
Comparative Pharmacokinetics of a Dual Inhibitor of HIV-1, NBD-14189, in Rats and Dogs with a Proof-of-Concept Evaluation of Antiviral Potency in SCID-hu Mouse Model
 , ,
, ,
Abstract
:1. Introduction
2. Materials & Methods
2.1. PK Study in Rats and Dogs
2.2. Drugs and Viruses
2.3. In Vitro Antiviral Assay
2.4. SCID-hu Thy/Liv Mice
2.5. Flow Cytometry
3. Results and Discussions
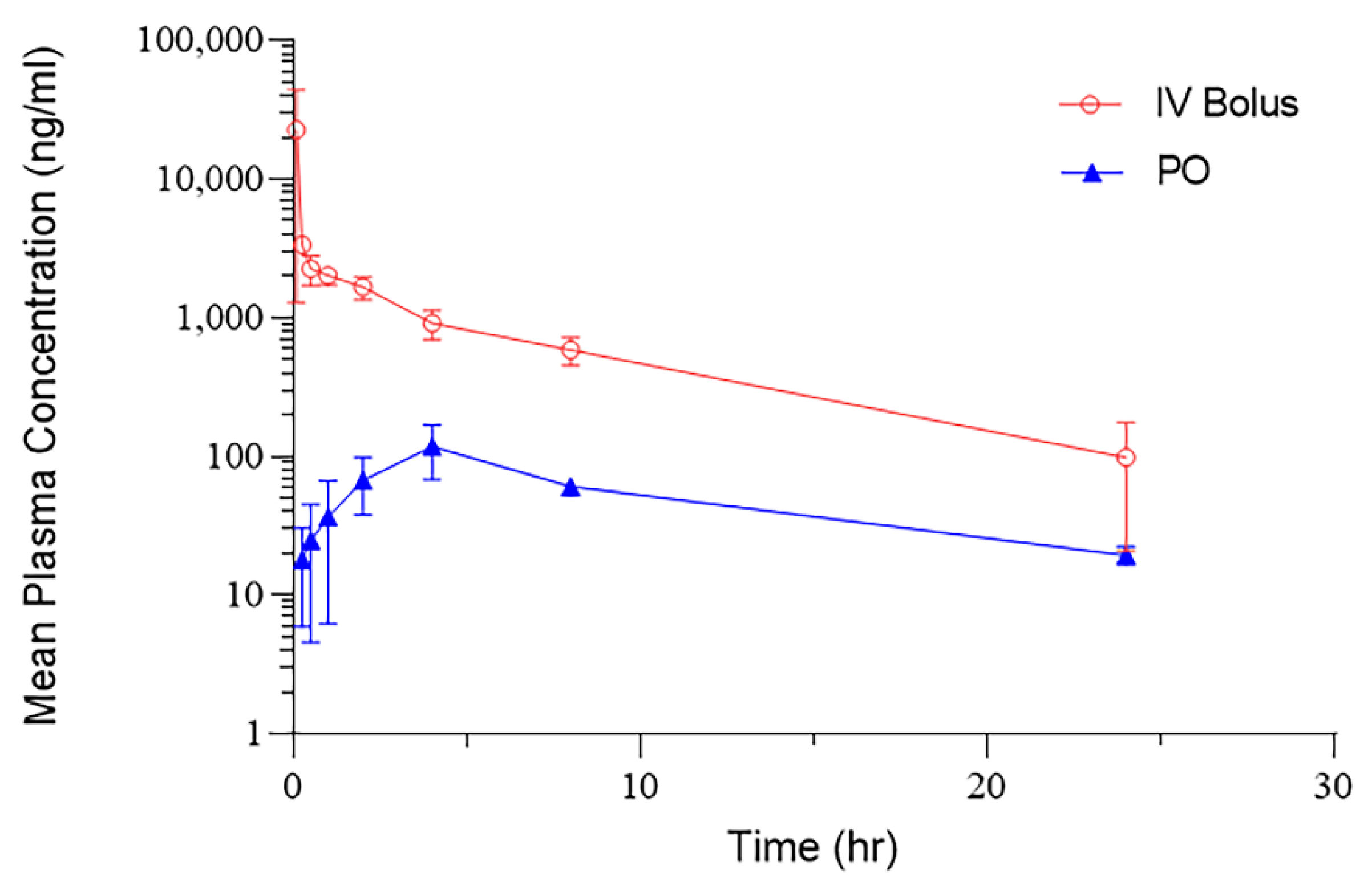
3.1. Pharmacokinetic Study in Rat
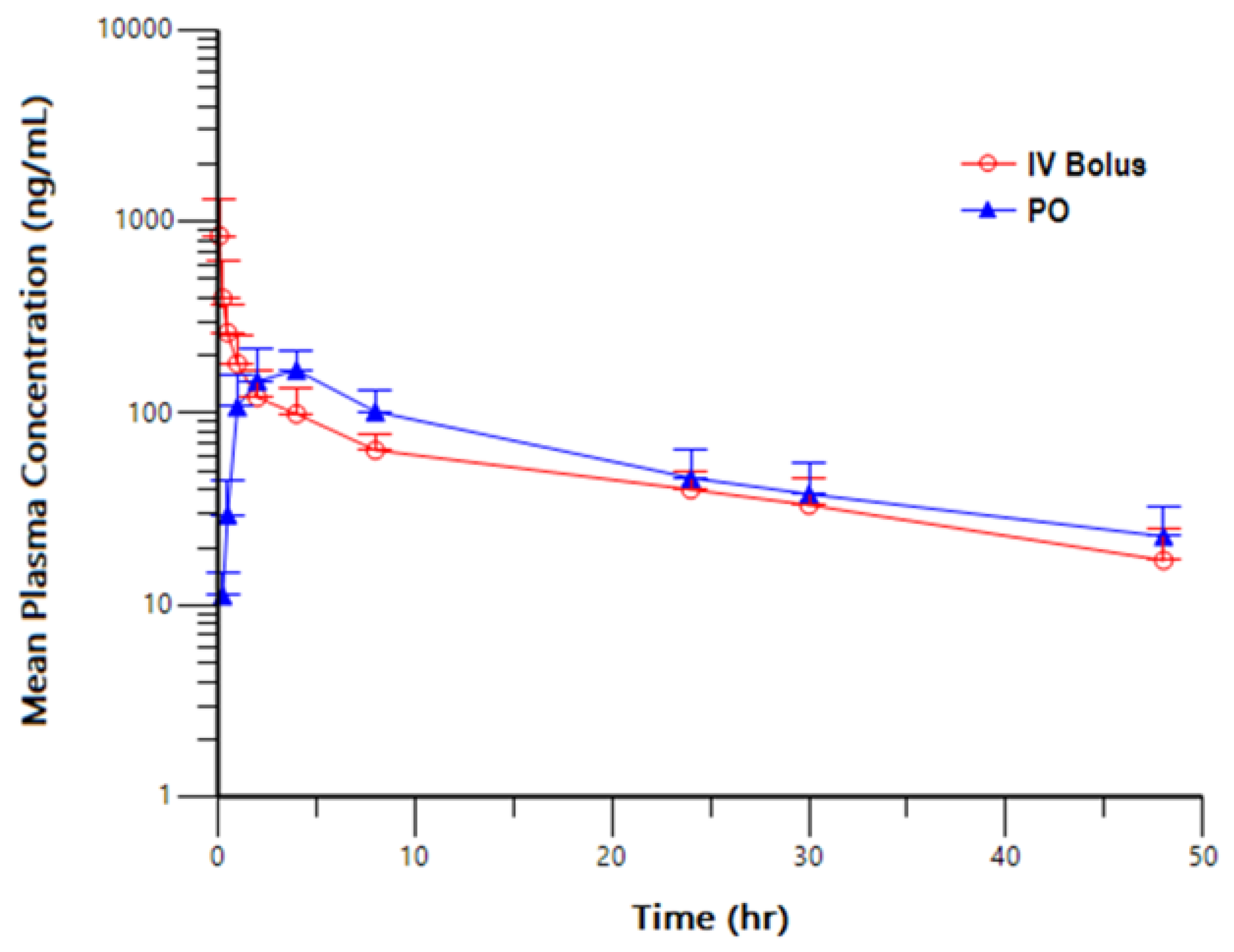
3.2. Pharmacokinetic Study in Dogs
3.3. Evaluation of the Anti-HIV-1 Activity of NBD-14189 against NL4-3 Virus in SCID-hu Thy/Liv Mice
3.4. NBD-14189 Tolerability and Plasma Levels in SCID-hu Thy/Liv Mice
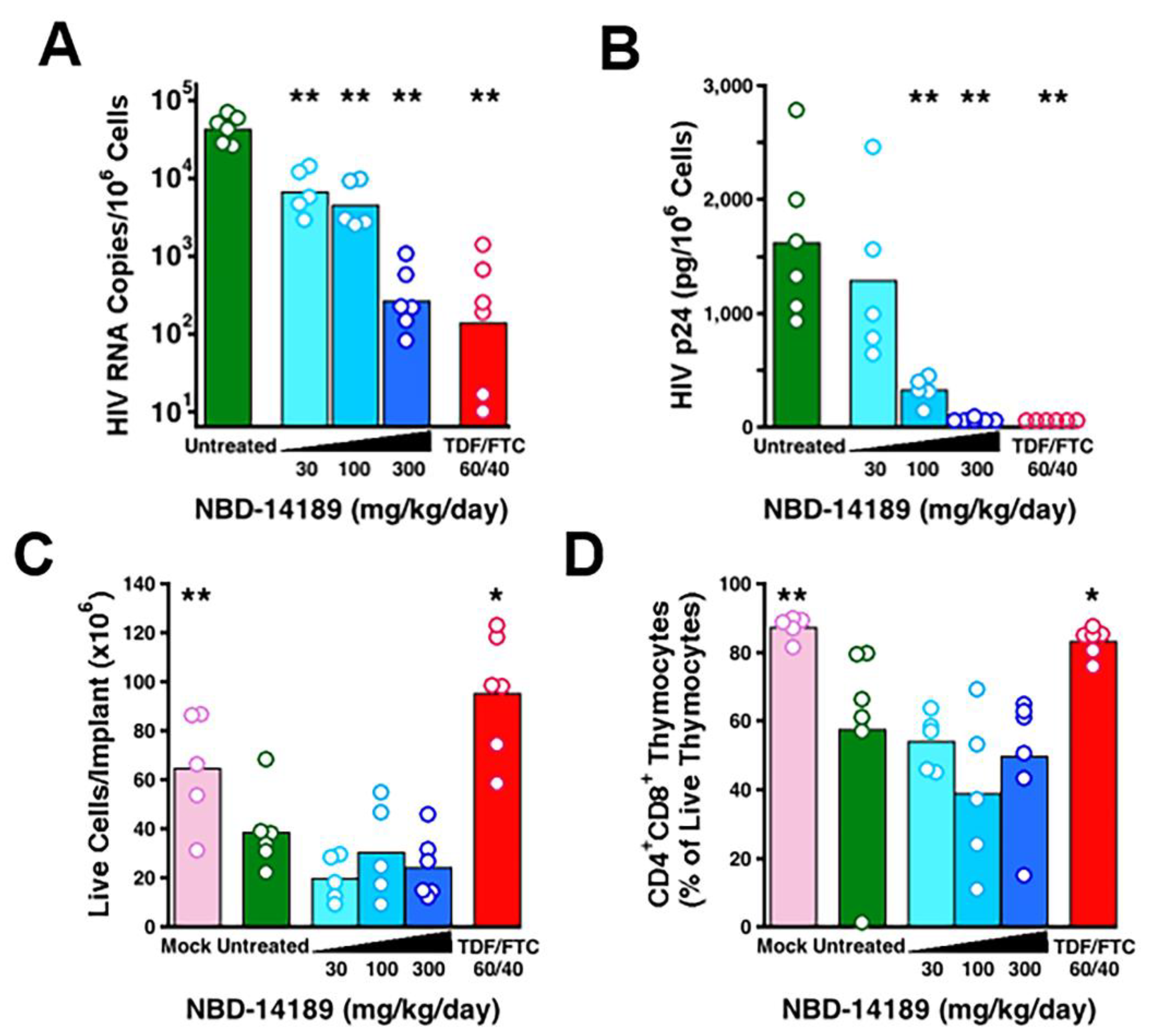
3.5. NBD-14189 in HIV-Infected SCID-hu Thy/Liv Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curreli, F.; Ahmed, S.; Benedict Victor, S.M.; Iusupov, I.R.; Belov, D.S.; Markov, P.O.; Kurkin, A.V.; Altieri, A.; Debnath, A.K. Preclinical Optimization of gp120 Entry Antagonists as anti-HIV-1 Agents with Improved Cytotoxicity and ADME Properties through Rational Design, Synthesis, and Antiviral Evaluation. J. Med. Chem. 2020, 63, 1724–1749. [Google Scholar] [CrossRef] [PubMed]
- Curreli, F.; Belov, D.S.; Kwon, Y.D.; Ramesh, R.; Furimsky, A.M.; O’Loughlin, K.; Byrge, P.C.; Iyer, L.V.; Mirsalis, J.C.; Kurkin, A.V.; et al. Structure-based lead optimization to improve antiviral potency and ADMET properties of phenyl-1H-pyrrole-carboxamide entry inhibitors targeted to HIV-1 gp120. Eur. J. Med. Chem. 2018, 154, 367–391. [Google Scholar] [CrossRef]
- Curreli, F.; Belov, D.S.; Ahmed, S.; Ramesh, R.R.; Kurkin, A.V.; Altieri, A.; Debnath, A.K. Synthesis, antiviral activity, and structure-activity relationship of 1,3-benzodioxolyl pyrrole-based entry inhibitors targeting the Phe43 cavity in HIV-1 gp120. ChemMedChem 2018, 13, 2332–2348. [Google Scholar] [CrossRef] [PubMed]
- Belov, D.S.; Curreli, F.; Kurkin, A.V.; Altieri, A.; Debnath, A.K. Guanidine-containing phenyl-pyrrole compounds as probes for generating HIV nntry inhibitors targeted to gp120. ChemistrySelect 2018, 3, 6450–6453. [Google Scholar] [CrossRef]
- Curreli, F.; Kwon, Y.D.; Belov, D.S.; Ramesh, R.R.; Kurkin, A.V.; Altieri, A.; Kwong, P.D.; Debnath, A.K. Synthesis, antiviral potency, in vitro ADMET and X-ray structure of potent CD4-mimics as entry inhibitors that target the Phe43 cavity of HIV-1 gp120. J. Med. Chem. 2017, 60, 3124–3153. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Yu, F.; Cai, L.; Debnath, A.K.; Jiang, S. Development of Small-molecule HIV Entry Inhibitors Specifically Targeting gp120 or gp41. Curr. Top. Med. Chem. 2016, 16, 1074–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.D.; Lalonde, J.M.; Yang, Y.; Elban, M.A.; Sugawara, A.; Courter, J.R.; Jones, D.M.; Smith, A.B., III; Debnath, A.K.; Kwong, P.D. Crystal structures of HIV-1 gp120 envelope glycoprotein in complex with NBD analogues that target the CD4-binding site. PLoS ONE 2014, 9, e85940. [Google Scholar] [CrossRef]
- Kwon, Y.D.; Finzi, A.; Wu, X.; Dogo-Isonagie, C.; Lee, L.K.; Moore, L.R.; Schmidt, S.D.; Stuckey, J.; Yang, Y.; Zhou, T.; et al. Unliganded HIV-1 gp120 core structures assume the CD4-bound conformation with regulation by quaternary interactions and variable loops. Proc. Natl. Acad. Sci. USA 2012, 109, 5663–5668. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Ma, L.; Jiang, S.; Lu, H.; Liu, S.; He, Y.; Strick, N.; Neamati, N.; Debnath, A.K. Identification of N-phenyl-N’-(2,2,6,6-tetramethyl-piperidin-4-yl)-oxalamides as a new class of HIV-1 entry inhibitors that prevent gp120 binding to CD4. Virology 2005, 339, 213–225. [Google Scholar] [CrossRef]
- Pancera, M.; Lai, Y.T.; Bylund, T.; Druz, A.; Narpala, S.; O’Dell, S.; Schon, A.; Bailer, R.T.; Chuang, G.Y.; Geng, H.; et al. Crystal structures of trimeric HIV envelope with entry inhibitors BMS-378806 and BMS-626529. Nat. Chem. Biol. 2017, 13, 1115–1122. [Google Scholar] [CrossRef]
- Grant, P.M.; Kozal, M.J. Fostemsavir: A first-in-class HIV-1 attachment inhibitor. Curr. Opin. HIV AIDS 2022, 17, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Lataillade, M.; Lalezari, J.P.; Kozal, M.; Aberg, J.A.; Pialoux, G.; Cahn, P.; Thompson, M.; Molina, J.M.; Moreno, S.; Grinsztejn, B.; et al. Safety and efficacy of the HIV-1 attachment inhibitor prodrug fostemsavir in heavily treatment-experienced individuals: Week 96 results of the phase 3 BRIGHTE study. Lancet HIV 2020, 7, e740–e751. [Google Scholar] [CrossRef]
- Zhou, N.; Nowicka-Sans, B.; McAuliffe, B.; Ray, N.; Eggers, B.; Fang, H.; Fan, L.; Healy, M.; Langley, D.R.; Hwang, C.; et al. Genotypic correlates of susceptibility to HIV-1 attachment inhibitor BMS-626529, the active agent of the prodrug BMS-663068. J. Antimicrob. Chemother. 2014, 69, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
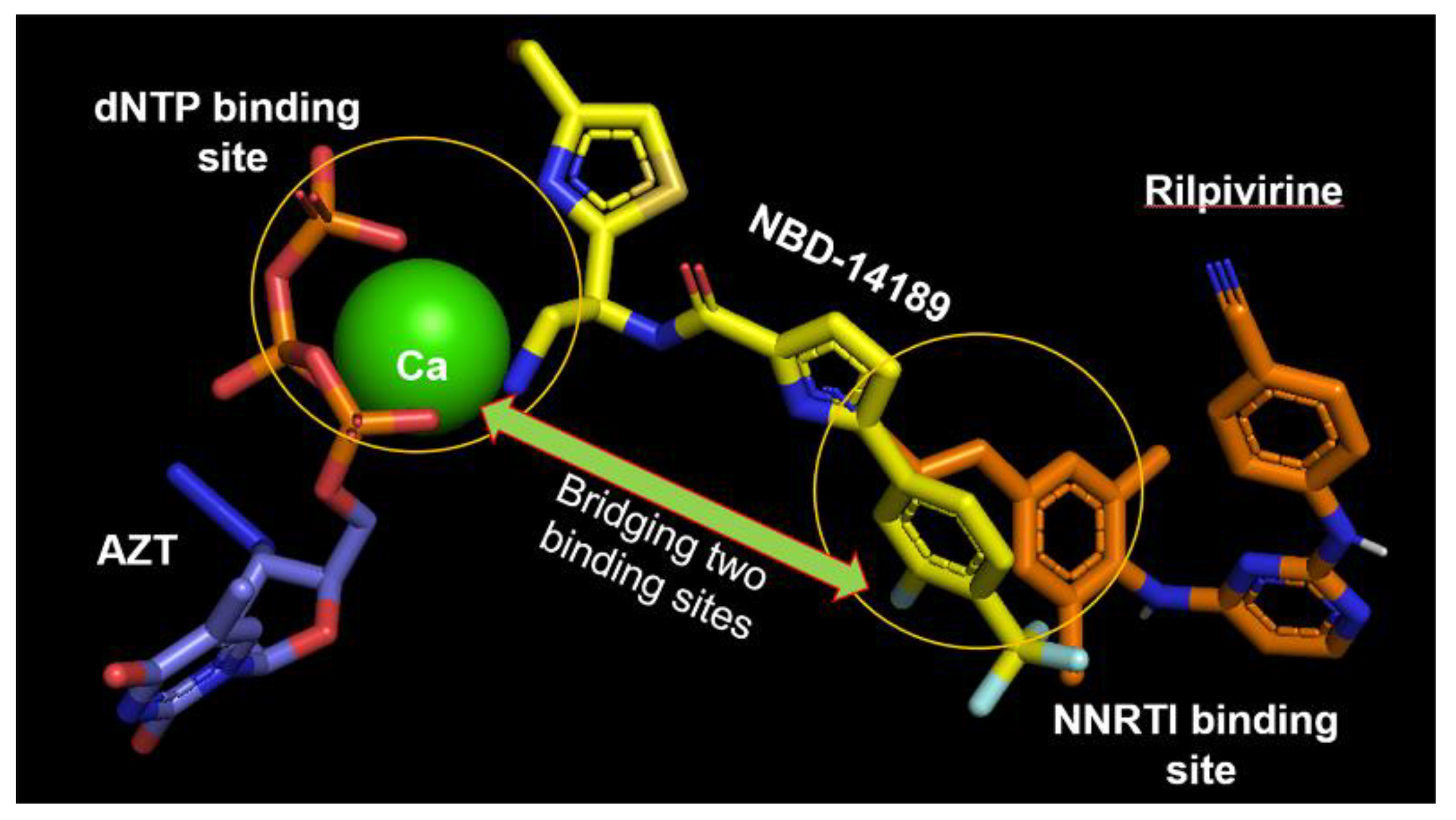
- Losada, N.; Ruiz, F.X.; Curreli, F.; Gruber, K.; Pilch, A.; Das, K.; Debnath, A.K.; Arnold, E. HIV-1 gp120 Antagonists Also Inhibit HIV-1 Reverse Transcriptase by Bridging the NNRTI and NRTI Sites. J. Med. Chem. 2021, 64, 16530–16540. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.T.; Tawa, P.; Auger, A.; Wang, D.; Su, H.P.; Yan, Y.; Hazuda, D.J.; Miller, M.D.; Asante-Appiah, E.; Melnyk, R.A. Identification of novel bifunctional HIV-1 reverse transcriptase inhibitors. J. Antimicrob. Chemother. 2018, 73, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himmel, D.M.; Sarafianos, S.G.; Dharmasena, S.; Hossain, M.M.; McCoy-Simandle, K.; Ilina, T.; Clark, A.D., Jr.; Knight, J.L.; Julias, J.G.; Clark, P.K.; et al. HIV-1 reverse transcriptase structure with RNase H inhibitor dihydroxy benzoyl naphthyl hydrazone bound at a novel site. ACS Chem. Biol. 2006, 1, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, D.G.; Bauman, J.D.; Challa, J.R.; Patel, D.; Troxler, T.; Das, K.; Arnold, E.; Hochstrasser, R.M. Snapshot of the equilibrium dynamics of a drug bound to HIV-1 reverse transcriptase. Nat. Chem. 2013, 5, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Balzarini, J.; Miller, M.T.; Maguire, A.R.; DeStefano, J.J.; Arnold, E. Conformational States of HIV-1 Reverse Transcriptase for Nucleotide Incorporation vs. Pyrophosphorolysis-Binding of Foscarnet. ACS Chem. Biol. 2016, 11, 2158–2164. [Google Scholar] [CrossRef] [Green Version]
- Waring, M.J.; Arrowsmith, J.; Leach, A.R.; Leeson, P.D.; Mandrell, S.; Owen, R.M.; Pairaudeau, G.; Pennie, W.D.; Pickett, S.D.; Wang, J.; et al. An analysis of the attrition of drug candidates from four major pharmaceutical companies. Nat. Rev. Drug Discov. 2015, 14, 475–486. [Google Scholar] [CrossRef]
- van de Waterbeemd, H.; Gifford, E. ADMET in silico modelling: Towards prediction paradise? Nat. Rev. Drug Discov. 2003, 2, 192–204. [Google Scholar] [CrossRef]
- Williams, J.; Siramshetty, V.; Nguyen Eth, T.; Padilha, E.C.; Kabir, M.; Yu, K.R.; Wang, A.Q.; Zhao, T.; Itkin, M.; Shinn, P.; et al. Using in vitro ADME data for lead compound selection: An emphasis on PAMPA pH 5 permeability and oral bioavailability. Bioorg. Med. Chem. 2022, 56, 116588. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.J.; Sproston, J.L.; Barton, P.; Riley, R.J. Estimating human ADME properties, pharmacokinetic parameters and likely clinical dose in drug discovery. Expert Opin. Drug Discov. 2019, 14, 1313–1327. [Google Scholar] [CrossRef] [PubMed]
- Kathia, M.H.; Moda, T.L.; Andricopulo, A.D. Pharmacokinetic Properties and In Silico ADME Modeling in Drug Discovery. Med. Chem. 2013, 9, 163–176. [Google Scholar]
- Caldwell, G.W.; Yan, Z.; Tang, W.; Dasgupta, M.; Hasting, B. ADME optimization and toxicity assessment in early- and late-phase drug discovery. Curr. Top. Med. Chem. 2009, 9, 965–980. [Google Scholar] [CrossRef]
- Sharma, P.; Patel, N.; Prasad, B.; Varma, M.V.S. Pharmacokinetics: Theory and Application in Drug Discovery and Development. In Drug Discovery and Development: From Targets and Molecules to Medicines; Poduri, R., Ed.; Springer: Singapore, 2021; pp. 297–355. [Google Scholar]
- Ledgerwood, J.E.; Coates, E.E.; Yamshchikov, G.; Saunders, J.G.; Holman, L.; Enama, M.E.; DeZure, A.; Lynch, R.M.; Gordon, I.; Plummer, S.; et al. Safety, pharmacokinetics and neutralization of the broadly neutralizing HIV-1 human monoclonal antibody VRC01 in healthy adults. Clin. Exp. Immunol. 2015, 182, 289–301. [Google Scholar]
- Hashimoto, C.; Narumi, T.; Otsuki, H.; Hirota, Y.; Arai, H.; Yoshimura, K.; Harada, S.; Ohashi, N.; Nomura, W.; Miura, T.; et al. A CD4 mimic as an HIV entry inhibitor: Pharmacokinetics. Bioorg. Med. Chem. 2013, 21, 7884–7889. [Google Scholar] [CrossRef]
- Hanna, G.J.; Lalezari, J.; Hellinger, J.A.; Wohl, D.A.; Nettles, R.; Persson, A.; Krystal, M.; Lin, P.; Colonno, R.; Grasela, D.M. Antiviral activity, pharmacokinetics, and safety of BMS-488043, a novel oral small-molecule HIV-1 attachment inhibitor, in HIV-1-infected subjects. Antimicrob. Agents Chemother. 2011, 55, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Gao, W.; Hu, H.; Zhou, S. Why 90% of clinical drug development fails and how to improve it? Acta Pharm. Sin. B 2022, 12, 3049–3062. [Google Scholar] [CrossRef]
- Lai, Y.T.; Wang, T.; O’Dell, S.; Louder, M.K.; Schon, A.; Cheung, C.S.F.; Chuang, G.Y.; Druz, A.; Lin, B.; McKee, K.; et al. Lattice engineering enables definition of molecular features allowing for potent small-molecule inhibition of HIV-1 entry. Nat. Commun. 2019, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Ueda, Y.; Zhang, Z.; Yin, Z.; Matiskella, J.; Pearce, B.C.; Yang, Z.; Zheng, M.; Parker, D.D.; Yamanaka, G.A.; et al. Discovery of the Human Immunodeficiency Virus Type 1 (HIV-1) Attachment Inhibitor Temsavir and Its Phosphonooxymethyl Prodrug Fostemsavir. J. Med. Chem. 2018, 61, 6308–6327. [Google Scholar] [CrossRef]
- Rabin, L.; Hincenbergs, M.; Moreno, M.B.; Warren, S.; Linquist, V.; Datema, R.; Charpiot, B.; Seifert, J.; Kaneshima, H.; McCune, J.M. Use of standardized SCID-hu Thy/Liv mouse model for preclinical efficacy testing of anti-human immunodeficiency virus type 1 compounds. Antimicrob. Agents Chemother. 1996, 40, 755–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddart, C.A.; Bales, C.A.; Bare, J.C.; Chkhenkeli, G.; Galkina, S.A.; Kinkade, A.N.; Moreno, M.E.; Rivera, J.M.; Ronquillo, R.E.; Sloan, B.; et al. Validation of the SCID-hu Thy/Liv mouse model with four classes of licensed antiretrovirals. PLoS ONE 2007, 2, e655. [Google Scholar] [CrossRef]
- Stoddart, C.A.; Joshi, P.; Sloan, B.; Bare, J.C.; Smith, P.C.; Allaway, G.P.; Wild, C.T.; Martin, D.E. Potent activity of the HIV-1 maturation inhibitor bevirimat in SCID-hu Thy/Liv mice. PLoS ONE 2007, 2, e1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddart, C.A.; Moreno, M.E.; Linquist-Stepps, V.D.; Bare, C.; Bogan, M.R.; Gobbi, A.; Buckheit, R.W., Jr.; Bedard, J.; Rando, R.F.; McCune, J.M. Antiviral activity of 2’-deoxy-3’-oxa-4’-thiocytidine (BCH-10652) against lamivudine-resistant human immunodeficiency virus type 1 in SCID-hu Thy/Liv mice. Antimicrob. Agents Chemother. 2000, 44, 783–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddart, C.A.; Nault, G.; Galkina, S.A.; Bousquet-Gagnon, N.; Bridon, D.; Quraishi, O. Preexposure prophylaxis with albumin-conjugated C34 peptide HIV-1 fusion inhibitor in SCID-hu Thy/Liv mice. Antimicrob. Agents Chemother. 2012, 56, 2162–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddart, C.A.; Nault, G.; Galkina, S.A.; Thibaudeau, K.; Bakis, P.; Bousquet-Gagnon, N.; Robitaille, M.; Bellomo, M.; Paradis, V.; Liscourt, P.; et al. Albumin-conjugated C34 peptide HIV-1 fusion inhibitor: Equipotent to C34 and T-20 in vitro with sustained activity in SCID-hu Thy/Liv mice. J. Biol. Chem. 2008, 283, 34045–34052. [Google Scholar] [CrossRef] [Green Version]
- Stoddart, C.A.; Rabin, L.; Hincenbergs, M.; Moreno, M.; Linquist-Stepps, V.; Leeds, J.M.; Truong, L.A.; Wyatt, J.R.; Ecker, D.J.; McCune, J.M. Inhibition of human immunodeficiency virus type 1 infection in SCID-hu Thy/Liv mice by the G-quartet-forming oligonucleotide, ISIS 5320. Antimicrob. Agents Chemother. 1998, 42, 2113–2115. [Google Scholar] [CrossRef] [Green Version]
- Adachi, A.; Gendelman, H.E.; Koenig, S.; Folks, T.; Willey, R.; Rabson, A.; Martin, M.A. Production of acquired immunideficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol. 1986, 59, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Nowicka-Sans, B.; Gong, Y.F.; McAuliffe, B.; Dicker, I.; Ho, H.T.; Zhou, N.; Eggers, B.; Lin, P.F.; Ray, N.; Wind-Rotolo, M.; et al. In vitro antiviral characteristics of HIV-1 attachment inhibitor BMS-626529, the active component of the prodrug BMS-663068. Antimicrob. Agents Chemother. 2012, 56, 3498–3507. [Google Scholar] [CrossRef] [Green Version]
- Kirby, K.A.; Michailidis, E.; Fetterly, T.L.; Steinbach, M.A.; Singh, K.; Marchand, B.; Leslie, M.D.; Hagedorn, A.N.; Kodama, E.N.; Marquez, V.E.; et al. Effects of substitutions at the 4’ and 2 positions on the bioactivity of 4’-ethynyl-2-fluoro-2’-deoxyadenosine. Antimicrob. Agents Chemother. 2013, 57, 6254–6264. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PK Parameters | IV | PO | Unit |
|---|---|---|---|
| C0 | 58,750 | ng/mL | |
| Cmax | 22,690 | 118 | ng/mL |
| T1/2 | 7.58 | 8.19 | H |
| AUC | 22,076 | 1482 | ng/mL*h |
| CL | 0.0005 | 0.0067 | (mg/kg)/(ng/mL)/h |
| Vss | 0.003 | (mg/kg)/(ng/mL) |
| Dose Level (mg/kg) | Analyte | Route | C0 (ng/mL) | Tmax (h) | Cmax (ng/mL) | Tlast (h) | AUClast (h*ng/mL) | T 1/2 (h) | AUCINF (h*ng/mL) | Cl (mL/min/kg) | Vz (L/kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | NBD-14189 | IV Bolus | 1230 | 0.083 | 844 | 48 | 2600 | 20.0 | 3130 | 5.77 | 9.60 |
| 2 | NBD-14189 | PO | - | 3.3 | 181 | 48 | 3020 | 24.3 | 3820 | - | - |
| Analyte | Dose Level (mg/kg) | Route | AUCinf/Dose (h*kg*ng/mL/mg) | F(%) |
|---|---|---|---|---|
| NBD-14189 | 1 | IV Bolus | 3130 | - |
| NBD-14189 | 2 | PO | 1910 | 61.0 |
| Drug | NL4-3 Antiviral (PBMC) (µM) | PBMC Toxicity (µM) | SI | ||
|---|---|---|---|---|---|
| IC50 | IC90 | CC50 | CC90 | CC50/IC50 | |
| NBD-14168 | 1.2 | >10 | 7.6 | >100 | 6.4 |
| NBD-14189 | 0.38 | >10 | 15 | >100 | 39 |
| BMS-626529 | 0.28 | >10 | 32 | >100 | 110 |
| EFdA | 0.0017 | >0.066 | 23 | >100 | 13,000 |
| Dose | Cmax | AUC0-t |
|---|---|---|
| 300 mg/kg | 2713 | 22,916 |
| 100 mg/kg | 1055 | 8509 |
| 30 mg/kg | 147 | 1196 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoddart, C.A.; Curreli, F.; Horrigan, S.; Altieri, A.; Kurkin, A.V.; Debnath, A.K. Comparative Pharmacokinetics of a Dual Inhibitor of HIV-1, NBD-14189, in Rats and Dogs with a Proof-of-Concept Evaluation of Antiviral Potency in SCID-hu Mouse Model. Viruses 2022, 14, 2268. https://0-doi-org.brum.beds.ac.uk/10.3390/v14102268
Stoddart CA, Curreli F, Horrigan S, Altieri A, Kurkin AV, Debnath AK. Comparative Pharmacokinetics of a Dual Inhibitor of HIV-1, NBD-14189, in Rats and Dogs with a Proof-of-Concept Evaluation of Antiviral Potency in SCID-hu Mouse Model. Viruses. 2022; 14(10):2268. https://0-doi-org.brum.beds.ac.uk/10.3390/v14102268
Chicago/Turabian StyleStoddart, Cheryl A., Francesca Curreli, Stephen Horrigan, Andrea Altieri, Alexander V. Kurkin, and Asim K. Debnath. 2022. "Comparative Pharmacokinetics of a Dual Inhibitor of HIV-1, NBD-14189, in Rats and Dogs with a Proof-of-Concept Evaluation of Antiviral Potency in SCID-hu Mouse Model" Viruses 14, no. 10: 2268. https://0-doi-org.brum.beds.ac.uk/10.3390/v14102268