
The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes


1
Center for Grain and Animal Health Research, Agricultural Research Service, United States Department of Agriculture, 1515 College Ave., Manhattan, KS 66502, USA
2
National Center for Emerging and Zoonotic Infectious Diseases, Centers for Disease Control and Prevention, Division of Vector-Borne Diseases, 3156 Rampart Road, Fort Collins, CO 80521, USA
*
Author to whom correspondence should be addressed.
Viruses 2021, 13(9), 1822; https://0-doi-org.brum.beds.ac.uk/10.3390/v13091822
Submission received: 11 August 2021
/
Revised: 10 September 2021
/
Accepted: 11 September 2021
/
Published: 14 September 2021
(This article belongs to the Special Issue Mosquito-Borne Virus Ecology)
Abstract
:Temperature plays a significant role in the vector competence, extrinsic incubation period, and intensity of infection of arboviruses within mosquito vectors. Most laboratory infection studies use static incubation temperatures that may not accurately reflect daily temperature ranges (DTR) to which mosquitoes are exposed. This could potentially compromise the application of results to real world scenarios. We evaluated the effect of fluctuating DTR versus static temperature treatments on the infection, dissemination, and transmission rates and viral titers of Culex tarsalis and Culex quinquefasciatus mosquitoes for West Nile virus. Two DTR regimens were tested including an 11 and 15 °C range, both fluctuating around an average temperature of 28 °C. Overall, no significant differences were found between DTR and static treatments for infection, dissemination, or transmission rates for either species. However, significant treatment differences were identified for both Cx. tarsalis and Cx. quinquefasciatus viral titers. These effects were species-specific and most prominent later in the infection. These results indicate that future studies on WNV infections in Culex mosquitoes should consider employing realistic DTRs to reflect interactions most accurately between the virus, vector, and environment.
1. Introduction
The epidemiology of vector-borne disease is inherently complex, involving multifaceted interactions between vectors, hosts, and pathogens occurring within a constantly changing environment. Numerous environmental variables impact transmission dynamics, but perhaps none greater than temperature [1]. Temperature is a factor in the calculation of metrics associated with the vector potential of an organism or population, including the extrinsic incubation period and vectorial capacity [2,3]. Aspects of vector biology, such as development time, body size, fecundity, and survival rates, are also closely associated with temperature [4,5,6,7,8]. Additionally, there is increasing evidence that gene expression of functions such as metabolism, immunity, and detoxification have a circadian component in mosquitoes [9], with light and temperature often being drivers of circadian rhythms [10]. Due to this close association between temperature and vector-borne disease, climate change is anticipated to shift the range of vectors and vector-borne diseases in the future [11,12,13,14]. Consequently, variations in temperatures used for laboratory vector competence assays have implications for real-world applications of results.
Vector competence studies investigate whether a suspected vector species can become infected with and later transmit a pathogen [15,16]. Most vector competence assays are conducted at a static temperature representing conditions expected within a region of interest [17,18,19]. However, these static temperature trials do not take into consideration circadian cycles that an insect vector would experience in nature [20,21]. Previously conducted experiments using daily temperature ranges (DTRs) rather than static temperatures reveal the significant impacts that these cycles can have. Aedes aegypti infected with dengue virus (DENV) serotypes 1 or 2 and held in a large DTR (20 °C) setting were found to have decreased rates of infection compared to those in a small DTR (10 °C) setting [22]. Additional studies from this system also revealed a significant effect of the mean temperature around which the DTR was fluctuating [23]. Similarly, fluctuating temperatures have been shown to impact Plasmodium infection in Anopheles stephensi, with variable outcomes depending on mean temperature [24].
Since its introduction in 1999, West Nile virus (WNV) has become the most prevalent mosquito-borne disease in the United States, with cases occurring in the continental US annually [25]. The United States is composed of a broad diversity of unique regions with variable average temperatures and DTRs, many of which report annual WNV activity. For example, the Mojave and Sonoran deserts of the Southwestern United States are home to Las Vegas, Nevada and Phoenix, Arizona-cities that experienced WNV outbreaks in 2019 (37 and 153 human cases, respectively [25]). These cities experience broad daily temperature fluctuations of approximately 13 °C during peak WNV transmission periods of July-September [26]. Similar temperature fluctuations are seen in other Southern cities of the United States, such as Houston, Texas where average daily temperatures experience 11 °C fluctuations. Despite significant daily temperature ramps and evidence that temperature significantly impacts transmission of WNV [27], we have little data on the impact of these circadian patterns on Culex vector competence and viral load for WNV. The purpose of this study was to compare the infection, dissemination, and transmission rates and viral titers of WNV in two mosquito vector species that occur in these regions, Cx. quinquefasciatus and Cx. tarsalis, when exposed to DTR conditions reflecting these southern US localities.
2. Materials and Methods
2.1. Study Insects
Two mosquito species were reared for these experiments: Culex quinquefasciatus and Culex tarsalis. The Cx. quinquefasciatus came from the Sebring colony originally colonized in 1988 from a population in Florida. The Cx. tarsalis were from the Bakersfield colony established in 1952 from a population present in Bakersfield, CA. Both species were reared in incubators held at 27.5 °C on a 12:12 L:D circadian cycle and provided 10% sugar solution ad libitum. On the day prior to infections, mosquitoes were transferred into 16 oz paper cups covered with a double layer of mesh and moved into the BSL-3 laboratory to acclimate to the new incubators, temperature regimens, and collection cups for 24 h. Mosquitoes were provided clean water only in the 24 h prior to blood feeding to improve blood feeding success.
2.2. Fluctuating DTR Conditions
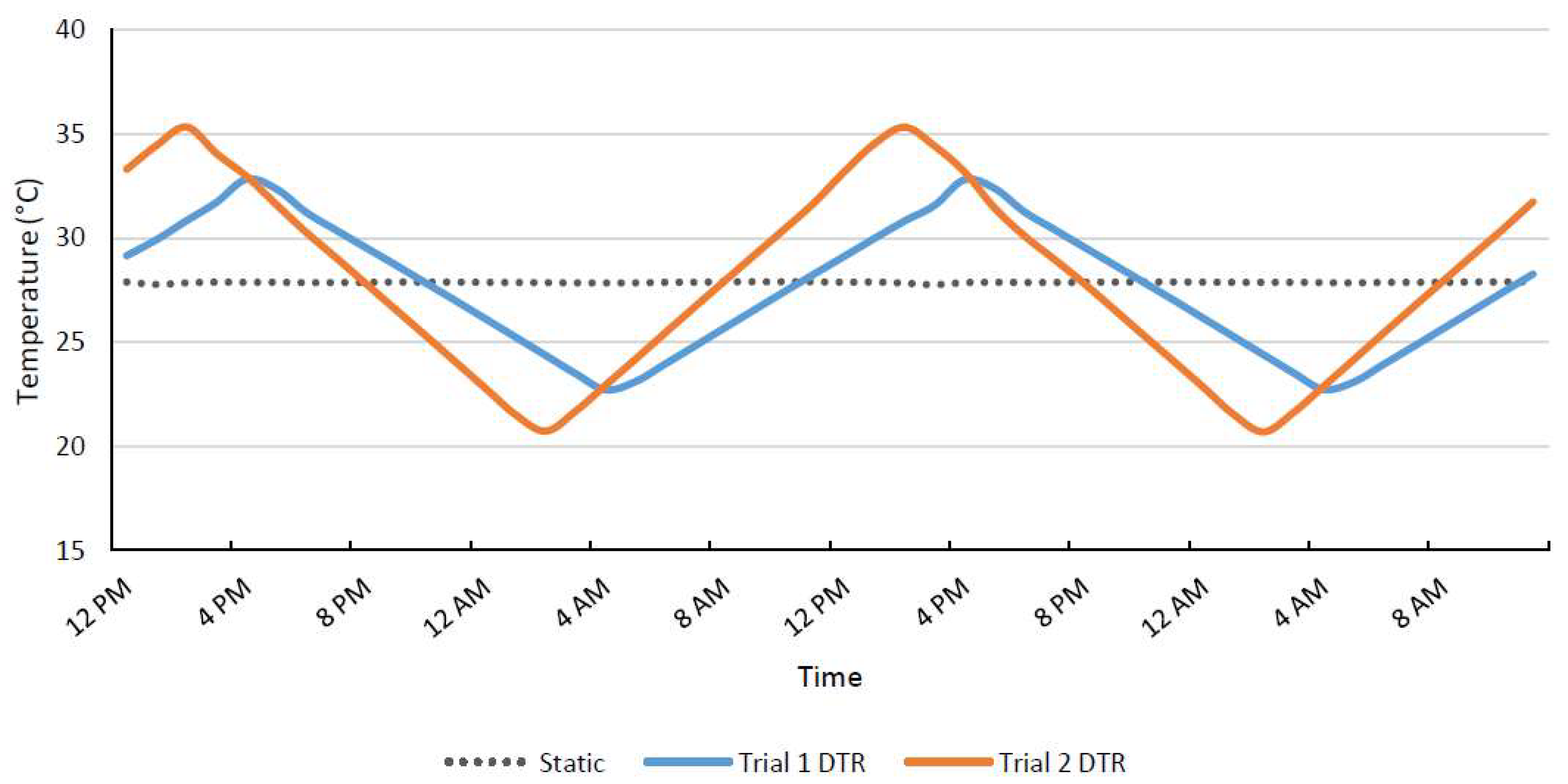
Two temperature trials were completed in which one incubator was set to oscillate temperatures gradually between a high and low temperature within a 24 h period (DTR treatment) and a second incubator was set to hold a stable temperature. Both incubators were Thermo Scientific Forma Environmental Chambers (Thermo Fisher Scientific, Waltham, MA, USA), and the DTR treatment incubator was also fitted with a temperature controller (Watlow Electric Manufacturing Co., St. Louis, MO, USA). The first trial used a DTR of 11 °C from 22.5 to 33.5 °C in one incubator (increasing at a rate of 0.015 °C/min for 12 h followed by a decrease of 0.015 °C/min for 12 h) and a static temperature of 28 °C in the other incubator (Figure 1). For the second trial, a DTR of 15 °C (20.5–35.5 °C) was used in the fluctuating incubator (increasing at a rate of 0.021 °C/min for 12 h followed by decreasing at 0.021 °C/min for 12 h) and the static incubator was set to 28 °C. Hobo monitors (Onset Computer Corporation, Bourne, MA, USA) were used throughout the experiment to monitor temperatures in both incubators.
2.3. Infection Trials
The viral strain of WNV used for this study was isolated from mosquitoes collected in Fort Collins, CO, USA in 2016. The stock virus was passaged twice on Vero cells and then mixed with defibrinated goose blood (Colorado Serum Company, Denver, CO, USA) to produce a final infectious blood titer of 7.0 log10 PFU/mL for both blood feedings in trial one and for the Cx. quinquefasciatus in trial 2. In trial 2, the infectious titer fed to Cx. tarsalis was reduced to 6.5 log10 PFU/mL in response to high infection rates observed in trial 1. Mosquitoes were fed infectious blood through a Parafilm membrane using a Hemotek blood-feeding system (Hemotek Ltd., Blackburn, UK) set to warm the blood to 37 °C. Mosquitoes were left to blood feed inside an incubator set at 28 °C for one hour. At the end of the blood feeding period, cups of mosquitoes were anesthetized by exposure to −20 °C for 45 s followed by placing anesthetized mosquitoes in a petri dish on ice. Blood fed mosquitoes were sorted into fresh 16 oz cups of <50 individuals while unfed and male mosquitoes were discarded.
Mosquitoes were monitored for mortality and administered fresh 10% sugar solution daily. Collections were made from both treatments and both species on days 4 and 8 post infection. An additional timepoint was added for Cx. quinquefasciatus at 12 days post infection in trial 2, but this timepoint was not added for Cx. tarsalis due to poor survival. Prior to collections, mosquitoes were anesthetized with triethylamine (TEA) (Thermo Fisher Scientific). Legs were dissected from each individual and collected into 2 mL microcentrifuge tubes containing 500 µL complete media (Dulbecco’s Modified Eagle Medium (Gibco, Thermo Fisher Scientific) supplemented with 10% fetal bovine serum (Omega Scientific Inc, Tarzana, CA, USA), 2% penicillin streptomycin solution (Gibco, Thermo Fisher Scientific), and 0.2% Amphotericin B solution (Gibco, Thermo Fisher Scientific)) and 4–8 2 mm zirconium oxide beads (Glen Mills Inc., Clifton, NJ, USA). Capillary assays were then conducted to collect mosquito saliva [28]. Capillary tubes were collected into 2 mL microcentrifuge tubes containing 300 µL complete media. Mosquito bodies were then collected into 2 mL microcentrifuge tubes containing 500 µL complete media and 4–8 2 mm zirconium oxide beads.
2.4. Sample Testing
Samples were homogenized and tested for virus using established protocols [29]. Plaque assays were conducted on Vero cells for all body samples. Legs were then tested for individuals with positive bodies followed by plaque assays on saliva samples for those individuals with positive legs. Serial dilutions in complete media were used when necessary to titrate samples for plaque assays.
2.5. Statistical Analyses
Infection rates were calculated as the total number of individuals with positive bodies divided by the total number of individuals that successfully blood fed on the infectious blood meal. Dissemination rates were calculated as the total number of individuals with positive legs divided by the total number of individuals with positive bodies. Transmission rates were calculated as the total number of individuals with positive saliva divided by the total number with positive bodies.
Infection, dissemination, and transmission rates were assessed by Fisher’s exact tests for both species and trials. Shapiro–Wilk tests were used to determine whether virus titer data followed a normal distribution. Titer data were determined to be non-normally distributed, so Wilcoxon–Mann–Whitney non-parametric tests were used to assess significant differences in viral titers between treatments in both experiments and for both species. Fisher’s exact tests and Wilcoxon–Mann–Whitney tests were also used to assess differences in the infection, dissemination, and transmission rates and viral titers between DTRs for Cx. quinquefasciatus.
3. Results
3.1. Trial 1: 11 °C DTR
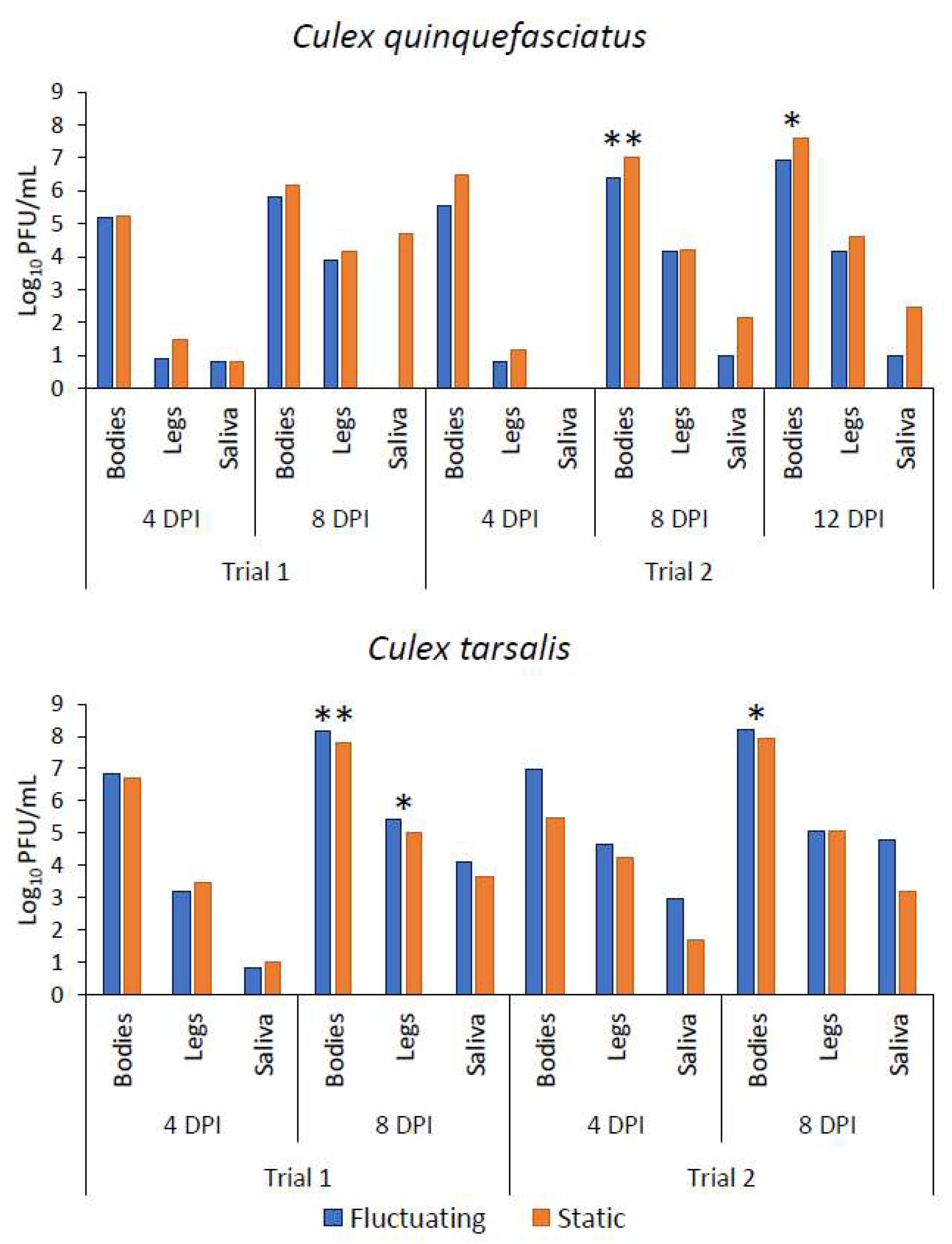
There were no significant differences identified in infection, dissemination, or transmission rates or in viral titers between treatments for Cx. quinquefasciatus in trial 1 at the 4 or 8 days post-infection (DPI) timepoint (Table 1 and Table 2). Too few Cx. quinquefasciatus individuals had positive saliva to produce a robust statistical analysis on salivary titers. Culex quinquefasciatus viral titers were higher overall in the static treatment than in the fluctuating treatment in trial 1 (Figure 2).
For Cx. tarsalis, Fisher’s exact results indicated that there was no difference between treatments in infection rates of bodies at 4 or 8 DPI (Table 1). There was also no statistical difference between titers of body samples at 4 DPI; however, a significant difference between body titers at 8 DPI was observed (Table 2). There were no differences between the two treatments in dissemination rates at either timepoint (Table 1). At day 4 post infection, there was no difference in viral titers of legs; however, a significant difference was identified in leg viral titers between treatments at 8 DPI (Table 2). No difference was identified in transmission rates between the two treatments at either timepoint (Table 1). There were too few positive saliva samples to run a robust statistical analysis on salivary titer results for 4 DPI. At 8 DPI, there was no difference identified in salivary titers between treatments (Table 2). Overall, viral titers for Cx. tarsalis were higher in the fluctuating group than the static group except for the 4 DPI legs and saliva (Figure 2).
3.2. Trial 2: 15 °C DTR
No treatment effect was observed for the infection, dissemination, or transmission rates of Cx. quinquefasciatus at any of the three timepoints for trial 2 (Table 1). At 4 DPI, no difference was observed between treatments in body titers or leg titers (Table 2). There were no positive saliva samples at 4 DPI to run titer analyses. At 8 DPI, a significantly higher viral titer was observed in body tissues of insects exposed to static temperatures. No difference was observed at 8 DPI for leg titers or saliva titers (Table 2). Similarly, at 12 DPI, significantly higher viral titers were observed in the body tissues of individuals in the static treatment (Table 2). Leg titers and saliva titers again showed no difference between treatments (Table 2). Similar to trial 1, Cx. quinquefasciatus had higher viral titers overall in the static treatment (Figure 2).
There was no treatment effect identified for Cx. tarsalis for infection, dissemination, or transmission rates (Table 1). No difference was identified in body titers or leg titers at 4 DPI, with too few positive saliva samples to run statistical tests. At 8 DPI, body titers were significantly higher in the fluctuating treatment group; however, leg titers and saliva titers were not statistically different at 8 DPI for Cx. tarsalis (Table 2). The overall titers for Cx. tarsalis were higher in the fluctuating group than in the static group in trial 2 except in the 8 DPI legs (Figure 2).
3.3. Trial Comparison—Culex quinquefasciatus
Culex quinquefasciatus was exposed to the same viral titer during each trial, permitting statistical comparison of outcomes between large and small DTRs at 4 and 8 DPI. At 4 DPI, outcomes in bodies, legs, and saliva were not different between the 11 and 15 °C DTR trials. Body and saliva outcomes were also not statistically different between fluctuation treatments at 8 DPI; however, there was a significant difference in leg infections at 8 DPI (p = 0.02) with 39.3% of individuals with a positive body also showing dissemination to legs in trial 1 versus 15.2% of individuals in trial 2 (Figure 3A). There were no differences identified between trials in body titers at 4 DPI (p = 0.06) or 8 DPI (p = 0.42) or in leg titers at either timepoint (4 DPI p = 0.41, 8 DPI p = 0.29) (Figure 3B).
4. Discussion
We tested whether Cx. quinquefasciatus and Cx. tarsalis fed blood containing WNV would show variable vector competence when exposed to either static or fluctuating incubation temperatures. The results of this study suggest that while infection, dissemination, and transmission rates appear to be comparable between treatments, viral titers may be more susceptible to changes in temperature treatment. Interestingly, these results appear to be species specific. Culex quinquefasciatus had higher viral titers in static treatments overall while Cx. tarsalis generally had higher viral titers in fluctuating treatments.
It is unclear what led to these species-specific responses to different temperature treatments, although this phenomenon has been seen in similar studies conducted with Aedes aegypti and Aedes albopictus exposed to Chikungunya virus [21]. It is possible that the physiology of infection differs between these species and between exposure temperatures. While WNV specific tissue tropisms have been investigated in Cx. quinquefasciatus [30], less information is available for Cx. tarsalis to permit direct comparisons. In general, previous evaluations of these two species have shown higher vector competence of Cx. tarsalis populations for WNV than Cx. quinquefasciatus [31]. Higher temperatures have been shown to result in higher titer infections and shortened extrinsic incubation periods in Cx. tarsalis, although this relationship may be dependent on viral strain [32]. Overall, the periods of high temperature in the fluctuating treatment may have had a disproportionate effect on the viral titer of body tissues in this species to the minimum temperatures experienced [33]. Furthermore, this phenomenon could be related to differences in the geographic range of the two species. The range of Cx. tarsalis extends much further north than that of Cx. quinquefasciatus [27]. This broader range into more northerly areas may result in different physiological responses to temperature changes in this species.
For Cx. quinquefasciatus, there appeared to be very little difference in outcomes between the two DTR treatments except for dissemination rates. At 8 DPI, significantly more Cx. quinquefasciatus displayed a disseminated infection in the 11 °C treatment compared with the 15 °C DTR treatment. Considering the lack of significant differences between fluctuating and static treatments, this was a surprising outcome. There is some evidence that at high temperatures of 32 °C, female Cx. tarsalis show some capacity to modulate and prevent dissemination of western equine encephalomyelitis virus compared with those incubated at lower temperatures of 18 and 25 °C [34]. If this is also the case in Cx. quinquefasciatus with WNV, periods of high temperatures associated with the high DTR treatment may have reduced dissemination overall. Mosquitoes in the high DTR treatment were exposed to temperatures >32 °C for around 5 h compared to under 2 h in the low DTR treatment.
These results are increasingly relevant in light of ongoing discussions of climate change impacts on global disease transmission dynamics. In general, climate change in North America is anticipated to result in elevated temperatures, shorter winters, and increasing extreme weather events including heavy rainfall—all of which have the potential to impact vector-borne disease dynamics [35]. Additionally, DTR’s are anticipated to shift in response to climate change with some models indicating that minimum daily temperatures are likely to increase more than maximum daily temperatures on average, potentially leading to smaller DTRs [36,37]. This makes our finding of significantly higher dissemination in Cx. quinquefasciatus at the smaller DTR more concerning. This also further emphasizes the need for research on impacts of climate change on vector-borne diseases.
The present study included some challenges and limitations that must be acknowledged. The first limitation was the age of the two mosquito colonies used for these experiments. The colonies used are well characterized, stable laboratory populations that can provide good baseline data; however, younger colonies would have been useful to understand how populations acclimated to temperature fluctuations responded to our treatments. Attempts to establish young colonies for these experiments to run alongside colony specimens unfortunately failed. Additional studies are encouraged to investigate these patterns in young colonies or field populations that may be more adapted to temperature fluctuations. Additionally, the DTR regimens used in these trials did not follow typical sinusoidal temperature patterns due to software limitations in the temperature controllers. While the software allowed us to program smooth, constant temperature ramping, it was not a perfect reflection of natural temperature fluctuations. We also only tested fluctuations occurring around a single mean temperature. Studies conducted on other systems have found that the magnitude and direction of effects of fluctuating temperatures may depend on the mean temperature being studied [23,24]. Further studies on the impact of fluctuations around additional mean temperatures are warranted. Finally, both Cx. tarsalis and Cx. quinquefasciatus are considered crepuscular to nocturnal [38,39,40,41] and are likely to be in sheltered locations during periods of peak heat. The temperature regimens we used replicated those that would be anticipated in areas such as Houston, TX or Las Vegas, NV during WNV outbreaks and were modelled after temperature records from these locations. Most temperature records are taken from exposed areas rather than sheltered locations, so the high temperatures of our fluctuating treatments may have been hotter than these mosquitoes would have experienced naturally.
5. Conclusions
These experiments provide evidence that virus titers in the bodies, and in some cases the legs, of Cx. quinquefasciatus and Cx. tarsalis can be significantly different between fluctuating and static temperature conditions. For studies investigating virus titer dynamics in arthropod hosts, replicating natural temperature fluctuations may be advantageous to get the most accurate representation of viral titers over time. However, for studies investigating infection, dissemination, or transmission rates, our results indicated that temperature fluctuations did not meaningfully impact these values and average, static temperatures may be adequate. These results are likely species specific and may need to be investigated on a case-by-case basis.
Author Contributions
Conceptualization, B.L.M.; methodology, B.L.M. and J.L.K.; software, B.L.M.; validation, B.L.M. and J.L.K.; formal analysis, B.L.M.; investigation, B.L.M.; resources, J.L.K. and C.R.C.; data curation, B.L.M.; writing—original draft preparation, B.L.M.; writing—review and editing, J.L.K. and C.R.C.; visualization, B.L.M.; supervision, J.L.K. and C.R.C.; project administration, B.L.M.; funding acquisition, C.R.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available from the authors by request.
Acknowledgments
We thank Sean Masters and the CDC animal care staff for rearing the Culex used in this study and Jason Velez for preparing cells for viral infection and plaque assays. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the CDC or the U.S. Department of Agriculture. The conclusions in this report are those of the authors and do not necessarily represent the views of the USDA or the CDC. USDA is an equal opportunity provider and employer.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mordecai, E.A.; Caldwell, J.M.; Grossman, M.K.; Lippi, C.A.; Johnson, L.R.; Neira, M.; Rohr, J.R.; Ryan, S.J.; Savage, V.; Shocket, M.S.; et al. Thermal biology of mosquito-borne disease. Ecol. Lett. 2019, 22, 1690–1708. [Google Scholar] [CrossRef] [Green Version]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Current. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, T.; Greischar, M.A.; Wadhawan, K.; Gilbert, B.; Paaijmans, K.; Mideo, N. Temperature-dependent variation in the extrinsic incubation period elevates the risk of vector-borne disease emergence. Epidemics 2019, 30, 100382. [Google Scholar] [CrossRef]
- Shelton, R.M. The effect of temperatures on development of eight mosquito species. Mosq. News 1973, 33, 1–12. [Google Scholar]
- Rueda, L.M.; Patel, K.J.; Axtell, R.C.; Stinner, R.E. Temperature-dependent development and survival rates of Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 1990, 27, 892–898. [Google Scholar] [CrossRef]
- Mohammed, A.; Chadee, D.D. Effects of different temperature regimens on the development of Aedes aegypti (L.) (Diptera: Culicidae) mosquitoes. Acta Tropica 2011, 119, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Reiskind, M.H.; Zarrabi, A.A. Is bigger really bigger? Differential responses to temperature in measures of body size of the mosquito, Aedes albopictus. J. Insect Physiol. 2012, 58, 911–917. [Google Scholar] [CrossRef]
- Ciota, A.T.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The effect of temperature on life history traits of Culex mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef]
- Rund, S.S.; Hou, T.Y.; Ward, S.M.; Collins, F.H.; Duffield, G.E. Genome-wide profiling of diel and circadian gene expression in the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2011, 108, E421–E430. [Google Scholar] [CrossRef] [Green Version]
- Rivas, G.B.S.; Teles-de-Freitas, R.; Pavan, M.G.; Lima, J.B.P.; Peixoto, A.A.; Bruno, R.V. Effects of Light and Temperature on Daily Activity and Clock Gene Expression in Two Mosquito Disease Vectors. J. Biol. Rhythm. 2018, 33, 272–288. [Google Scholar] [CrossRef]
- Reeves, W.C.; Hardy, J.L.; Reisen, W.K.; Milby, M.M. Potential effect of global warming on mosquito-borne arboviruses. J. Med. Entomol. 1994, 31, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, E.D.; Thomas, M.B. Local adaptation to temperature and the implications for vector-borne diseases. Trends Parasitol. 2014, 30, 115–122. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Koenraadt, C.J.M.; Meiswinkel, R. Mosquitoes and Culicoides biting midges: Vector range and the influence of climate change. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, S.P.; Keeling, M.J. The impact of temperature changes on vector-borne disease transmission: Culicoides midges and bluetongue virus. J. R. Soc. Interface 2017, 14, 20160481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turell, M.J.; O’Guinn, M.L.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef]
- Anderson, S.L.; Richards, S.L.; Tabachnick, W.J.; Smartt, C.T. Effects of West Nile virus dose and extrinsic incubation temperature on temporal progression of vector competence in Culex pipiens quinquefasciatus. J. Am. Mosq. Control. Assoc. 2010, 26, 103–107. [Google Scholar] [CrossRef]
- Vogels, C.B.; Goertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef] [Green Version]
- Heitmann, A.; Jansen, S.; Luhken, R.; Helms, M.; Pluskota, B.; Becker, N.; Kuhn, C.; Schmidt-Chanasit, J.; Tannich, E. Experimental risk assessment for chikungunya virus transmission based on vector competence, distribution and temperature suitability in Europe, 2018. Eurosurveillance 2018, 23, 1800033. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Meyer, R.P.; Hardy, J.L.; Reisen, W.K. Diel changes in adult mosquito microhabitat temperatures and their relationship to the extrinsic incubation of arboviruses in mosquitoes in Kern County, California. J. Med. Entomol. 1990, 27, 607–614. [Google Scholar] [CrossRef]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Ortiz, S.; Zirbel, K.; Lounibos, L.P. Diurnal Temperature Range and Chikungunya Virus Infection in Invasive Mosquito Vectors. J. Med. Entomol. 2018, 55, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Paaijmans, K.P.; Fansiri, T.; Carrington, L.B.; Kramer, L.D.; Thomas, M.B.; Scott, T.W. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 7460–7465. [Google Scholar] [CrossRef] [Green Version]
- Carrington, L.B.; Seifert, S.N.; Armijos, M.V.; Lambrechts, L.; Scott, T.W. Reduction of Aedes aegypti vector competence for dengue virus under large temperature fluctuations. Am. J. Trop. Med. Hyg. 2013, 88, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paaijmans, K.P.; Blanford, S.; Bell, A.S.; Blanford, J.I.; Read, A.F.; Thomas, M.B. Influence of climate on malaria transmission depends on daily temperature variation. Proc. Natl. Acad. Sci. USA 2010, 107, 15135–15139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. West Nile Virus-Final Cumulative Maps & Data for 1999–2019. Available online: https://www.cdc.gov/westnile/statsmaps/cumMapsData.html (accessed on 23 March 2021).
- U.S. Climate Data. Available online: https://www.usclimatedata.com/climate/united-states/us (accessed on 18 October 2019).
- Shocket, M.S.; Verwillow, A.B.; Numazu, M.G.; Slamani, H.; Cohen, J.M.; El Moustaid, F.; Rohr, J.; Johnson, L.R.; Mordecai, E.A. Transmission of West Nile and five other temperate mosquito-borne viruses peaks at temperatures between 23 degrees C and 26 degrees C. Elife 2020, 9, e58511. [Google Scholar] [CrossRef]
- Anderson, S.L.; Richards, S.L.; Smartt, C.T. A simple method for determining arbovirus transmission in mosquitoes. J. Am. Mosq. Control. Assoc. 2010, 26, 108–111. [Google Scholar] [CrossRef] [Green Version]
- McGregor, B.L.; Connelly, C.R.; Kenney, J.L. Infection, Dissemination, and Transmission Potential of North American Culex quinquefasciatus, Culex tarsalis, and Culicoides sonorensis for Oropouche Virus. Viruses 2021, 13, 226. [Google Scholar] [CrossRef]
- Girard, Y.A.; Klingler, K.A.; Higgs, S. West Nile virus dissemination and tissue tropisms in orally infected Culex pipiens quinquefasciatus. Vector-Borne Zoonotic Dis. 2004, 4, 109–122. [Google Scholar] [CrossRef]
- Goddard, L.B.; Roth, A.E.; Reisen, W.K.; Scott, T.W. Vector competence of California mosquitoes for West Nile virus. Emerg. Infect. Dis. 2002, 8, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Fang, Y.; Martinez, V.M. Effects of temperature on the transmission of West Nile virus by Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 2014, 43, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Cornel, A.J.; Jupp, P.G.; Blackburn, N.K. Environmental temperature on the vector competence of Culex univittatus (Diptera: Culicidae) for West Nile Virus. J. Med. Entomol. 1993, 30, 449–456. [Google Scholar] [CrossRef]
- Kramer, L.D.; Hardy, J.L.; Presser, S.B. Effect of temperature of extrinsic incubation on the vector competence of Culex tarsalis for Western equine encephalomyelitis virus. Am. J. Trop. Med. Hyg. 1983, 32, 1130–1139. [Google Scholar] [CrossRef]
- Paz, S. Climate change impacts on West Nile virus transmission in a global context. Philos. Trans. R. Soc. B 2015, 370, 20130561. [Google Scholar] [CrossRef]
- Braganza, K.; Karoly, D.J.; Arblaster, J.M. Diurnal temperature range as an index of global climate change during the twentieth century. Geophys. Res. Lett. 2004, 31, 1–4. [Google Scholar] [CrossRef]
- Lobell, D.B.; Bonfils, C.; Duffy, P.B. Climate change uncertainty for daily minimum and maximum temperatures: A model inter-comparison. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef] [Green Version]
- Yee, W.L.; Foster, W.A. Diel sugar-feeding and host-seeking rhythms in mosquitoes (Diptera: Culicidae) under laboratory conditions. J. Med. Entomol. 1992, 29, 784–791. [Google Scholar] [CrossRef]
- Reisen, W.K.; Lothrop, H.D.; Meyer, R.P. Time of host-seeking by Culex tarsalis (Diptera: Culicidae) in California. J. Med. Entomol. 1997, 34, 430–437. [Google Scholar] [CrossRef]
- Godsey, M.S.G., Jr.; Burkhalter, K.; Delorey, M.; Savage, H.M. Seasonality and time of host-seeking activity of Culex tarsalis and floodwater Aedes in northern Colorado, 2006–2007. J. Am. Mosq. Control. Assoc. 2010, 26, 148–159. [Google Scholar]
- Savage, H.M.; Anderson, M.; Gordon, E.; McMillen, L.; Colton, L.; Delorey, M.; Sutherland, G.; Aspen, S.; Charnetzky, D.; Burkhalter, K.; et al. Host-seeking heights, host-seeking activity patterns, and West Nile virus infection rates for members of the Culex pipiens complex at different habitat types within the Hybrid Zone, Shelby County, TN, 2002 (Diptera: Culicidae). J. Med. Entomol. 2008, 45, 276–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Temperature readouts of Hobo monitors placed inside incubators held at a static temperature of 28 °C, trial 1 11 °C DTR between 22.5 and 33.5 °C, and trial 2 15 °C DTR between 20.5 and 35.5 °C.
Figure 1.
Temperature readouts of Hobo monitors placed inside incubators held at a static temperature of 28 °C, trial 1 11 °C DTR between 22.5 and 33.5 °C, and trial 2 15 °C DTR between 20.5 and 35.5 °C.

Figure 2.
Mean viral titer in bodies, legs, and saliva of Cx. quinquefasciatus and Cx. tarsalis exposed to fluctuating and static temperature treatments at 11 °C DTR (Trial 1) and 15 °C DTR (Trial 2). Overall, Cx. quinquefasciatus had higher titers in static treatments in both trials while Cx. tarsalis had higher mean titers in fluctuating treatments. Asterisks denote significance at p = 0.05 (*) or p < 0.001 (**).
Figure 2.
Mean viral titer in bodies, legs, and saliva of Cx. quinquefasciatus and Cx. tarsalis exposed to fluctuating and static temperature treatments at 11 °C DTR (Trial 1) and 15 °C DTR (Trial 2). Overall, Cx. quinquefasciatus had higher titers in static treatments in both trials while Cx. tarsalis had higher mean titers in fluctuating treatments. Asterisks denote significance at p = 0.05 (*) or p < 0.001 (**).

Figure 3.
Comparison of Culex quinquefasciatus outcomes when exposed to DTRs of 11 °C (Trial 1) and 15 °C (Trial 2). (A) Overall percent positive bodies (infection), legs (dissemination), and saliva (transmission potential); Significance at p = 0.05 is denoted with *. (B) Box plots showing comparative body and leg titers at 4 DPI (on left) and 8 DPI (on right).
Figure 3.
Comparison of Culex quinquefasciatus outcomes when exposed to DTRs of 11 °C (Trial 1) and 15 °C (Trial 2). (A) Overall percent positive bodies (infection), legs (dissemination), and saliva (transmission potential); Significance at p = 0.05 is denoted with *. (B) Box plots showing comparative body and leg titers at 4 DPI (on left) and 8 DPI (on right).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Positive bodies (infection), legs (dissemination), and saliva (transmission potential) for Culex quinquefasciatus and Culex tarsalis exposed to static or fluctuating incubation treatments. No significant differences were observed in infection, dissemination, or transmission rates between temperature treatments for either species or trial.
Table 1.
Positive bodies (infection), legs (dissemination), and saliva (transmission potential) for Culex quinquefasciatus and Culex tarsalis exposed to static or fluctuating incubation treatments. No significant differences were observed in infection, dissemination, or transmission rates between temperature treatments for either species or trial.
| Culex Species | Trial | Time (DPI) | Treatment | Bodies † | Bodies p | Legs † | Legs p | Saliva † | Saliva p |
|---|---|---|---|---|---|---|---|---|---|
| quinquefasciatus | 1 | 4 | Static | 29/31 | 0.255 | 2/29 | 1 | 1/2 | 1 |
| Fluctuating | 25/31 | 2/25 | 2/2 | ||||||
| 8 | Static | 28/32 | 1 | 9/28 | 0.781 | 1/9 | 0.450 | ||
| Fluctuating | 28/31 | 11/28 | 0/11 | ||||||
| 2 | 4 | Static | 69/89 | 0.319 | 4/69 | 1 | 0/4 | 1 | |
| Fluctuating | 63/90 | 3/63 | 0/3 | ||||||
| 8 | Static | 62/90 | 0.622 | 10/62 | 0.622 | 2/10 | 1 | ||
| Fluctuating | 66/90 | 10/66 | 1/10 | ||||||
| 12 | Static | 76/113 | 0.241 | 39/76 | 1 | 10/39 | 0.219 | ||
| Fluctuating | 51/87 | 25/51 | 2/25 | ||||||
| tarsalis | 1 | 4 | Static | 19/23 | 0.109 | 11/19 | 0.213 | 4/11 | 0.338 |
| Fluctuating | 23/23 | 8/23 | 1/8 | ||||||
| 8 | Static | 24/25 | 1 | 22/24 | 0.489 | 20/22 | 0.223 | ||
| Fluctuating | 24/26 | 24/24 | 24/24 | ||||||
| 2 | 4 | Static | 42/63 | 0.714 | 7/42 | 0.757 | 1/7 | 1 | |
| Fluctuating | 41/65 | 5/41 | 1/5 | ||||||
| 8 | Static | 19/41 | 0.691 | 15/19 | 0.456 | 10/15 | 0.716 | ||
| Fluctuating | 28/68 | 25/28 | 19/25 |
† Bodies: positive bodies/total bodies tested; Legs: positive legs/positive bodies; Saliva: positive saliva/positive bodies.
Table 2.
Average viral titer present in bodies, legs, and saliva of Culex quinquefasciatus and Culex tarsalis in trial 1 (11 °C DTR) and trial 2 (15 °C DTR). All viral titers are expressed in log10 PFU/mL.
Table 2.
Average viral titer present in bodies, legs, and saliva of Culex quinquefasciatus and Culex tarsalis in trial 1 (11 °C DTR) and trial 2 (15 °C DTR). All viral titers are expressed in log10 PFU/mL.
| Culex Species | Trial | Time (DPI) | Treatment | Body Titer | Body p | Leg Titer | Leg p | Saliva Titer | Saliva p |
|---|---|---|---|---|---|---|---|---|---|
| quinquefasciatus | 1 | 4 | Static | 5.24 | 0.574 | 1.48 | 0.333 | 0.82 | n/a |
| Fluctuating | 5.20 | 0.92 | 0.82 | ||||||
| 8 | Static | 6.18 | 0.054 | 4.17 | 0.619 | 4.69 | n/a | ||
| Fluctuating | 5.82 | 3.90 | n/a | ||||||
| 2 | 4 | Static | 6.49 | 0.421 | 1.18 | 0.270 | n/a | n/a | |
| Fluctuating | 5.55 | 0.82 | n/a | ||||||
| 8 | Static | 7.02 | <0.001 | 4.20 | 0.791 | 2.15 | 0.667 | ||
| Fluctuating | 6.39 | 4.17 | 1.00 | ||||||
| 12 | Static | 7.62 | 0.007 | 4.60 | 0.821 | 2.49 | 0.075 | ||
| Fluctuating | 6.92 | 4.18 | 1.00 | ||||||
| tarsalis | 1 | 4 | Static | 6.71 | 0.062 | 3.47 | 0.342 | 1.00 | n/a |
| Fluctuating | 6.84 | 3.19 | 0.82 | ||||||
| 8 | Static | 7.81 | <0.001 | 5.01 | 0.008 | 3.67 | 0.191 | ||
| Fluctuating | 8.15 | 5.44 | 4.11 | ||||||
| 2 | 4 | Static | 5.46 | 0.068 | 4.25 | 0.222 | 1.70 | n/a | |
| Fluctuating | 6.97 | 4.66 | 2.99 | ||||||
| 8 | Static | 7.95 | 0.025 | 5.07 | 0.665 | 3.18 | 0.854 | ||
| Fluctuating | 8.23 | 5.06 | 4.79 |
Significant p-values are shown in bold.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
McGregor, B.L.; Kenney, J.L.; Connelly, C.R. The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes. Viruses 2021, 13, 1822. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091822
AMA Style
McGregor BL, Kenney JL, Connelly CR. The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes. Viruses. 2021; 13(9):1822. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091822
Chicago/Turabian StyleMcGregor, Bethany L., Joan L. Kenney, and C. Roxanne Connelly. 2021. "The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes" Viruses 13, no. 9: 1822. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091822
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.