
Inhibitory Effect of IL-1β on HBV and HDV Replication and HBs Antigen-Dependent Modulation of Its Secretion by Macrophages

 , and
, and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Cells Purification and Cells Culture
2.2. Viral Inoculation
2.3. Cytokine and Drugs
2.4. SiRNA Treatment
2.5. Quantification of Secreted Proteins by ELISA
2.6. RNA Extraction and RT-qPCR
2.7. Statistical Analysis
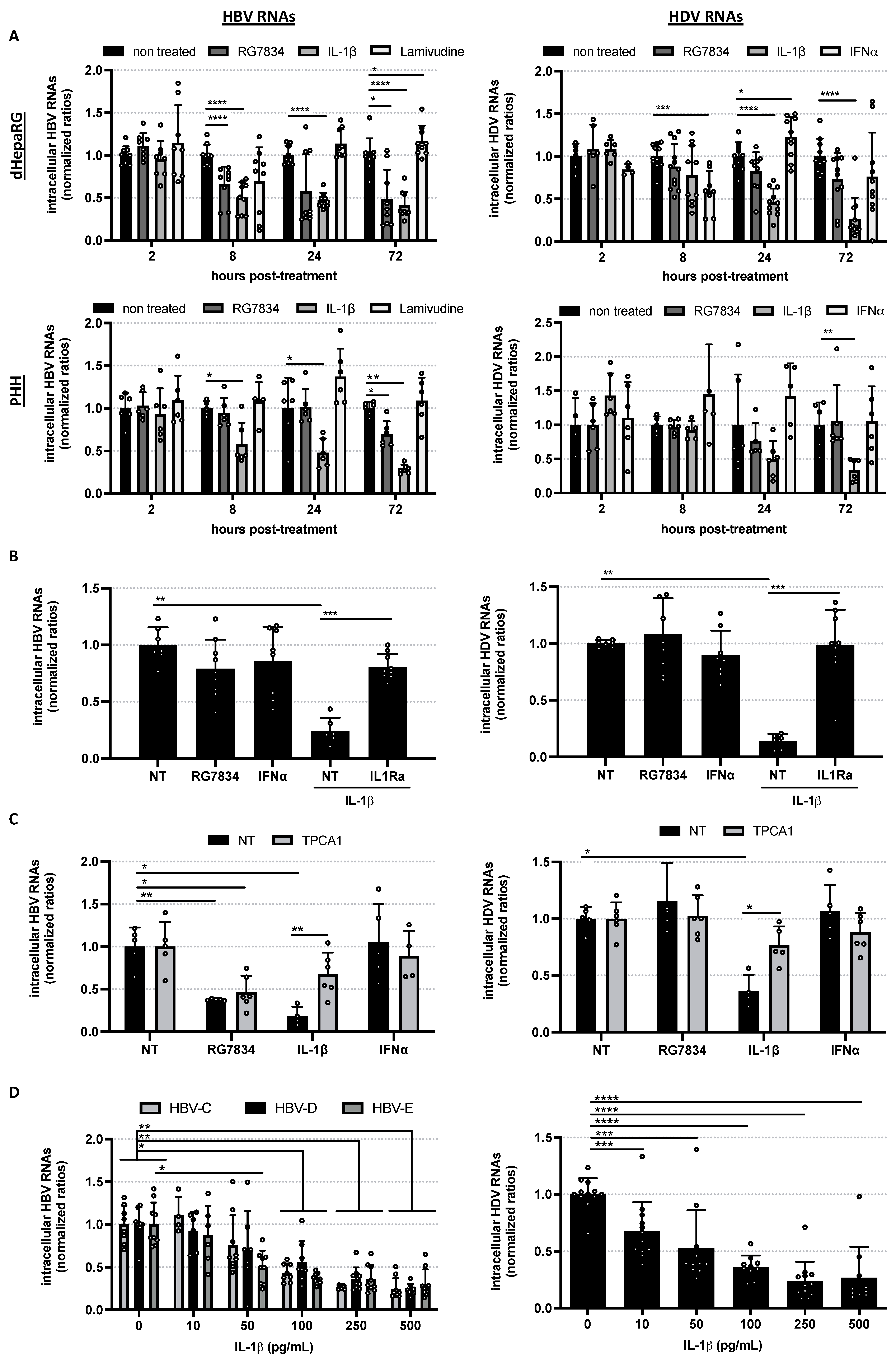
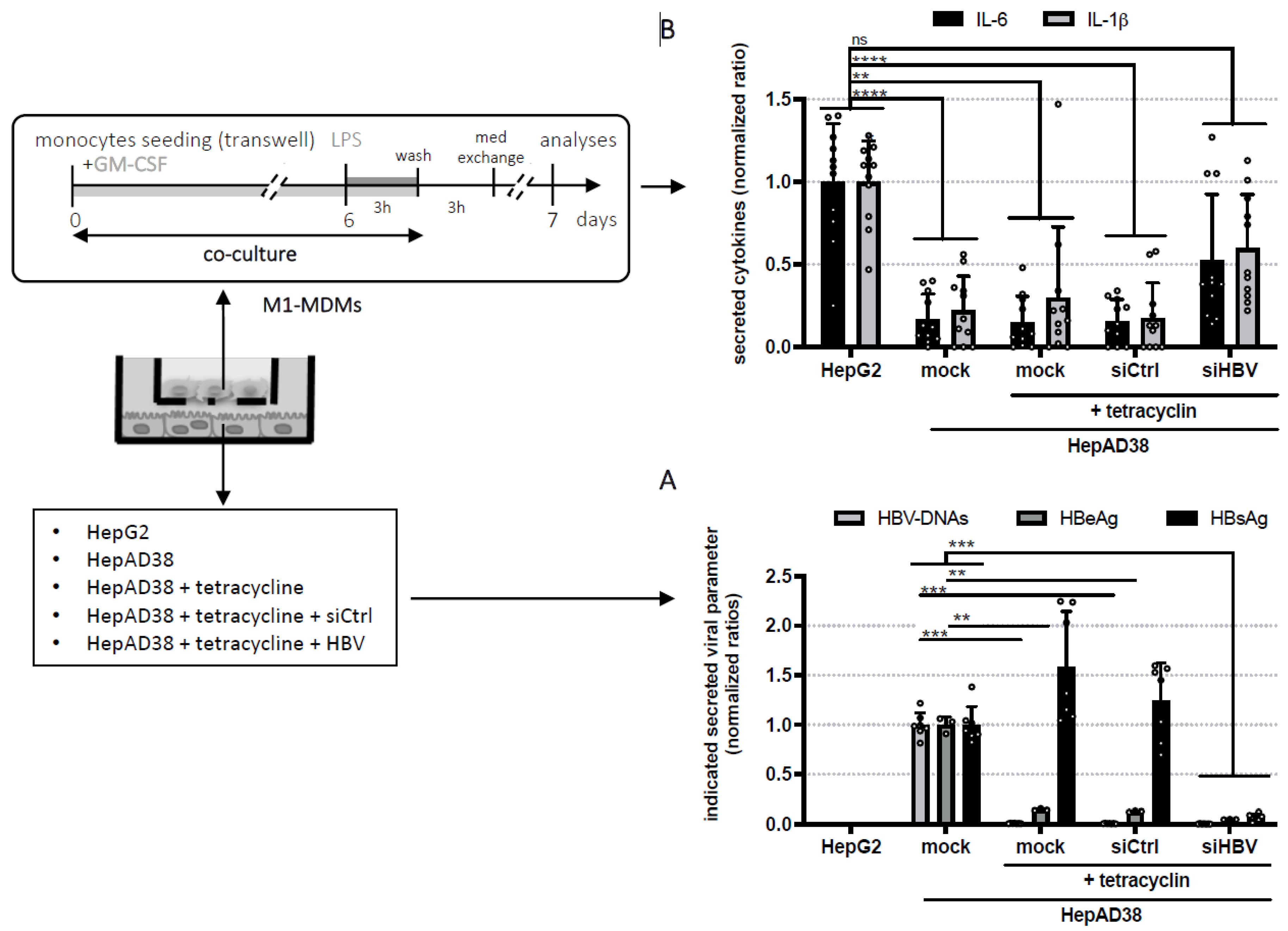
3. Results and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartoli, A.; Gabrielli, F.; Tassi, A.; Cursaro, C.; Pinelli, A.; Andreone, P. Treatments for HBV: A Glimpse into the Future. Viruses 2021, 13, 1767. [Google Scholar] [CrossRef]
- Rizzetto, M. The Adventure of Delta. Liver Int. 2016, 36, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Tout, I.; Loureiro, D.; Mansouri, A.; Soumelis, V.; Boyer, N.; Asselah, T. Hepatitis B Surface Antigen Seroclearance: Immune Mechanisms, Clinical Impact, Importance for Drug Development. J. Hepatol. 2020, 73, 409–422. [Google Scholar] [CrossRef]
- Vaillant, A. HBsAg, Subviral Particles, and Their Clearance in Establishing a Functional Cure of Chronic Hepatitis B Virus Infection. ACS Infect Dis. 2021, 7, 1351–1368. [Google Scholar] [CrossRef]
- Wranke, A.; Hardtke, S.; Heidrich, B.; Dalekos, G.; Yalçin, K.; Tabak, F.; Gürel, S.; Çakaloğlu, Y.; Akarca, U.S.; Lammert, F.; et al. Ten-Year Follow-up of a Randomized Controlled Clinical Trial in Chronic Hepatitis Delta. J. Viral. Hepat. 2020, 27, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Bremer, B.; Anastasiou, O.E.; Hardtke, S.; Caruntu, F.A.; Curescu, M.G.; Yalcin, K.; Akarca, U.S.; Gürel, S.; Zeuzem, S.; Erhardt, A.; et al. Residual Low HDV Viraemia Is Associated HDV RNA Relapse after PEG-IFNa-Based Antiviral Treatment of Hepatitis Delta: Results from the HIDIT-II Study. Liver Int. 2021, 41, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A. Genotypes and Genetic Variability of Hepatitis B Virus. INT 2014, 57, 141–150. [Google Scholar] [CrossRef]
- Ladner, S.K.; Otto, M.J.; Barker, C.S.; Zaifert, K.; Wang, G.H.; Guo, J.T.; Seeger, C.; King, R.W. Inducible Expression of Human Hepatitis B Virus (HBV) in Stably Transfected Hepatoblastoma Cells: A Novel System for Screening Potential Inhibitors of HBV Replication. Antimicrob. Agents Chemother. 1997, 41, 1715–1720. [Google Scholar] [CrossRef] [Green Version]
- Faure-Dupuy, S.; Delphin, M.; Aillot, L.; Dimier, L.; Lebossé, F.; Fresquet, J.; Parent, R.; Sebastian Matter, M.; Rivoire, M.; Bendriss-Vermare, N.; et al. Hepatitis B Virus-Induced Modulation of Liver Macrophage Function Promotes Hepatocyte Infection. J. Hepatol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Alfaiate, D.; Lucifora, J.; Abeywickrama-Samarakoon, N.; Michelet, M.; Testoni, B.; Cortay, J.-C.; Sureau, C.; Zoulim, F.; Dény, P.; Durantel, D. HDV RNA Replication Is Associated with HBV Repression and Interferon-Stimulated Genes Induction in Super-Infected Hepatocytes. Antivir. Res. 2016, 136, 19–31. [Google Scholar] [CrossRef]
- Luangsay, S.; Gruffaz, M.; Isorce, N.; Testoni, B.; Michelet, M.; Faure-Dupuy, S.; Maadadi, S.; Ait-Goughoulte, M.; Parent, R.; Rivoire, M.; et al. Early Inhibition of Hepatocyte Innate Responses by Hepatitis B Virus. J. Hepatol. 2015, 63, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Watashi, K.; Tsukuda, S.; Aly, H.H.; Fukasawa, M.; Fujimoto, A.; Suzuki, R.; Aizaki, H.; Ito, T.; Koiwai, O.; et al. Evaluation and Identification of Hepatitis B Virus Entry Inhibitors Using HepG2 Cells Overexpressing a Membrane Transporter NTCP. Biochem. Biophys. Res. Commun. 2014, 443, 808–813. [Google Scholar] [CrossRef] [Green Version]
- Lucifora, J.; Salvetti, A.; Marniquet, X.; Mailly, L.; Testoni, B.; Fusil, F.; Inchauspé, A.; Michelet, M.; Michel, M.-L.; Levrero, M.; et al. Detection of the Hepatitis B Virus (HBV) Covalently-Closed-Circular DNA (CccDNA) in Mice Transduced with a Recombinant AAV-HBV Vector. Antiviral Res. 2017, 145, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.; Tal, G.; Lider, O.; Shaul, Y. Cytokine Induction by the Hepatitis B Virus Capsid in Macrophages Is Facilitated by Membrane Heparan Sulfate and Involves TLR2. J. Immunol. 2005, 175, 3165–3176. [Google Scholar] [CrossRef]
- Isorce, N.; Testoni, B.; Locatelli, M.; Fresquet, J.; Rivoire, M.; Luangsay, S.; Zoulim, F.; Durantel, D. Antiviral Activity of Various Interferons and Pro-Inflammatory Cytokines in Non-Transformed Cultured Hepatocytes Infected with Hepatitis B Virus. Antiviral Res. 2016, 130, 36–45. [Google Scholar] [CrossRef]
- Watashi, K.; Liang, G.; Iwamoto, M.; Marusawa, H.; Uchida, N.; Daito, T.; Kitamura, K.; Muramatsu, M.; Ohashi, H.; Kiyohara, T.; et al. Interleukin-1 and Tumor Necrosis Factor-α Trigger Restriction of Hepatitis B Virus Infection via a Cytidine Deaminase Activation-Induced Cytidine Deaminase (AID). J. Biol. Chem. 2013, 288, 31715–31727. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Zhou, C.; Jiang, M.; Wang, Y.; Wang, J.; Cheng, Z.; Wang, M.; Liu, Y.; Liang, C.; Wang, J.; et al. Discovery of RG7834: The First-in-Class Selective and Orally Available Small Molecule Hepatitis B Virus Expression Inhibitor with Novel Mechanism of Action. J. Med. Chem. 2018, 61, 10619–10634. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Weil, S.; Martin, M.U. The Family of the Interleukin-1 Receptors. Immunol. Rev. 2018, 281, 197–232. [Google Scholar] [CrossRef]
- Nan, J.; Du, Y.; Chen, X.; Bai, Q.; Wang, Y.; Zhang, X.; Zhu, N.; Zhang, J.; Hou, J.; Wang, Q.; et al. TPCA-1 Is a Direct Dual Inhibitor of STAT3 and NF-ΚB and Regresses Mutant EGFR-Associated Human Non-Small Cell Lung Cancers. Mol. Cancer Ther. 2014, 13, 617–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faure-Dupuy, S.; Durantel, D.; Lucifora, J. Liver Macrophages: Friend or Foe during Hepatitis B Infection? Liver Int. 2018, 38, 1718–1729. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, N.R.; Chadha, R.; Kumar, S.; Choedon, T.; Reddy, V.S.; Kumar, V. The HBx Gene of Hepatitis B Virus Can Influence Hepatic Microenvironment via Exosomes by Transferring Its MRNA and Protein. Virus Res. 2017, 240, 166–174. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delphin, M.; Faure-Dupuy, S.; Isorce, N.; Rivoire, M.; Salvetti, A.; Durantel, D.; Lucifora, J. Inhibitory Effect of IL-1β on HBV and HDV Replication and HBs Antigen-Dependent Modulation of Its Secretion by Macrophages. Viruses 2022, 14, 65. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010065
Delphin M, Faure-Dupuy S, Isorce N, Rivoire M, Salvetti A, Durantel D, Lucifora J. Inhibitory Effect of IL-1β on HBV and HDV Replication and HBs Antigen-Dependent Modulation of Its Secretion by Macrophages. Viruses. 2022; 14(1):65. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010065
Chicago/Turabian StyleDelphin, Marion, Suzanne Faure-Dupuy, Nathalie Isorce, Michel Rivoire, Anna Salvetti, David Durantel, and Julie Lucifora. 2022. "Inhibitory Effect of IL-1β on HBV and HDV Replication and HBs Antigen-Dependent Modulation of Its Secretion by Macrophages" Viruses 14, no. 1: 65. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010065