
The Pathobiology of H7N3 Low and High Pathogenicity Avian Influenza Viruses from the United States Outbreak in 2020 Differs between Turkeys and Chickens


 ,
,
Abstract
: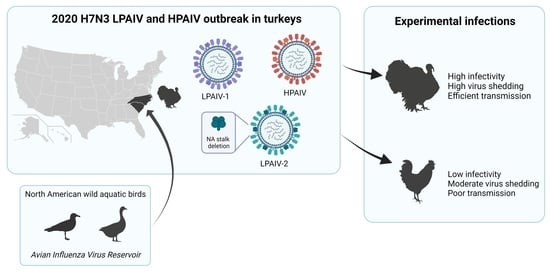
1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Animals and Housing
2.3. Experimental Design
2.4. Viral Titration in Swabs and Tissues
2.5. Statistical Analysis
2.6. Sequence Analyses
3. Results
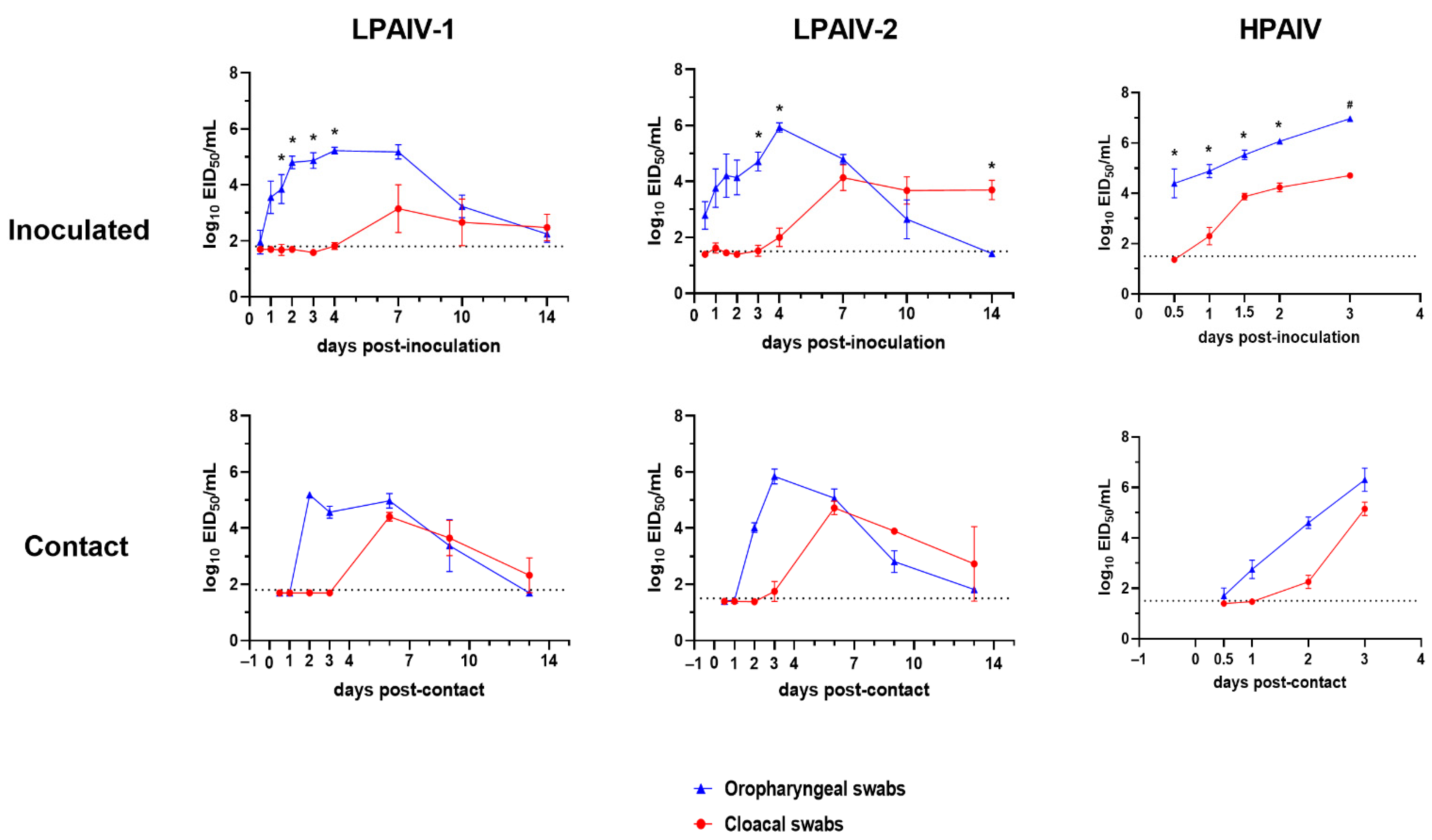
3.1. Infectivity, Transmission, and Pathogenicity of the H7N3 LPAIVs in Turkeys and Chickens
3.2. Infectivity, Transmission, and Pathogenicity of the H7N3 HPAIV in Turkeys and Chickens
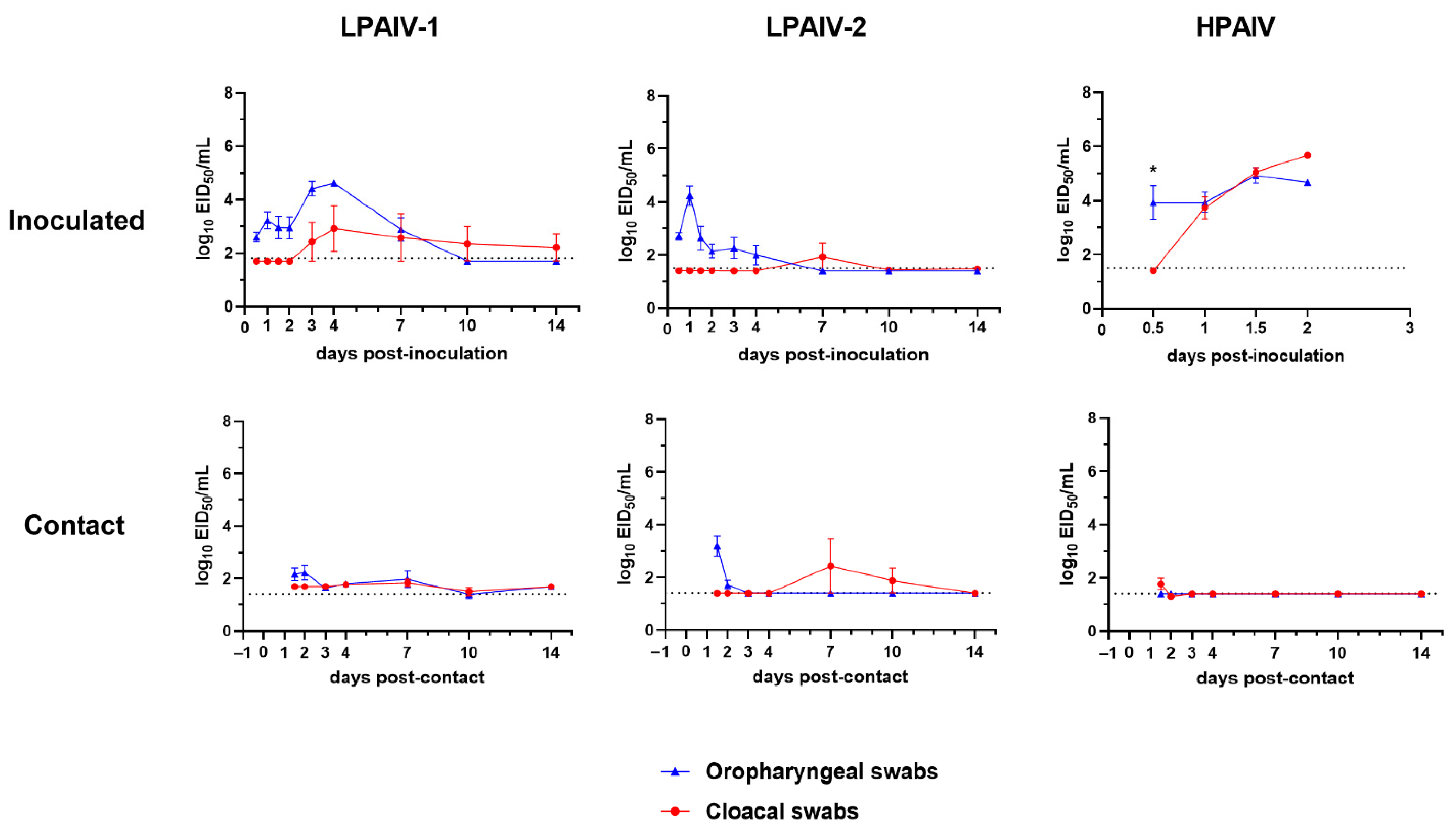
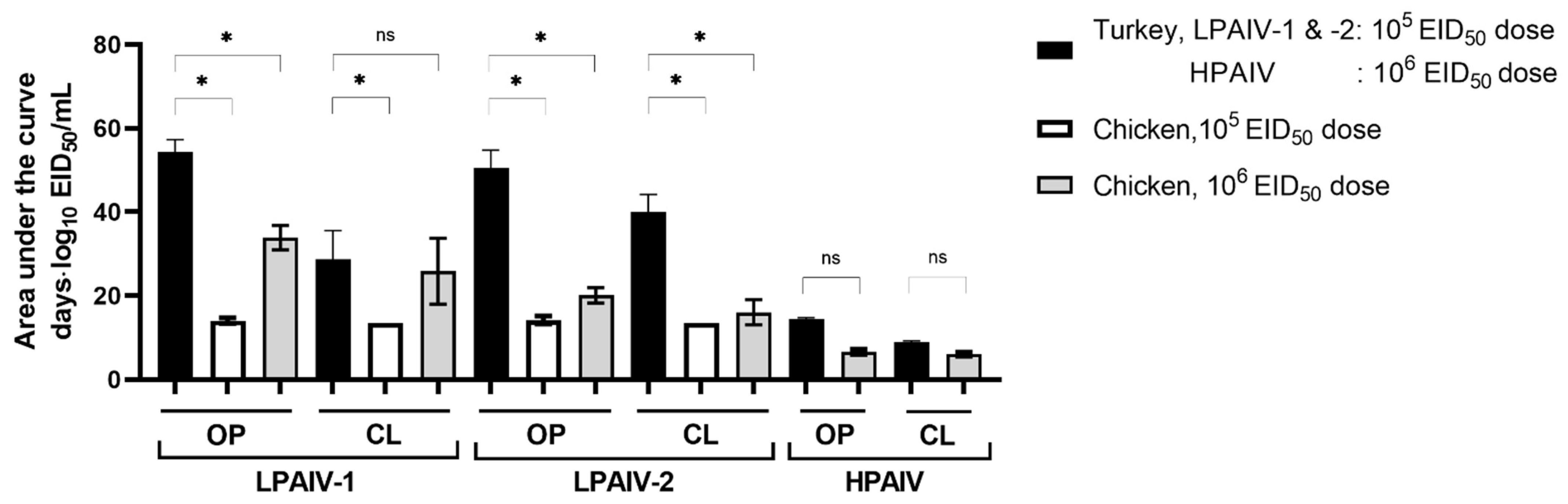
3.3. Viral Shedding and Virus Detection in Tissues

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bird Species | Virus | Dose (log10 EID50) | Inoculated | Contact Exposed | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Birds Shedding Virus/Total | No. of Dead Birds/Total (MDT) 1 | No. of Birds HI Positive/Total (Mean HI Titer) 2 | No. of Birds Infected/Total 3 | BID50 (log10) 4 | No. of Birds Shedding Virus/Total | No. of Dead Birds/Total (MDT) | No. of Birds HI Positive/Total (Mean HI Titer) 2 | No. of Birds Infected/Total 3 | |||
| Turkeys | LPAIV-1 | 2 | 0/4 | 0/4 | 0/4 (<3.0) | 0/4 | 2.5 | na | na | na | na |
| 3 | 5/5 | 0/5 | 5/5 (9.4) | 5/5 | 2/2 | 0/2 | 2/2 (10) | 2/2 | |||
| 5 | 5/5 | 0/5 | 5/5 (9.4) | 5/5 | 2/2 | 0/2 | 2/2 (10) | 2/2 | |||
| LPAIV-2 | 2 | 5/5 | 0/5 | 5/5 (8.0) | 5/5 | <2 | na | na | na | na | |
| 3 | 5/5 | 1/5 5 | 4/4 (6.5) | 5/5 | 2/2 | 1/2 5 | 1/1 (8.0) | 2/2 | |||
| 5 | 5/5 | 0/5 | 5/5 (7.6) | 5/5 | 2/2 | 0/2 | 2/2 (6.0) | 2/2 | |||
| HPAIV | 2 | 5/5 | 5/5 (2.4) | na | 5/5 | <2 | na | na | na | na | |
| 4 | 5/5 | 5/5 (2.2) | na | 5/5 | 2/2 | 2/2 (3) | na | 2/2 | |||
| 6 | 5/5 | 5/5 (2.0) | na | 5/5 | 2/2 | 2/2 (3) | na | 2/2 | |||
| Chickens | LPAIV-1 | 2 | 0/5 | 0/5 | 0/5 (<3.0) | 0/5 | 5.7 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 |
| 3 | 0/5 | 0/5 | 0/5 (<3.0) | 0/5 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |||
| 5 | 1/8 | 0/8 | 0/5 6,7 (<3.0) | 1/8 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |||
| 6 | 8/8 | 0/8 | 5/5 6 (6.0) | 8/8 | 3/3 | 0/3 | 1/3 (5.0) | 3/3 | |||
| LPAIV-2 | 2 | 0/5 | 0/5 | 0/5 (<3.0) | 0/5 | 5.4 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |
| 3 | 1/5 | 0/5 | 1/5 (3.0) | 1/5 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |||
| 5 | 1/8 | 0/8 | 0/5 6,7 (3.0) | 1/8 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |||
| 6 | 8/8 | 0/8 | 5/5 6 (4.5) | 8/8 | 3/3 | 0/3 | 1/3 (7.0) | 3/3 | |||
| HPAIV | 2 | 1/5 | 1/5 (1) | 0/5 (<3.0) | 1/5 | 3.7 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |
| 4 | 5/5 | 5/5 (1.6) | na | 5/5 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |||
| 6 | 8/8 | 8/8 (1.6) | na | 8/8 | 0/3 | 0/3 | 0/3 (<3.0) | 0/3 | |||
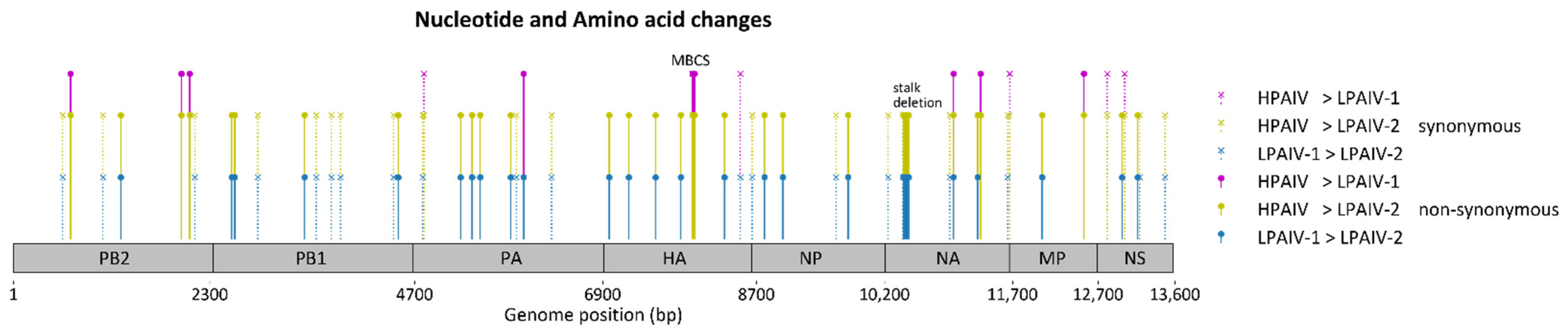
3.4. Sequence Comparisons of the H7N3 Viruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Avian Influenza (Including Infection with High Pathogenicity Avian Influenza Viruses). Manual for Diagnostic Tests and Vaccines for Terrestrial Animals, 2021 edition; World Health Organization (OIE), Posted Online 30 June 2021; Available online: https://www.oie.int/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 6 May 2021).
- Swayne, D.E.; Suarez, D.L.; Sims, L.D. Influenza. In Diseases of Poultry; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 210–256. [Google Scholar]
- Pantin-Jackwood, M.J.; Swayne, D.E. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev. Sci. Tech. 2009, 28, 113–136. [Google Scholar] [CrossRef]
- Lee, D.-H.; Criado, M.F.; Swayne, D.E. Pathobiological Origins and Evolutionary History of Highly Pathogenic Avian Influenza Viruses. Cold Spring Harb. Perspect. Med. 2021, 11, a038679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, H.; Moreira, R.; Avalos, P.; Capua, I.; Marangon, S. Avian influenza in poultry in Chile. Vet. Rec. 2002, 151, 188. [Google Scholar]
- Hirst, M.; Astell, C.R.; Griffith, M.; Coughlin, S.M.; Moksa, M.; Zeng, T.; Smailus, D.E.; Holt, R.; Jones, S.; Marra, M.A.; et al. Novel Avian Influenza H7N3 Strain Outbreak, British Columbia. Emerg. Infect. Dis. 2004, 10, 2192–2195. [Google Scholar] [CrossRef] [PubMed]
- Berhane, Y.; Hisanaga, T.; Kehler, H.; Neufeld, J.; Manning, L.; Argue, C.; Handel, K.; Hooper-McGrevy, K.; Jonas, M.; Robinson, J.; et al. Highly Pathogenic Avian Influenza Virus A (H7N3) in Domestic Poultry, Saskatchewan, Canada, 2007. Emerg. Infect. Dis. 2009, 15, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Maurer-Stroh, S.; Lee, R.T.C.; Gunalan, V.; Eisenhaber, F. The highly pathogenic H7N3 avian influenza strain from July 2012 in Mexico acquired an extended cleavage site through recombination with host 28S rRNA. Virol. J. 2013, 10, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapczynski, D.R.; Pantin-Jackwood, M.; Guzman, S.G.; Ricardez, Y.; Spackman, E.; Bertran, K.; Suarez, D.L.; Swayne, D. Characterization of the 2012 Highly Pathogenic Avian Influenza H7N3 Virus Isolated from Poultry in an Outbreak in Mexico: Pathobiology and Vaccine Protection. J. Virol. 2013, 87, 9086–9096. [Google Scholar] [CrossRef] [Green Version]
- Youk, S.; Lee, D.-H.; Ferreira, H.L.; Afonso, C.L.; Absalon, A.E.; Swayne, D.E.; Suarez, D.L.; Pantin-Jackwood, M.J. Rapid evolution of Mexican H7N3 highly pathogenic avian influenza viruses in poultry. PLoS ONE 2019, 14, e0222457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killian, M.L.; Kim-Torchetti, M.; Hines, N.; Yingst, S.; DeLiberto, T.; Lee, D.-H. Outbreak of H7N8 Low Pathogenic Avian Influenza in Commercial Turkeys with Spontaneous Mutation to Highly Pathogenic Avian Influenza. Genome Announc. 2016, 4, e00457-16. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Torchetti, M.K.; Killian, M.L.; Berhane, Y.; Swayne, D.E. Highly Pathogenic Avian Influenza A(H7N9) Virus, Tennessee, USA, March 2017. Emerg. Infect. Dis. 2017, 23, 1860–1863. [Google Scholar] [CrossRef]
- Youk, S.; Lee, D.-H.; Killian, M.L.; Pantin-Jackwood, M.J.; Swayne, D.E.; Torchetti, M.K. Highly Pathogenic Avian Influenza A(H7N3) Virus in Poultry, United States, 2020. Emerg. Infect. Dis. 2020, 26, 2966–2969. [Google Scholar] [CrossRef]
- Li, J.; Zu Dohna, H.; Cardona, C.J.; Miller, J.; Carpenter, T.E. Emergence and Genetic Variation of Neuraminidase Stalk Deletions in Avian Influenza Viruses. PLoS ONE 2011, 6, e14722. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E.; Gelb, J.; Preskenis, L.A.; Ladman, B.S.; Pope, C.R.; Pantin-Jackwood, M.J.; McKinley, E.T. The pathogenesis of low pathogenicity H7 avian influenza viruses in chickens, ducks and turkeys. Virol. J. 2010, 7, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, T.W.; Munier, S.; Larcher, T.; Soubieux, D.; Ledevin, M.; Esnault, E.; Tourdes, A.; Croville, G.; Guerin, J.-L.; Quere, P.; et al. Length Variations in the NA Stalk of an H7N1 Influenza Virus Have Opposite Effects on Viral Excretion in Chickens and Ducks. J. Virol. 2011, 86, 584–588. [Google Scholar] [CrossRef] [Green Version]
- Munier, S.; Larcher, T.; Cormier-Aline, F.; Soubieux, D.; Su, B.; Guigand, L.; Labrosse, B.; Cherel, Y.; Quéré, P.; Marc, D.; et al. A Genetically Engineered Waterfowl Influenza Virus with a Deletion in the Stalk of the Neuraminidase Has Increased Virulence for Chickens. J. Virol. 2010, 84, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Swayne, D.; Senne, D.; Beard, C. Influenza; American Association of Avian Pathologists: Kennett Square, PA, USA, 1998. [Google Scholar]
- Chrzastek, K.; Lee, D.-H.; Smith, D.; Sharma, P.; Suarez, D.; Pantin-Jackwood, M.J.; Kapczynski, D.R. Use of Sequence-Independent, Single-Primer-Amplification (SISPA) for rapid detection, identification, and characterization of avian RNA viruses. Virology 2017, 509, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Horimoto, T.; Kawaoka, Y. Molecular Changes in virulent mutants arising from avirulent avian influenza viruses during Replication in 14-day-old embryonated eggs. Virology 1995, 206, 755–759. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Lee, Y.J.; Swayne, D.; Senne, D.; Linares, D.J.; Suarez, D. Assessing potential pathogenicity of avian influenza virus: Current and experimental system. Avian Dis. 2007, 51, 260–263. [Google Scholar] [CrossRef]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. Available online: https://www.arxiv-vanity.com/papers/1303.3997/ (accessed on 24 June 2018).
- Dimitrov, K.M.; Sharma, P.; Volkening, J.D.; Goraichuk, I.V.; Wajid, A.; Rehmani, S.F.; Basharat, A.; Shittu, I.; Joannis, T.M.; Miller, P.J.; et al. A robust and cost-effective approach to sequence and analyze complete genomes of small RNA viruses. Virol. J. 2017, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Wilm, A.; Aw, P.P.K.; Bertrand, D.; Yeo, G.H.T.; Ong, S.H.; Wong, C.H.; Khor, C.C.; Petric, R.; Hibberd, M.L.; Nagarajan, N. LoFreq: A sequence-quality aware, ultra-sensitive variant caller for uncovering cell-population heterogeneity from high-throughput sequencing datasets. Nucleic Acids Res. 2012, 40, 11189–11201. [Google Scholar] [CrossRef] [Green Version]
- Swayne, D.E.; Slemons, R.D. Using Mean Infectious Dose of High- and Low-Pathogenicity Avian Influenza Viruses Originating from Wild Duck and Poultry as One Measure of Infectivity and Adaptation to Poultry. Avian Dis. 2008, 52, 455–460. [Google Scholar] [CrossRef]
- Youk, S.-S.; Lee, D.-H.; Leyson, C.M.; Smith, D.; Criado, M.F.; DeJesus, E.; Swayne, D.; Pantin-Jackwood, M.J. Loss of Fitness of Mexican H7N3 Highly Pathogenic Avian Influenza Virus in Mallards after Circulating in Chickens. J. Virol. 2019, 93, e00543-19. [Google Scholar] [CrossRef] [Green Version]
- Spackman, E.; Prosser, D.J.; Pantin-Jackwood, M.J.; Berlin, A.M.; Stephens, C.B. The Pathogenesis of Clade 2.3.4.4 H5 Highly Pathogenic Avian Influenza Viruses in Ruddy Duck (Oxyura Jamaicensis) and Lesser Scaup (Aythya affinis). J. Wildl. Dis. 2017, 53, 832–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J.C. Hemagglutination-Inhibition Assay for Influenza VIrus Subtype Identification and the Detection and Quantification of Serum Antibodies to Influenza Virus. In Animal Influenza Virus; Spackman, E., Ed.; Humana Press: Totova, NJ, USA, 2014. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Lee, C.-W.; Suarez, D.L. Application of real-time RT-PCR for the quantitation and competitive replication study of H5 and H7 subtype avian influenza virus. J. Virol. Methods 2004, 119, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bertran, K.; Lee, D.-H.; Pantin-Jackwood, M.J.; Spackman, E.; Balzli, C.; Suarez, D.L.; Swayne, D.E. Pathobiology of Clade 2.3.4.4 H5Nx High-Pathogenicity Avian Influenza Virus Infections in Minor Gallinaceous Poultry Supports Early Backyard Flock Introductions in the Western United States in 2014–2015. J. Virol. 2017, 91, e00960-17. [Google Scholar] [CrossRef] [Green Version]
- Banks, J.; Speidel, E.S.; Moore, E.; Plowright, L.; Piccirillo, A.; Capua, I.; Cordioli, P.; Fioretti, A.; Alexander, D.J. Changes in the haemagglutinin and the neuraminidase genes prior to the emergence of highly pathogenic H7N1 avian influenza viruses in Italy. Arch. Virol. 2001, 146, 963–973. [Google Scholar] [CrossRef]
- Campitelli, L.; Mogavero, E.; De Marco, M.A.; Delogu, M.; Puzelli, S.; Frezza, F.; Facchini, M.; Chiapponi, C.; Foni, E.; Cordioli, P.; et al. Interspecies transmission of an H7N3 influenza virus from wild birds to intensively reared domestic poultry in Italy. Virology 2004, 323, 24–36. [Google Scholar] [CrossRef]
- Durrant, J.D.; Bush, R.M.; Amaro, R.E. Microsecond Molecular Dynamics Simulations of Influenza Neuraminidase Suggest a Mechanism for the Increased Virulence of Stalk-Deletion Mutants. J. Phys. Chem. B 2016, 120, 8590–8599. [Google Scholar] [CrossRef]
- Giannecchini, S.; Clausi, V.; Di Trani, L.; Falcone, E.; Terregino, C.; Toffan, A.; Cilloni, F.; Matrosovich, M.; Gambaryan, A.S.; Bovin, N.V.; et al. Molecular adaptation of an H7N3 wild duck influenza virus following experimental multiple passages in quail and turkey. Virology 2010, 408, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Dlugolenski, D.; Jones, L.; Saavedra, G.; Tompkins, S.M.; Tripp, R.A.; Mundt, E. Passage of low-pathogenic avian influenza (LPAI) viruses mediates rapid genetic adaptation of a wild-bird isolate in poultry. Arch. Virol. 2011, 156, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Quan, K.; Wang, D.; Du, Y.; Qin, T.; Peng, D.; Liu, X. Truncation or Deglycosylation of the Neuraminidase Stalk Enhances the Pathogenicity of the H5N1 Subtype Avian Influenza Virus in Mallard Ducks. Front. Microbiol. 2020, 11, 583588. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Miller, P.J.; Spackman, E.; Swayne, D.E.; Susta, L.; Costa-Hurtado, M.; Suarez, D.L. Role of Poultry in the Spread of Novel H7N9 Influenza Virus in China. J. Virol. 2014, 88, 5381–5390. [Google Scholar] [CrossRef] [Green Version]
- Spackman, E.; Pantin-Jackwood, M.; Swayne, D.E.; Suarez, D.L.; Kapczynski, D.R. Impact of route of exposure and challenge dose on the pathogenesis of H7N9 low pathogenicity avian influenza virus in chickens. Virology 2015, 477, 72–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, L.E.L.; Swayne, D. Comparative Susceptibility of Selected Avian and Mammalian Species to a Hong Kong–Origin H5N1 High-Pathogenicity Avian Influenza Virus. Avian Dis. 2003, 47, 956–967. [Google Scholar] [CrossRef]
- Bertran, K.; Lee, D.-H.; Criado, M.F.; Smith, D.; Swayne, D.E.; Pantin-Jackwood, M.J. Pathobiology of Tennessee 2017 H7N9 low and high pathogenicity avian influenza viruses in commercial broiler breeders and specific pathogen free layer chickens. Vet. Res. 2018, 49, 82. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Costa-Hurtado, M.; Bertran, K.; DeJesus, E.; Smith, D.; Swayne, D.E. Infectivity, transmission and pathogenicity of H5 highly pathogenic avian influenza clade 2.3.4.4 (H5N8 and H5N2) United States index viruses in Pekin ducks and Chinese geese. Vet. Res. 2017, 48, 33. [Google Scholar] [CrossRef] [Green Version]
- Bertran, K.; Pantin-Jackwood, M.J.; Criado, M.F.; Lee, D.-H.; Balzli, C.L.; Spackman, E.; Suarez, D.L.; Swayne, D.E. Pathobiology and innate immune responses of gallinaceous poultry to clade 2.3.4.4A H5Nx highly pathogenic avian influenza virus infection. Vet. Res. 2019, 50, 89. [Google Scholar] [CrossRef] [Green Version]
- Bertran, K.; Swayne, D.E.; Pantin-Jackwood, M.J.; Kapczynski, D.R.; Spackman, E.; Suarez, D.L. Lack of chicken adaptation of newly emergent Eurasian H5N8 and reassortant H5N2 high pathogenicity avian influenza viruses in the U.S. is consistent with restricted poultry outbreaks in the Pacific flyway during 2014–2015. Virology 2016, 494, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Leyson, C.; Youk, S.; Smith, D.; Dimitrov, K.; Lee, D.-H.; Larsen, L.E.; Swayne, D.E.; Pantin-Jackwood, M.J. Pathogenicity and genomic changes of a 2016 European H5N8 highly pathogenic avian influenza virus (clade 2.3.4.4) in experimentally infected mallards and chickens. Virology 2019, 537, 172–185. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Costa-Hurtado, M.; Shepherd, E.; DeJesus, E.; Smith, D.; Spackman, E.; Kapczynski, D.R.; Suarez, D.L.; Stallknecht, D.E.; Swayne, D. Pathogenicity and Transmission of H5 and H7 Highly Pathogenic Avian Influenza Viruses in Mallards. J. Virol. 2016, 90, 9967–9982. [Google Scholar] [CrossRef] [Green Version]
- Spackman, E.; Prosser, D.J.; Pantin-Jackwood, M.; Stephens, C.B.; Berlin, A.M. Clade 2.3.4.4 H5 North American Highly Pathogenic Avian Influenza Viruses Infect, but Do Not Cause Clinical Signs in, American Black Ducks (Anas rubripes). Avian Dis. 2019, 63, 366–370. [Google Scholar] [CrossRef]
- Stephens, C.B.; Prosser, D.J.; Pantin-Jackwood, M.J.; Berlin, A.M.; Spackman, E. The Pathogenesis of H7 Highly Pathogenic Avian Influenza Viruses in Lesser Scaup (Aythya affinis). Avian Dis. 2019, 63, 230–234. [Google Scholar] [CrossRef]
- Luczo, J.M.; Prosser, D.J.; Pantin-Jackwood, M.J.; Berlin, A.M.; Spackman, E. The pathogenesis of a North American H5N2 clade 2.3.4.4 group A highly pathogenic avian influenza virus in surf scoters (Melanitta perspicillata). BMC Vet. Res. 2020, 16, 351. [Google Scholar] [CrossRef]
- Slemons, R.D.; Johnson, D.C.; Osborn, J.S.; Hayes, F. Type-A Influenza Viruses Isolated from Wild Free-Flying Ducks in California. Avian Dis. 1974, 18, 119–124. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Brown, J.D. Ecology of Avian Influenza in Wild Birds. In Avian Influenza; Wiley: Hoboken, NJ, USA, 2009; pp. 43–58. [Google Scholar]
- Halvorson, D.A.; Kelleher, C.J.; Senne, D.A. Epizootiology of avian influenza: Effect of season on incidence in sentinel ducks and domestic turkeys in Minnesota. Appl. Environ. Microbiol. 1985, 49, 914–919. [Google Scholar] [CrossRef] [Green Version]
- Morales, A.C., Jr.; Hilt, D.A.; Williams, S.M.; Pantin-Jackwood, M.J.; Suarez, D.L.; Spackman, E.; Stallknecht, D.E.; Jackwood, M.W. Biologic characterization of H4, H6, and H9 type low pathogenicity avian influenza viruses from wild birds in chickens and turkeys. Avian Dis. 2009, 53, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.P.S.; Pantin-Jackwood, M.; Suarez, D.; Saif, Y.M.; Lee, C.-W. Pathobiological characterization of low-pathogenicity H5 avian influenza viruses of diverse origins in chickens, ducks and turkeys. Arch. Virol. 2010, 155, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.P.; Pantin-Jackwood, M.; Yassine, H.M.; Saif, Y.M.; Lee, C.W. The high susceptibility of turkeys to influenza viruses of different origins implies their importance as potential intermediate hosts. Avian Dis. 2010, 54, 522–526. [Google Scholar] [CrossRef]
- Tumpey, T.M.; Kapczynski, D.R.; Swayne, D. Comparative Susceptibility of Chickens and Turkeys to Avian Influenza A H7N2 Virus Infection and Protective Efficacy of a Commercial Avian Influenza H7N2 Virus Vaccine. Avian Dis. 2004, 48, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.; Zhou, N.; Kawaoka, Y.; Webster, R. The surface glycoproteins of H5 influenza viruses isolated from humans, chickens, and wild aquatic birds have distinguishable properties. J. Virol. 1999, 73, 1146–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, Y.; Swayne, D.; Thomas, C.; Rameix-Welti, M.-A.; Naffakh, N.; Warnes, C.; Altholtz, M.; Donis, R.; Subbarao, K. Neuraminidase Stalk Length and Additional Glycosylation of the Hemagglutinin Influence the Virulence of Influenza H5N1 Viruses for Mice. J. Virol. 2009, 83, 4704–4708. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, E.M.; Song, H.; Pena, L.; Perez, D.R. A 27-Amino-Acid Deletion in the Neuraminidase Stalk Supports Replication of an Avian H2N2 Influenza A Virus in the Respiratory Tract of Chickens. J. Virol. 2010, 84, 11831–11840. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human Infection with a Novel Avian-Origin Influenza A (H7N9) Virus. N. Engl. J. Med. 2013, 368, 1888–1897. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Tan, Y.; Wei, K.; Sun, H.; Shi, Y.; Pu, J.; Yang, H.; Gao, G.F.; Yin, Y.; Feng, W.; et al. Amino Acid 316 of Hemagglutinin and the Neuraminidase Stalk Length Influence Virulence of H9N2 Influenza Virus in Chickens and Mice. J. Virol. 2012, 87, 2963–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, J.; Youk, S.; Pantin-Jackwood, M.J.; Suarez, D.L.; Lee, D.-H.; Killian, M.L.; Bergeson, N.H.; Spackman, E. The pathogenicity and transmission of live bird market H2N2 avian influenza viruses in chickens, Pekin ducks, and guinea fowl. Vet. Microbiol. 2021, 260, 109180. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-Z.; Long, J.-X.; Hu, S.-L.; Wu, Y.-T.; Liu, X.-F. Biological significance of amino acids deletion in NA stalk of H5N1 avian influenza virus. Acta Microbiol. Sin. 2006, 46, 542–546. [Google Scholar]
- Luo, G.; Chung, J.; Palese, P. Alterations of the stalk of the influenza virus neuraminidase: Deletions and insertions. Virus Res. 1993, 29, 141–153. [Google Scholar] [PubMed]
- Kim, P.; Jang, Y.H.; Bin Kwon, S.; Lee, C.M.; Han, G.; Seong, B.L. Glycosylation of Hemagglutinin and Neuraminidase of Influenza A Virus as Signature for Ecological Spillover and Adaptation among Influenza Reservoirs. Viruses 2018, 10, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Organisation for Animal Health Avian Influenza Portal. Available online: http://www.oie.int/en/animal-health-in-the-world/update-on-avian-influenza/ (accessed on 5 January 2021).





| Species | Bird Number | Dose Received (log10 EID50) | Virus Titer (log10 EID50/g) | ||||
|---|---|---|---|---|---|---|---|
| Brain | Heart | Lung | Muscle | Spleen | |||
| Turkey | 1 | 6 | 6.5 | 7.7 | 6.3 | 5.9 | 6.0 |
| 2 | 6 | 6.7 | 7.0 | 6.2 | 5.8 | 6.0 | |
| 3 | 6 | 6.6 | 7.7 | 6.2 | 5.7 | 6.3 | |
| Chicken | 1 | 2 | 2.5 | 7.4 | - | 2.4 | - |
| 2 | 4 | 6.8 | 7.5 | 6.4 | 7.0 | 6.8 | |
| 3 | 6 * | 7.0 | 1.8 | 6.3 | 7.0 | 6.7 | |
| LPAIV-1 | LPAIV-2 | HPAIV | |
|---|---|---|---|
| LPAIV-1 | 99.205 | 99.713 | |
| LPAIV-2 | 99.205 | 98.956 | |
| HPAIV | 99.713 | 98.956 |
| LPAIV-1 | LPAIV-2 | HPAIV | Position | Amino Acid Change | Protein | References | Remarks |
|---|---|---|---|---|---|---|---|
| na | na | DRKSRHRRI | 339–347 | Insertion: DRKSRHRRI | Hemagglutinin | Multibasic cleavage site | |
| R | R | K | 98 | R98K | Nucleoprotein | [33] | Change found in samples from bobwhite quail infected with a 2014 H5N2 HPAIV virus from the US 2014–2015 H5 outbreak. Also found in later 2015 virus isolates from turkeys in Minnesota. |
| na | LNCSDTIITYNNTVINNITTTI | na | 56–77 | Deletion: LNCSDTIITYNNTVINNITTTI | Neuraminidase | [16,17,34,35,36,37,38,39] | Associated with adaptation to gallinaceous species |
| G | R | G | 345 | G345R | Neuraminidase | [26] | Found in a 2016 H7N3 virus from Mexico, when compared to an earlier 2012 H7N3 virus. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Criado, M.F.; Leyson, C.M.; Youk, S.; DeBlois, S.; Olivier, T.; Killian, M.L.; Torchetti, M.L.; Parris, D.J.; Spackman, E.; Kapczynski, D.R.; et al. The Pathobiology of H7N3 Low and High Pathogenicity Avian Influenza Viruses from the United States Outbreak in 2020 Differs between Turkeys and Chickens. Viruses 2021, 13, 1851. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091851
Criado MF, Leyson CM, Youk S, DeBlois S, Olivier T, Killian ML, Torchetti ML, Parris DJ, Spackman E, Kapczynski DR, et al. The Pathobiology of H7N3 Low and High Pathogenicity Avian Influenza Viruses from the United States Outbreak in 2020 Differs between Turkeys and Chickens. Viruses. 2021; 13(9):1851. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091851
Chicago/Turabian StyleCriado, Miriã F., Christina M. Leyson, Sungsu Youk, Suzanne DeBlois, Tim Olivier, Mary Lea Killian, Mia L. Torchetti, Darren J. Parris, Erica Spackman, Darrell R. Kapczynski, and et al. 2021. "The Pathobiology of H7N3 Low and High Pathogenicity Avian Influenza Viruses from the United States Outbreak in 2020 Differs between Turkeys and Chickens" Viruses 13, no. 9: 1851. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091851