
Hyperendemic Dengue and Possible Zika Circulation in the Westernmost Region of the Indonesian Archipelago

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Serum Samples
2.2. Viruses for PRNT
2.3. DENV and ZIKV PRNT
2.4. PRNT Criteria
3. Results
3.1. Participants’ Characteristics
3.2. Prevalence of DENV Neutralizing Antibodies
3.3. DENV Infection Status
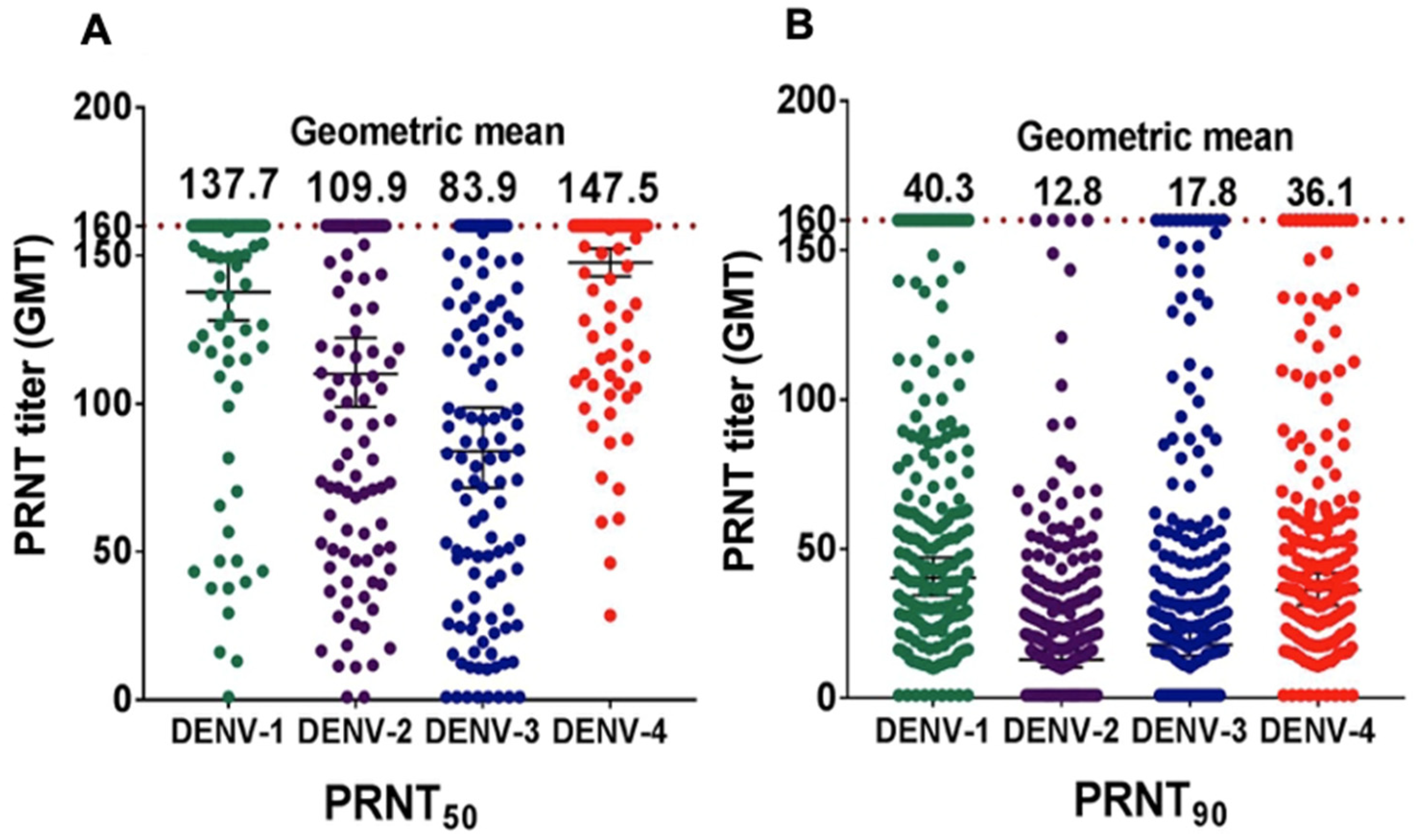
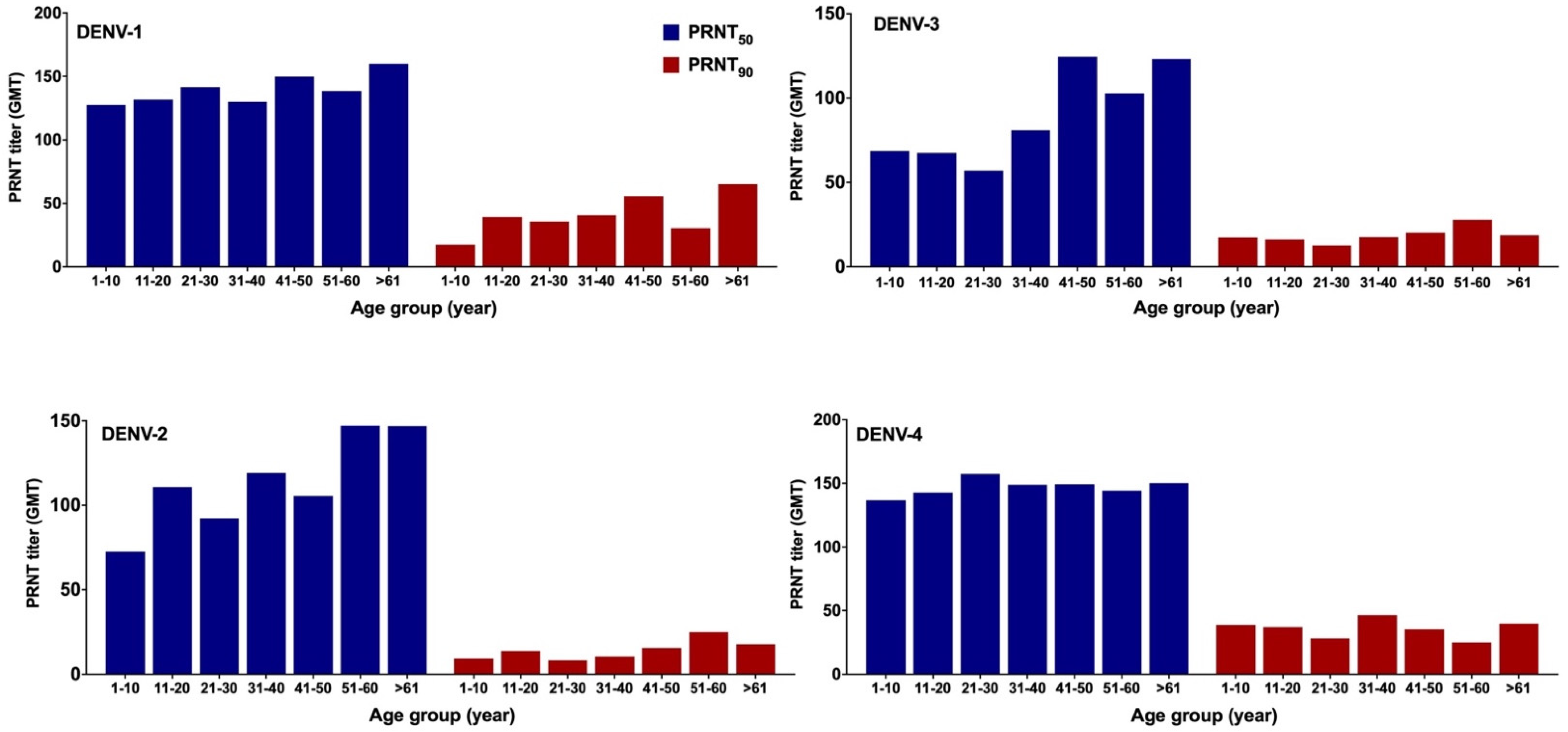
3.4. DENV NAb Geometric Mean Titer (GMT)
3.5. Prevalence of ZIKV NAb
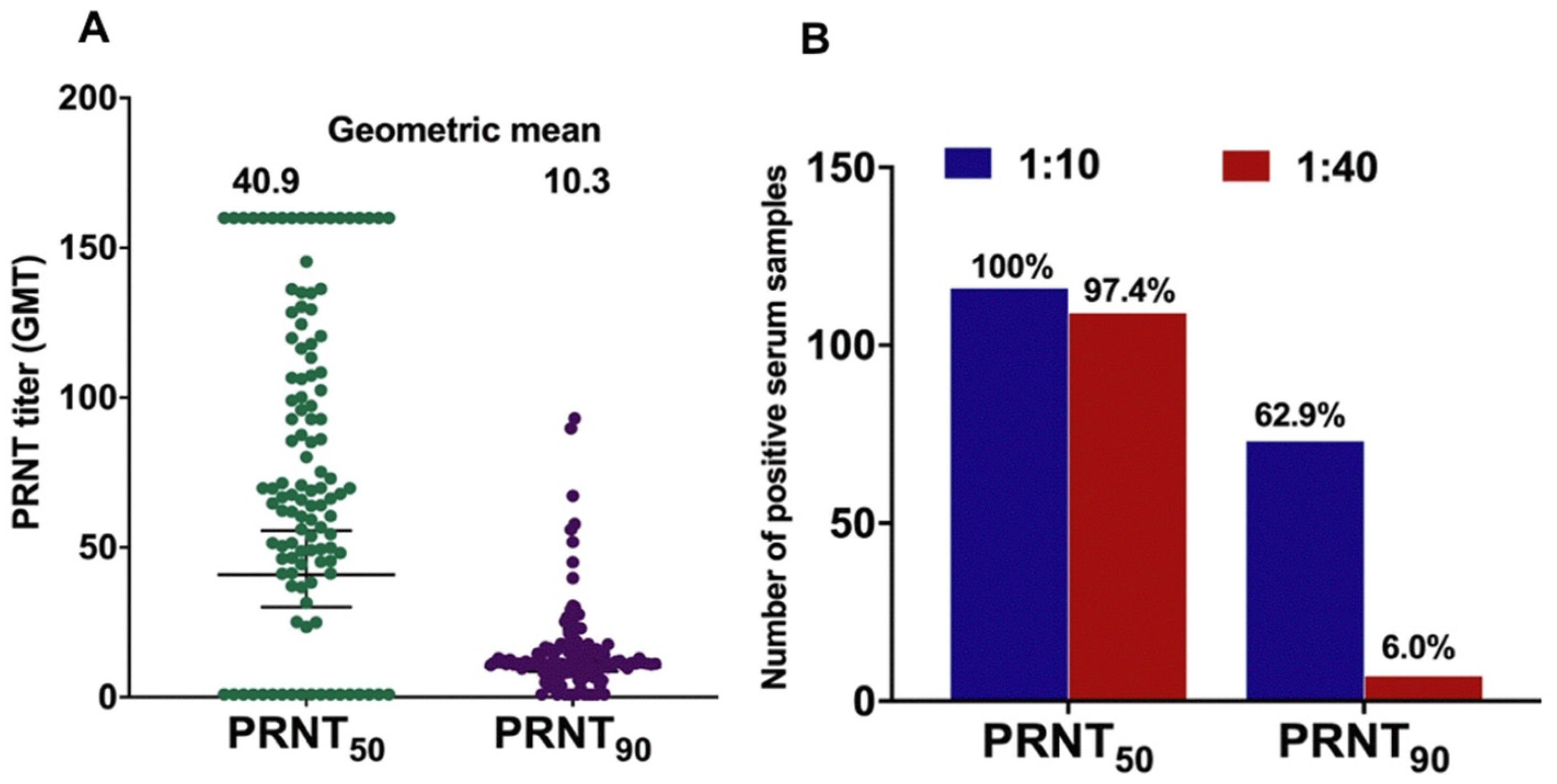
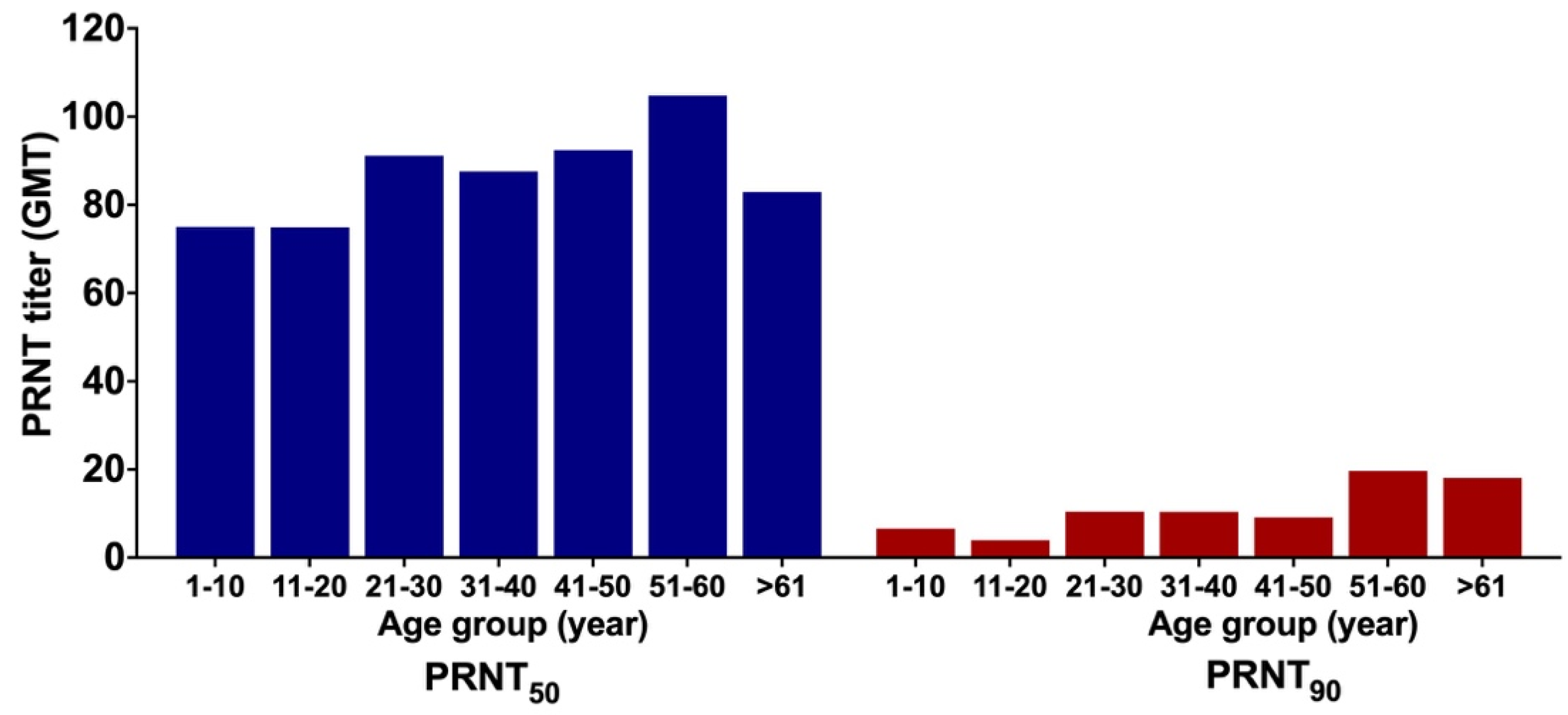
3.6. GMT of ZIKV NAb
3.7. Confirmation of ZIKV Infection
4. Discussion
4.1. Prevalence of DENV-Specific NAb
4.2. Prevalence of ZIKV-Specific NAb
4.3. Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Setiati, T.E. Changing epidemiology of dengue haemorrhagic fever in Indonesia. Dengue Bull. 2006, 30, 1–14. [Google Scholar]
- Dhewantara, P.W.; Jamil, K.F.; Fajar, J.K.; Saktianggi, P.P.; Nusa, R.; Garjito, T.A.; Anwar, S.; Firzan, F.; Megawati, D.; Sasmono, R.T. Decline of notified dengue infections in Indonesia in 2017: Discussion of the possible determinants. Narra J. 2021, 1. [Google Scholar] [CrossRef]
- Harapan, H.; Michie, A.; Yohan, B.; Shu, P.; Mudatsir, M.; Sasmono, R.T.; Imrie, A. Dengue viruses circulating in Indonesia: A systematic review and phylogenetic analysis of data from five decades. Rev. Med Virol. 2019, 29, e2037. [Google Scholar] [CrossRef]
- Megawati, D.; Masyeni, S.; Yohan, B.; Lestarini, A.; Hayati, R.F.; Meutiawati, F.; Suryana, K.; Widarsa, T.; Budiyasa, D.G.; Budiyasa, N.; et al. Dengue in Bali: Clinical characteristics and genetic diversity of circulating dengue viruses. PLoS Negl. Trop. Dis. 2017, 11, e0005483. [Google Scholar] [CrossRef]
- Lestari, C.S.W.; Yohan, B.; Yunita, A.; Meutiawati, F.; Hayati, R.F.; Trimarsanto, H.; Sasmono, R.T. Phylogenetic and evolutionary analyses of dengue viruses isolated in Jakarta, Indonesia. Virus Genes 2017, 53, 778–788. [Google Scholar] [CrossRef]
- Haryanto, S.; Hayati, R.F.; Yohan, B.; Sijabat, L.; Sihite, I.F.; Fahri, S.; Meutiawati, F.; Halim, J.A.N.; Halim, S.N.; Soebandrio, A.; et al. The molecular and clinical features of dengue during outbreak in Jambi, Indonesia in 2015. Pathog. Glob. Health 2016, 110, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Sasmono, R.T.; Wahid, I.; Trimarsanto, H.; Yohan, B.; Wahyuni, S.; Hertanto, M.; Yusuf, I.; Mubin, H.; Ganda, I.J.; Latief, R.; et al. Genomic analysis and growth characteristic of dengue viruses from Makassar, Indonesia. Infect. Genet. Evol. 2015, 32, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Fahri, S.; Yohan, B.; Trimarsanto, H.; Sayono, S.; Hadisaputro, S.; Dharmana, E.; Syafruddin, D.; Sasmono, R.T. Molecular Surveillance of Dengue in Semarang, Indonesia Revealed the Circulation of an Old Genotype of Dengue Virus Serotype-1. PLoS Negl. Trop. Dis. 2013, 7, e2354. [Google Scholar] [CrossRef] [Green Version]
- Nusa, R.; Prasetyowati, H.; Meutiawati, F.; Yohan, B.; Trimarsanto, H.; Setianingsih, T.Y.; Sasmono, R.T. Molecular surveillance of Dengue in Sukabumi, West Java province, Indonesia. J. Infect. Dev. Ctries. 2014, 8, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Wardhani, P.; Aryati, A.; Yohan, B.; Trimarsanto, H.; Setianingsih, T.Y.; Puspitasari, D.; Arfijanto, M.V.; Bramantono, B.; Suharto, S.; Sasmono, R.T. Clinical and virological characteristics of dengue in Surabaya, Indonesia. PLoS ONE 2017, 12, e0178443. [Google Scholar] [CrossRef] [Green Version]
- Kusmintarsih, E.S.; Hayati, R.F.; Turnip, O.N.; Yohan, B.; Suryaningsih, S.; Pratiknyo, H.; Denis, D.; Sasmono, R.T. Molecular characterization of dengue viruses isolated from patients in Central Java, Indonesia. J. Infect. Public Health 2018, 11, 617–625. [Google Scholar] [CrossRef]
- Sasmono, R.T.; Taurel, A.-F.; Prayitno, A.; Sitompul, H.; Yohan, B.; Hayati, R.F.; Bouckenooghe, A.; Hadinegoro, S.R.; Nealon, J. Dengue virus serotype distribution based on serological evidence in pediatric urban population in Indonesia. PLoS Negl. Trop. Dis. 2018, 12, e0006616. [Google Scholar] [CrossRef] [Green Version]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; Gael, C.; et al. Association between Zika virus and microcephaly in French Polynesia, 2013–15: A retrospective study. Lancet 2016, 387, 2125–2132. [Google Scholar] [CrossRef] [Green Version]
- de Araújo, T.V.B.; Rodrigues, L.C.; de Alencar Ximenes, R.A.; de Barros Miranda-Filho, D.; Montarroyos, U.R.; de Melo, A.P.L.; Valongueiro, S.; de Albuquerque, M.D.F.P.M.; Souza, W.V.; Braga, C.; et al. Association between Zika virus infection and microcephaly in Brazil, January to May 2016: Preliminary report of a case-control study. Lancet Infect. Dis. 2016, 16, 1356–1363. [Google Scholar] [CrossRef] [Green Version]
- de Araújo, T.V.B.; Ximenes, R.A.D.A.; Miranda-Filho, D.D.B.; Souza, W.; Montarroyos, U.R.; de Melo, A.P.L.; Valongueiro, S.; Albuquerque, M.D.F.P.M.D.; Braga, C.; Filho, S.B.; et al. Association between microcephaly, Zika virus infection, and other risk factors in Brazil: Final report of a case-control study. Lancet Infect. Dis. 2018, 18, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Schuler-Faccini, L.; Ribeiro, E.M.; Feitosa, I.M.L.; Horovitz, D.D.G.; Cavalcanti, D.P.; Pessoa, A.; Doriqui, M.J.R.; Neri, J.I.; Neto, J.M.D.; Wanderley, H.Y.C.; et al. Possible association between Zika virus Infection and microcephaly—Brazil, 2015. Morbid. Mortal. Wkly. Rep. 2016, 65, 59–62. [Google Scholar] [CrossRef]
- de Oliveira, W.K.; de França, G.V.A.; Carmo, E.H.; Duncan, B.B.; Kuchenbecker, R.D.S.; Schmidt, M.I. Infection-related microcephaly after the 2015 and 2016 Zika virus outbreaks in Brazil: A surveillance-based analysis. Lancet 2017, 390, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Moi, M.L.; Nguyen, T.T.T.; Nguyen, C.T.; Vu, T.B.H.; Tun, M.M.N.; Pham, T.D.; Pham, N.T.; Tran, T.; Morita, K.; Le, T.Q.M.; et al. Zika virus infection and microcephaly in Vietnam. Lancet Infect. Dis. 2017, 17, 805–806. [Google Scholar] [CrossRef] [Green Version]
- Wongsurawat, T.; Athipanyasilp, N.; Jenjaroenpun, P.; Jun, S.-R.; Kaewnapan, B.; Wassenaar, T.M.; Leelahakorn, N.; Angkasekwinai, N.; Kantakamalakul, W.; Ussery, D.W.; et al. Case of Microcephaly after Congenital Infection with Asian Lineage Zika Virus, Thailand. Emerg. Infect. Dis. 2018, 24, 1758–1761. [Google Scholar] [CrossRef]
- Lim, S.K.; Lim, J.K.; Yoon, I.K. An Update on Zika Virus in Asia. Infect. Chemother. 2017, 49, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Marchette, N.J.; Garcia, R.; Rudnick, A. Isolation of Zika Virus from Aedes Aegypti Mosquitoes in Malaysia. Am. J. Trop. Med. Hyg. 1969, 18, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Buathong, R.; Hermann, L.; Thaisomboonsuk, B.; Rutvisuttinunt, W.; Klungthong, C.; Chinnawirotpisan, P.; Manasatienkij, W.; Nisalak, A.; Fernandez, S.; Yoon, I.-K.; et al. Detection of Zika Virus Infection in Thailand, 2012–2014. Am. J. Trop. Med. Hyg. 2015, 93, 380–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alera, M.T.; Hermann, L.; Tac-An, I.A.; Klungthong, C.; Rutvisuttinunt, W.; Manasatienkij, W.; Villa, D.; Thaisomboonsuk, B.; Velasco, J.M.; Chinnawirotpisan, P.; et al. Zika virus infection, Philippines, 2012. Emerg. Infect. Dis. 2015, 21, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.-T.; Ngoc, V.T.N.; Tao, Y. Zika virus infection in Vietnam: Current epidemic, strain origin, spreading risk, and perspective. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2041–2042. [Google Scholar] [CrossRef] [PubMed]
- Heang, V.; Yasuda, C.Y.; Sovann, L.; Haddow, A.D.; Travassos da Rosa, A.P.; Tesh, R.B.; Kasper, M.R. Zika virus infection, Cambodia, 2010. Emerg. Infect. Dis. 2012, 18, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Perkasa, A.; Yudhaputri, F.; Haryanto, S.; Hayati, R.F.; Ma’roef, C.N.; Antonjaya, U.; Yohan, B.; Myint, K.S.A.; Ledermann, J.P.; Rosenberg, R.; et al. Isolation of Zika Virus from Febrile Patient, Indonesia. Emerg. Infect. Dis. 2016, 22, 924–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffree, S.; Dony, J.J.F.; Ahmed, K. The first outbreak of autochthonous Zika virus in Sabah, Malaysian Borneo. Int. J. Infect. Dis. 2018, 73, 213. [Google Scholar] [CrossRef]
- Duong, V.; Dussart, P.; Buchy, P. Zika virus in Asia. Int. J. Infect. Dis. 2017, 54, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Singapore Zika Study Group. Outbreak of Zika virus infection in Singapore: An epidemiological, entomological, virological, and clinical analysis. Lancet Infect. Dis. 2017, 17, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Musso, D.; Lanteri, M.C. Zika virus in Singapore: Unanswered questions. Lancet Infect. Dis. 2017, 17, 782–783. [Google Scholar] [CrossRef] [Green Version]
- Gordon, A.; Gresh, L.; Ojeda, S.; Katzelnick, L.C.; Sanchez, N.; Mercado, J.C.; Chowell, G.; Lopez, B.; Elizondo, D.; Coloma, J.; et al. Prior dengue virus infection and risk of Zika: A pediatric cohort in Nicaragua. PLoS Med. 2019, 16, e1002726. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.G.; Ksiazek, T.G.; Gubler, D.; Lubis, S.I.; Simanjuntak, G.; Lee, V.H.; Nalim, S.; Juslis, K.; See, R. A survey for arboviral antibodies in sera of humans and animals in Lombok, Republic of Indonesia. Ann. Trop. Med. Parasitol. 1983, 77, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.G.; Ksiazek, T.G.; Suhandiman; Triwibowo. Zika virus, a cause of fever in Central Java, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 389–393. [Google Scholar] [CrossRef]
- Sasmono, R.T.; Dhenni, R.; Yohan, B.; Pronyk, P.; Hadinegoro, S.R.; Soepardi, E.J.; Ma’roef, C.N.; Satari, H.I.; Menzies, H.; Hawley, W.A.; et al. Zika Virus Seropositivity in 1–4-Year-Old Children, Indonesia, 2014. Emerg. Infect. Dis. 2018, 24, 1740. [Google Scholar] [CrossRef]
- Kwong, J.C.; Druce, J.D.; Leder, K. Case report: Zika virus infection acquired during brief travel to Indonesia. Am. J. Trop. Med. Hyg. 2013, 89, 516–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, G.H.Y.; Baird, R.; Druce, J.; Anstey, N.M. Zika virus infection in australia following a monkey bite in indonesia. Southeast Asian J. Trop. Med. Public Health 2015, 46, 460. [Google Scholar] [PubMed]
- Bogoch, I.I.; Brady, O.J.; Kraemer, M.U.G.; German, M.; Creatore, I.M.; Brent, S.; Watts, A.G.; Hay, S.; A Kulkarni, M.; Brownstein, J.S.; et al. Potential for Zika virus introduction and transmission in resource-limited countries in Africa and the Asia-Pacific region: A modelling study. Lancet Infect. Dis. 2016, 16, 1237–1245. [Google Scholar] [CrossRef] [Green Version]
- Sasmono, R.T.; Johar, E.; Yohan, B.; Ma’roef, C.N.; Pronyk, P.; Hadinegoro, S.R.; Soepardi, E.J.; Bouckenooghe, A.; Hawley, W.A.; Rosenberg, R.; et al. Spatiotemporal Heterogeneity of Zika Virus Transmission in Indonesia: Serosurveillance Data from a Pediatric Population. Am. J. Trop. Med. Hyg. 2021, 104, 2220–2223. [Google Scholar] [CrossRef]
- Badan Pusat Statistik. Proyeksi Jumlah Penduduk Kabupaten Aceh Jaya (Jiwa), 2017–2019; Badan Pusat Statistik Kabupaten Aceh Jaya: Jakarta, Indonesia, 2019. [Google Scholar]
- Ernst, T.; McCarthy, S.; Chidlow, G.; Luang-Suarkia, D.; Holmes, E.C.; Smith, D.W.; Imrie, A. Emergence of a New Lineage of Dengue Virus Type 2 Identified in Travelers Entering Western Australia from Indonesia, 2010–2012. PLoS Negl. Trop. Dis. 2015, 9, e0003442. [Google Scholar] [CrossRef]
- WHO. Guidelines for Plaque Reduction Neutralization Testing of Human Antibodies to Dengue Viruses; WHO: Geneva, Switzerland, 2008; Available online: http://whqlibdoc.who.int/hq/2007/who_ivb_07.07_eng.pdf (accessed on 29 August 2018).
- Timiryasova, T.M.; Bonaparte, M.I.; Luo, P.; Zedar, R.; Hu, B.T.; Hildreth, S.W. Optimization and Validation of a Plaque Reduction Neutralization Test for the Detection of Neutralizing Antibodies to Four Serotypes of Dengue Virus Used in Support of Dengue Vaccine Development. Am. J. Trop. Med. Hyg. 2013, 88, 962–970. [Google Scholar] [CrossRef]
- Panta, K. Antibody Mediated Immune Response Againts Flavivirus Infection in Western Australia Travellers; The University of Western Australia: Perth, Australia, 2018. [Google Scholar]
- Garg, S.; Chakravarti, A.; Singh, R.; Masthi, N.R.; Goyal, R.C.; Jammy, G.R.; Ganguly, E.; Sharma, N.; Singh, M.M.; Ferreira, G.; et al. Dengue serotype-specific seroprevalence among 5- to 10-year-old children in India: A community-based cross-sectional study. Int. J. Infect. Dis. 2017, 54, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeewandara, C.; Gomes, L.; Paranavitane, S.A.; Tantirimudalige, M.; Panapitiya, S.S.; Jayewardene, A.; Fernando, S.; Fernando, R.H.; Prathapan, S.; Ogg, G.S.; et al. Change in Dengue and Japanese Encephalitis Seroprevalence Rates in Sri Lanka. PLoS ONE 2015, 10, e0144799. [Google Scholar] [CrossRef] [Green Version]
- Thai, K.T.D.; Binh, T.Q.; Giao, P.T.; Phuöng, H.L.; Hung, L.Q.; Van Nam, N.; Nga, T.T.; Groen, J.; Nagelkerke, N.; Vries, P.J. Seroprevalence of dengue antibodies, annual incidence and risk factors among children in southern Vietnam. Trop. Med. Int. Health 2005, 10, 379–386. [Google Scholar] [CrossRef]
- Amaya-Larios, I.Y.; Ramos-Castañeda, J.; Sepúlveda-Salinas, K.J.; Mayer, S.V.; Falcón-Lezama, J.A.; Galeana-Hernández, M.; Vasilakis, N.; Comas-García, A.; Martínez-Vega, R.A. Seroprevalence of Neutralizing Antibodies Against Dengue Virus in Two Localities in the State of Morelos, Mexico. Am. J. Trop. Med. Hyg. 2014, 91, 1057–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortuna, C.; Remoli, M.E.; Rizzo, C.; Benedetti, E.; Fiorentini, C.; Bella, A.; Argentini, C.; Farchi, F.; Castilletti, C.; Capobianchi, M.R.; et al. Imported arboviral infections in Italy, July 2014–October 2015: A National Reference Laboratory report. BMC Infect. Dis. 2017, 17, 216. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Lee, J.; Kim, Y.-E.; Chong, C.-K.; Pinchemel, Y.; Reisdörfer, F.; Coelho, J.B.; Dias, R.F.; Bae, P.K.; Gusmão, Z.P.M.; et al. Development of a Rapid Diagnostic Test Kit to Detect IgG/IgM Antibody against Zika Virus Using Monoclonal Antibodies to the Envelope and Non-structural Protein 1 of the Virus. Korean J. Parasitol. 2018, 56, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Chua, A.; Prat, I.; Nuebling, C.M.; Wood, D.; Moussy, F. Update on Zika Diagnostic Tests and WHO’s Related Activities. PLoS Negl. Trop. Dis. 2017, 11, e0005269. [Google Scholar] [CrossRef]
- Roehrig, J.T.; Hombach, J.; Barrett, A.D.T. Guidelines for Plaque-Reduction Neutralization Testing of Human Antibodies to Dengue Viruses. Viral Immunol. 2008, 21, 123–132. [Google Scholar] [CrossRef]
- Shan, C.; Xie, X.; Ren, P.; Loeffelholz, M.J.; Yang, Y.; Furuya, A.; Dupuis, A.P., 2nd; Kramer, L.D.; Wong, S.J.; Shi, P.Y. A Rapid Zika Diagnostic Assay to Measure Neutralizing Antibodies in Patients. EBioMedicine 2017, 17, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.T.; Wong, S.J.; Kulas, K.E.; Dupuis, A.P., 2nd; Payne, A.F.; Kramer, L.D.; Dean, A.B.; St George, K.; White, J.L.; Sommer, J.N.; et al. Development of Zika Virus Serological Testing Strategies in New York State. J. Clin. Microbiol. 2018, 56, e01591-17. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M.; Khouri, R.; Sousa, P.; Branco, M.; Batista, R.; Costa, E.; Alves, M.; Amaral, G.; Borges, M.; Takahasi, E.; et al. Plaque Reduction Neutralization Test (PRNT) in the Congenital Zika Syndrome: Positivity and Associations with Laboratory, Clinical, and Imaging Characteristics. Viruses 2020, 12, 1244. [Google Scholar] [CrossRef] [PubMed]
- Koishi, A.C.; Suzukawa, A.A.; Zanluca, C.; Camacho, D.E.; Comach, G.; Dos Santos, C.N.D. Development and evaluation of a novel high-throughput image-based fluorescent neutralization test for detection of Zika virus infection. PLoS Negl. Trop. Dis. 2018, 12, e0006342. [Google Scholar] [CrossRef] [PubMed]
- L’Huillier, A.G.; Hamid-Allie, A.; Kristjanson, E.; Papageorgiou, L.; Hung, S.; Wong, C.F.; Stein, D.R.; Olsha, R.; Goneau, L.W.; Dimitrova, K.; et al. Evaluation of Euroimmun Anti-Zika Virus IgM and IgG Enzyme-Linked Immunosorbent Assays for Zika Virus Serologic Testing. J. Clin. Microbiol. 2017, 55, 2462–2471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, I.; Martin, S.W.; Fischer, M.; Chambers, T.V.; Kosoy, O.; Al, I.G.E.; Ponomareva, O.; Gillis, L.D.; Blackmore, C.; Jean, R. Zika Virus IgM Detection and Neutralizing Antibody Profiles 12–19 Months after Illness Onset. Emerg. Infect. Dis. 2019, 25, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.; Onlamoon, N.; Sewatanon, J.; Edupuganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.J.; Wilson, P.C.; et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus. Proc. Natl. Acad. Sci. USA 2016, 113, 7852–7857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.A.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, L.M.; Carlin, E.R.; Jenkins, M.M.; Tan, A.L.; Barcellona, C.M.; Nicholson, C.O.; Michael, S.F.; Isern, S. Dengue virus antibodies enhance Zika virus infection. Clin. Transl. Immunol. 2016, 5, e117. [Google Scholar] [CrossRef] [PubMed]
- Castanha, P.; Nascimento, E.J.M.; Cynthia, B.; Cordeiro, M.T.; De Carvalho, O.V.; De Mendonça, L.R.; Azevedo, E.; França, R.F.; Rafael, D.; Marques, E.T. Dengue virus (DENV)-specific antibodies enhance Brazilian Zika virus (ZIKV) infection. J. Infect. Dis. 2016, 215, 781–785. [Google Scholar] [CrossRef] [Green Version]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.-M.; Malasit, P.; Rey, F.A.; et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef]
- Swanstrom, J.A.; Plante, J.A.; Plante, K.S.; Young, E.F.; McGowan, E.; Gallichotte, E.N.; Widman, D.G.; Heise, M.T.; de Silva, A.M.; Baric, R.S. Dengue Virus Envelope Dimer Epitope Monoclonal Antibodies Isolated from Dengue Patients Are Protective against Zika Virus. mBio 2016, 7, e01123-16. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, G.S.; Kikuti, M.; Tauro, L.B.; Nascimento, L.C.J.; Cardoso, C.; Campos, G.S.; Ko, A.; Weaver, S.C.; Reis, M.G.; Kitron, U.; et al. Does immunity after Zika virus infection cross-protect against dengue? Lancet Glob. Health 2018, 6, e140–e141. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Elong Ngono, A.; Regla-Nava, J.A.; Kim, K.; Gorman, M.J.; Diamond, M.S.; Shresta, S. Dengue virus-reactive CD8+ T cells mediate cross-protection against subsequent Zika virus challenge. Nat. Commun. 2017, 8, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantoja, P.; Pérez-Guzmán, E.X.; Rodríguez, I.V.; White, L.J.; González, O.; Serrano, C.; Giavedoni, L.; Hodara, V.; Cruz, L.; Arana, T.; et al. Zika virus pathogenesis in rhesus macaques is unaffected by pre-existing immunity to dengue virus. Nat. Commun. 2017, 8, 15674. [Google Scholar] [CrossRef] [PubMed]
- McCracken, M.; Gromowski, G.D.; Friberg, H.L.; Lin, X.; Abbink, P.; De La Barrera, R.; Eckles, K.H.; Garver, L.S.; Boyd, M.; Jetton, D.; et al. Impact of prior flavivirus immunity on Zika virus infection in rhesus macaques. PLoS Pathog. 2017, 13, e1006487. [Google Scholar] [CrossRef] [PubMed]
- Terzian, A.C.B.; Schanoski, A.S.; Mota, M.T.D.O.; Da Silva, R.A.; Estofolete, C.F.; Colombo, T.E.; Rahal, P.; Hanley, K.A.; Vasilakis, N.; Kalil, J.; et al. Viral Load and Cytokine Response Profile Does Not Support Antibody-Dependent Enhancement in Dengue-Primed Zika Virus–Infected Patients. Clin. Infect. Dis. 2017, 65, 1260–1265. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Threshold and Titer Cut-Off | DENV-1 n (%) | DENV-2 n (%) | DENV-3 n (%) | DENV-4 n (%) |
|---|---|---|---|---|
| PRNT50 | ||||
| 10 | 198 (99.5) | 197 (99.0) | 191 (96.0) | 199 (100.0) |
| 40 | 192 (96.5) | 181 (91.0) | 168 (84.4) | 198 (99.5) |
| PRNT90 | ||||
| 10 | 190 (95.5) | 151 (75.9) | 156 (78.4) | 190 (95.5) |
| 40 | 111 (55.8) | 45 (22.6) | 68 (34.2) | 97 (48.7) |
| ID | Age (Year) | PRNT90 Titer for DENV | Status of Dengue Infection | ZIKV PRNT90 Titer | Infection Status * | ||||
|---|---|---|---|---|---|---|---|---|---|
| DENV-1 | DENV-2 | DENV-3 | DENV-4 | PRNT90 > 10 | PRNT90 > 40 | ||||
| A-157 | 53 | 100.05 | 120.75 | >160 | >160 | Multitypic | Multitypic | 93.119 | Flavivirus |
| A-52 | 63 | 39.19 | 11.55 | 36.86 | 100.23 | Multitypic | DENV-4 | 89.746 | Flavivirus |
| A-97 | 60 | 104.86 | 43.27 | 57.94 | 117.66 | Multitypic | Multitypic | 67.247 | Flavivirus |
| A-71 | 53 | 32.43 | 16.61 | 31.40 | 36.67 | Multitypic | Naïve | 57.835 | Flavivirus |
| A-119 | 22 | 27.48 | 36.11 | 29.55 | 39.67 | Multitypic | Naïve | 55.986 | Flavivirus |
| A-94 | 50 | >160 | 24.31 | 28.42 | 89.72 | Multitypic | Multitypic | 51.872 | Flavivirus |
| A-13 | 55 | 48.83 | 24.40 | 15.89 | 160.00 | Multitypic | DENV-4 | 45.076 | Flavivirus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harapan, H.; Panta, K.; Michie, A.; Ernst, T.; McCarthy, S.; Muhsin, M.; Safarianti, S.; Zanaria, T.M.; Mudatsir, M.; Sasmono, R.T.; et al. Hyperendemic Dengue and Possible Zika Circulation in the Westernmost Region of the Indonesian Archipelago. Viruses 2022, 14, 219. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020219
Harapan H, Panta K, Michie A, Ernst T, McCarthy S, Muhsin M, Safarianti S, Zanaria TM, Mudatsir M, Sasmono RT, et al. Hyperendemic Dengue and Possible Zika Circulation in the Westernmost Region of the Indonesian Archipelago. Viruses. 2022; 14(2):219. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020219
Chicago/Turabian StyleHarapan, Harapan, Kritu Panta, Alice Michie, Timo Ernst, Suzi McCarthy, Muhsin Muhsin, Safarianti Safarianti, Tjut Mariam Zanaria, Mudatsir Mudatsir, R. Tedjo Sasmono, and et al. 2022. "Hyperendemic Dengue and Possible Zika Circulation in the Westernmost Region of the Indonesian Archipelago" Viruses 14, no. 2: 219. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020219