Molina, J.-M. Failure of rituximab in human immunodeficiency virus-associated multicentric Castleman disease. Am. J. Hematol. 2005, 79, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Nagao, A.; Nakazawa, S.; Hanabusa, H. Short-Term Efficacy of the IL6 Receptor Antibody Tocilizumab in Patients with HIV-Associated Multicentric Castleman Disease: Report of Two Cases. J. Hematol. Oncol. 2014, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Aita, T.; Hamaguchi, S.; Shimotani, Y.; Nakamoto, Y. Idiopathic Multicentric Castleman Disease Preceded by Cutaneous Plasmacytosis Successfully Treated by Tocilizumab. BMJ Case Rep. 2020, 13, e236283. [Google Scholar] [CrossRef]
- Ramaswami, R.; Lurain, K.; Peer, C.J.; Serquiña, A.; Wang, V.Y.; Widell, A.; Goncalves, P.; Steinberg, S.M.; Marshall, V.; George, J.; et al. Tocilizumab in patients with symptomatic Kaposi sarcoma herpesvirus–associated multicentric Castleman disease. Blood 2020, 135, 2316–2319. [Google Scholar] [CrossRef]
- Moodad, S.; El Hajj, R.; Hleihel, R.; Hajjar, L.; Tawil, N.; Karam, M.; Hamie, M.; Merhi, R.A.; El Sabban, M.; El Hajj, H. Lenalidomide in Combination with Arsenic Trioxide: An Effective Therapy for Primary Effusion Lymphoma. Cancers 2020, 12, 2483. [Google Scholar] [CrossRef]
- Shrestha, P.; Davis, D.A.; Jaeger, H.K.; Stream, A.; Aisabor, A.I.; Yarchoan, R. Pomalidomide restores immune recognition of primary effusion lymphoma through upregulation of ICAM-1 and B7-2. PLoS Pathog. 2021, 17, e1009091. [Google Scholar] [CrossRef]
- Polizzotto, M.; Uldrick, T.S.; Wyvill, K.M.; Aleman, K.; Peer, C.J.; Bevans, M.; Sereti, I.; Maldarelli, F.; Whitby, D.; Marshall, V.; et al. Pomalidomide for Symptomatic Kaposi’s Sarcoma in People With and Without HIV Infection: A Phase I/II Study. J. Clin. Oncol. 2016, 34, 4125–4131. [Google Scholar] [CrossRef]
- Neil, R.; Halaby, T.; Weverling, G.J.; Dukers, N.H.T.M.; Simpson, G.R.; Coutinho, R.A.; Lange, J.M.A.; Schulz, T.F.; Goudsmit, J. Seroconversion for Human Herpesvirus 8 during HIV Infection Is Highly Predictive of Kaposi’s Sarcoma. AIDS 1998, 12, 2481–2488. [Google Scholar]
- Blasig, C.; Zietz, C.; Haar, B.; Neipel, F.; Esser, S.; Brockmeyer, N.H.; Tschachler, E.; Colombini, S.; Ensoli, B.; Stürzl, M. Monocytes in Kaposi’s Sarcoma Lesions Are Productively Infected by Human Herpesvirus 8. J. Virol. 1997, 71, 7963–7968. [Google Scholar] [CrossRef] [Green Version]
- Boshoff, C.; Schulz, T.F.; Kennedy, M.M.; Graham, A.K.; Fisher, C.; Thomas, A.; McGee, J.O.; Weiss, R.A.; O’Leary, J.J. Kaposi’s Sarcoma-Associated Herpesvirus Infects Endothelial and Spindle Cells. Nat. Med. 1995, 1, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Flore, O.; Rafii, S.; Ely, S.; O’Leary, J.J.; Hyjek, E.M.; Cesarman, E. Transformation of primary human endothelial cells by Kaposi’s sarcoma-associated herpesvirus. Nature 1998, 394, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Diamond, C.; Brodie, S.J.; Krieger, J.N.; Huang, M.-L.; Koelle, D.M.; Diem, K.; Muthui, D.; Corey, L. Human Herpesvirus 8 in the Prostate Glands of Men with Kaposi’s Sarcoma. J. Virol. 1998, 72, 6223–6227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moses, A.V.; Fish, K.N.; Ruhl, R.; Smith, P.P.; Strussenberg, J.G.; Zhu, L.; Chandran, B.; Nelson, J.A. Long-Term Infection and Transformation of Dermal Microvascular Endothelial Cells by Human Herpesvirus 8. J. Virol. 1999, 73, 6892–6902. [Google Scholar] [CrossRef] [Green Version]
- Blackbourn, D.J.; Lennette, E.; Klencke, B.; Moses, A.; Chandran, B.; Weinstein, M.; Glogau, R.G.; Levy, J.A. The Restricted Cellular Host Range of Human Herpesvirus 8. AIDS 2000, 14, 1123–1133. [Google Scholar] [CrossRef]
- Chandran, B.; Hutt-Fletcher, L. Gammaherpesviruses entry and early events during infection. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Bechtel, J.T.; Liang, Y.; Hvidding, J.; Ganem, D. Host Range of Kaposi’s Sarcoma-Associated Herpesvirus in Cultured Cells. J. Virol. 2003, 77, 6474–6481. [Google Scholar] [CrossRef] [Green Version]
- Ojala, P.M.; Schulz, T. Manipulation of endothelial cells by KSHV: Implications for angiogenesis and aberrant vascular differentiation. Semin. Cancer Biol. 2014, 26, 69–77. [Google Scholar] [CrossRef]
- Dubich, T.; Dittrich, A.; Bousset, K.; Geffers, R.; Büsche, G.; Köster, M.; Hauser, H.; Schulz, T.F.; Wirth, D. 3D culture conditions support Kaposi’s sarcoma herpesvirus (KSHV) maintenance and viral spread in endothelial cells. J. Mol. Med. 2021, 99, 425–438. [Google Scholar] [CrossRef]
- Fujiwara, S.; Nakamura, H. Animal Models for Gammaherpesvirus Infections: Recent Development in the Analysis of Virus-Induced Pathogenesis. Pathogens 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.; Wachtman, L.M.; Pearson, C.B.; Lee, J.-S.; Lee, H.-R.; Lee, S.H.; Vieira, J.; Mansfield, K.G.; Jung, J.U. Non-Human Primate Model of Kaposi’s Sarcoma-Associated Herpesvirus Infection. PLoS Pathog. 2009, 5, e1000606. [Google Scholar] [CrossRef] [Green Version]
- Orzechowska, B.; Powers, M.F.; Sprague, J.; Li, H.; Yen, B.; Searles, R.P.; Axthelm, M.K.; Wong, S.W. Rhesus macaque rhadinovirus-associated non-Hodgkin lymphoma: Animal model for KSHV-associated malignancies. Blood 2008, 112, 4227–4234. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.U.; Choi, J.-K.; Ensser, A.; Biesinger, B. Herpesvirus saimiri as a model for gammaherpesvirus oncogenesis. Semin. Cancer Biol. 1999, 9, 231–239. [Google Scholar] [CrossRef]
- Efstathiou, S.; Ho, Y.M.; Minson, A.C. Cloning and molecular characterization of the murine herpesvirus 68 genome. J. Gen. Virol. 1990, 71, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Virgin, H.; Latreille, P.; Wamsley, P.; Hallsworth, K.; Weck, E.K.; Canto, A.J.D.; Speck, S.H. Complete sequence and genomic analysis of murine gammaherpesvirus 68. J. Virol. 1997, 71, 5894–5904. [Google Scholar] [CrossRef] [Green Version]
- Louna, K.; Houshaymi, B.; Abdel-Samad, R.; Jaafar, M.; Halloum, I.; Pisano, C.; Neipel, F.; Darwiche, N.; Merhi, R.A. Antitumor Activity of the Synthetic Retinoid ST1926 on Primary Effusion Lymphoma in Vitro and in Vivo Models. Oncol. Rep. 2018, 39, 721–730. [Google Scholar] [CrossRef] [Green Version]
- McHugh, D.; Caduff, N.; Barros, M.H.M.; Rämer, P.C.; Raykova, A.; Murer, A.; Landtwing, V.; Quast, I.; Styles, C.; Spohn, M.; et al. Persistent KSHV Infection Increases EBV-Associated Tumor Formation In Vivo via Enhanced EBV Lytic Gene Expression. Cell Host Microbe 2017, 22, 61–73.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Naimo, E.; Zischke, J.; Schulz, T.F. Recent Advances in Developing Treatments of Kaposi’s Sarcoma Herpesvirus-Related Diseases. Viruses 2021, 13, 1797. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091797
AMA Style
Naimo E, Zischke J, Schulz TF. Recent Advances in Developing Treatments of Kaposi’s Sarcoma Herpesvirus-Related Diseases. Viruses. 2021; 13(9):1797. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091797
Chicago/Turabian StyleNaimo, Eleonora, Jasmin Zischke, and Thomas F. Schulz. 2021. "Recent Advances in Developing Treatments of Kaposi’s Sarcoma Herpesvirus-Related Diseases" Viruses 13, no. 9: 1797. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091797
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.