
Characterization of the Novel Phage vB_VpaP_FE11 and Its Potential Role in Controlling Vibrio parahaemolyticus Biofilms

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Phage Isolation and Purification
2.3. Transmission Electron Microscopy (TEM) Analysis
2.4. Phage Host Range
2.5. Isolation, Genome Sequencing, and Assembly of Phage DNA
2.6. Genome Analysis and Phylogenetic Analysis
2.7. One-Step Growth Experiment
2.8. Effects of Temperature, pH, and Ultraviolet (UV) Irradiation on Phage Activity
2.9. Effect of Phages on the Biofilm Formed
2.10. Effect of Phages on the Formation of Biofilm
2.11. Scanning Electron Microscopy (SEM) Analysis
2.12. Statistical Analysis
2.13. Accession Number
3. Results
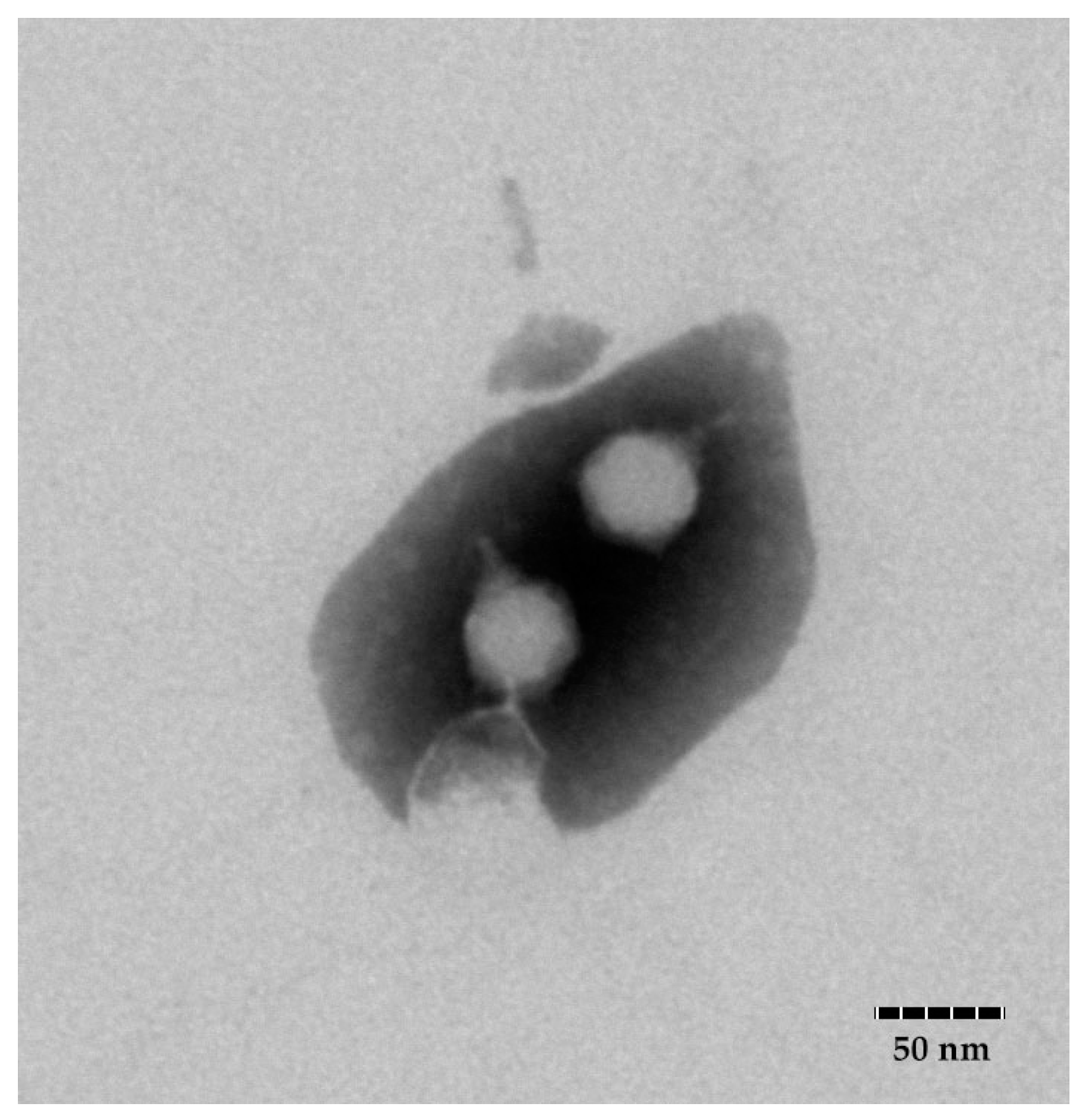
3.1. Isolation and Morphological Characterization of Phage FE11
3.2. Host Range of Phage FE11
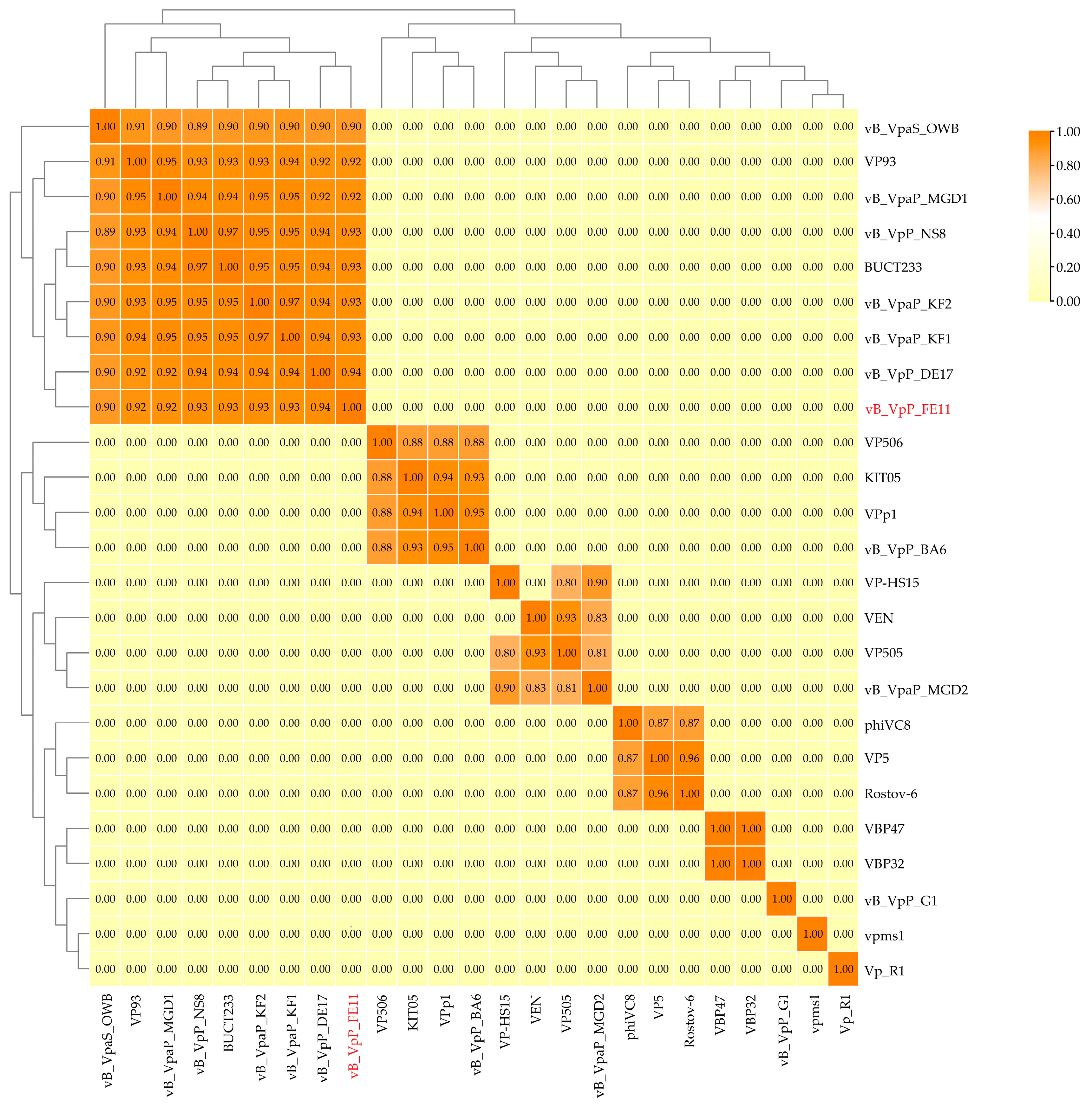
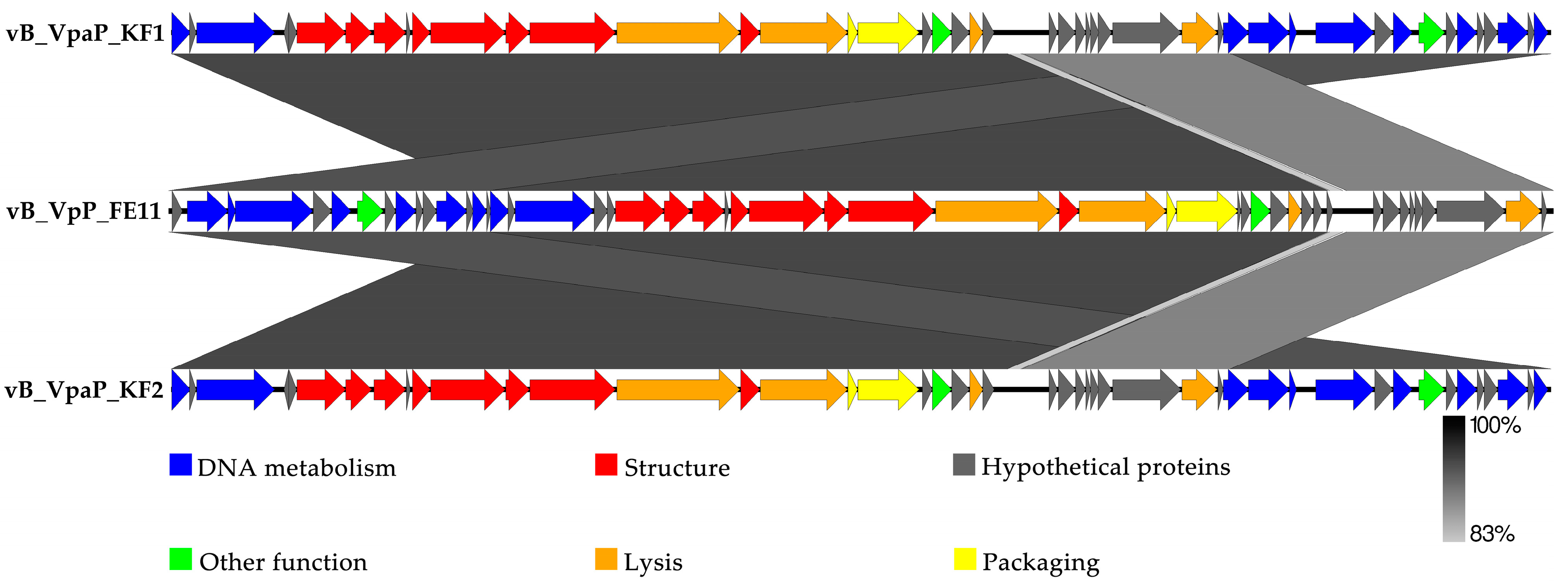
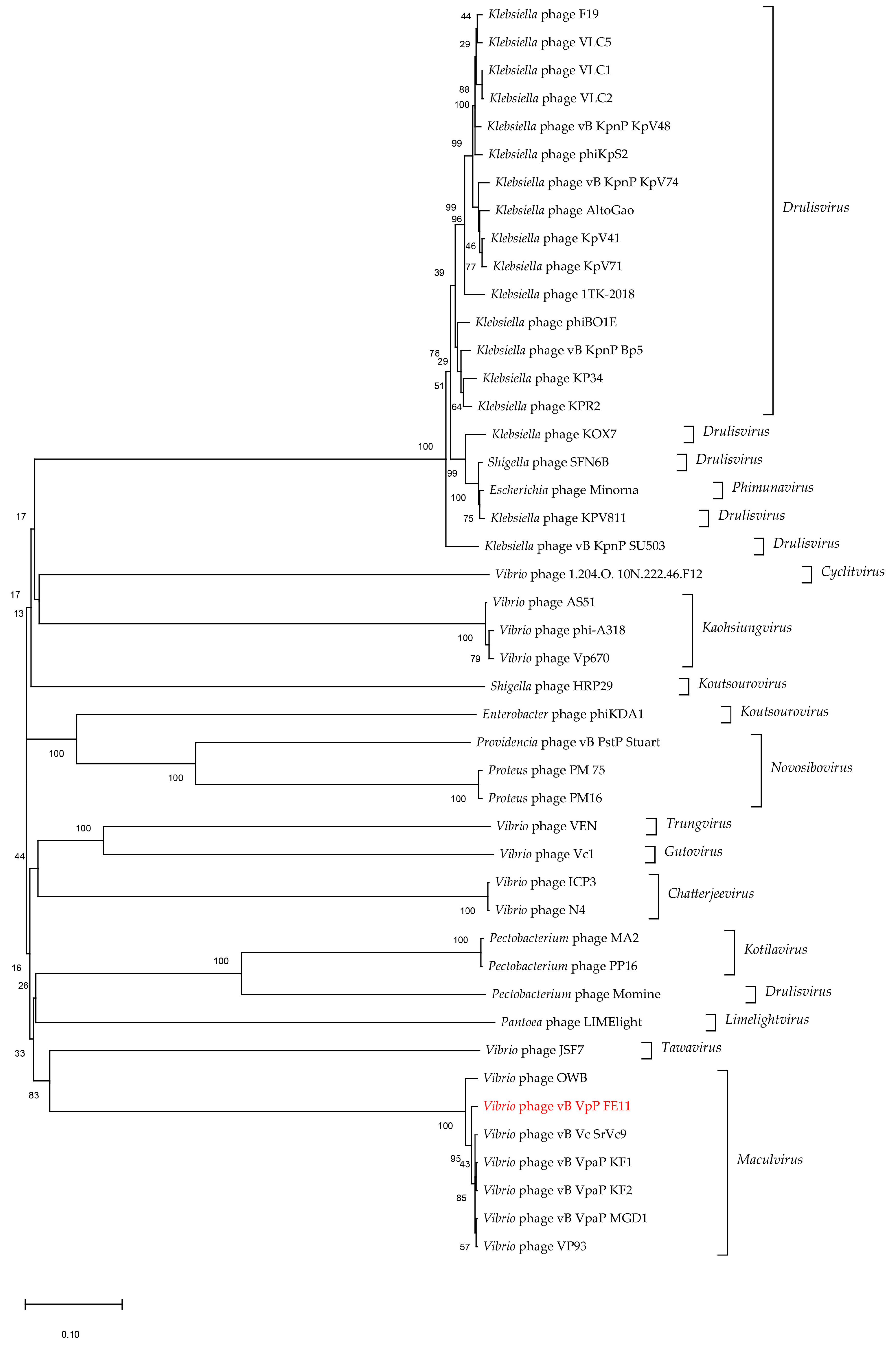
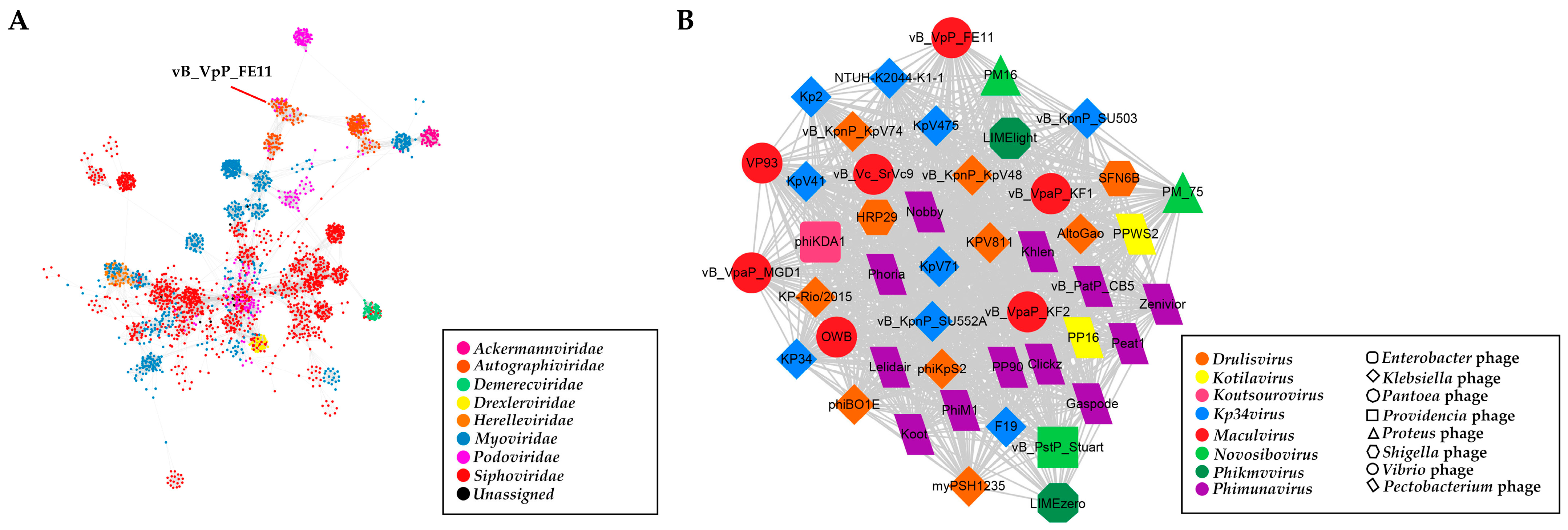
3.3. The Genome Analysis of FE11
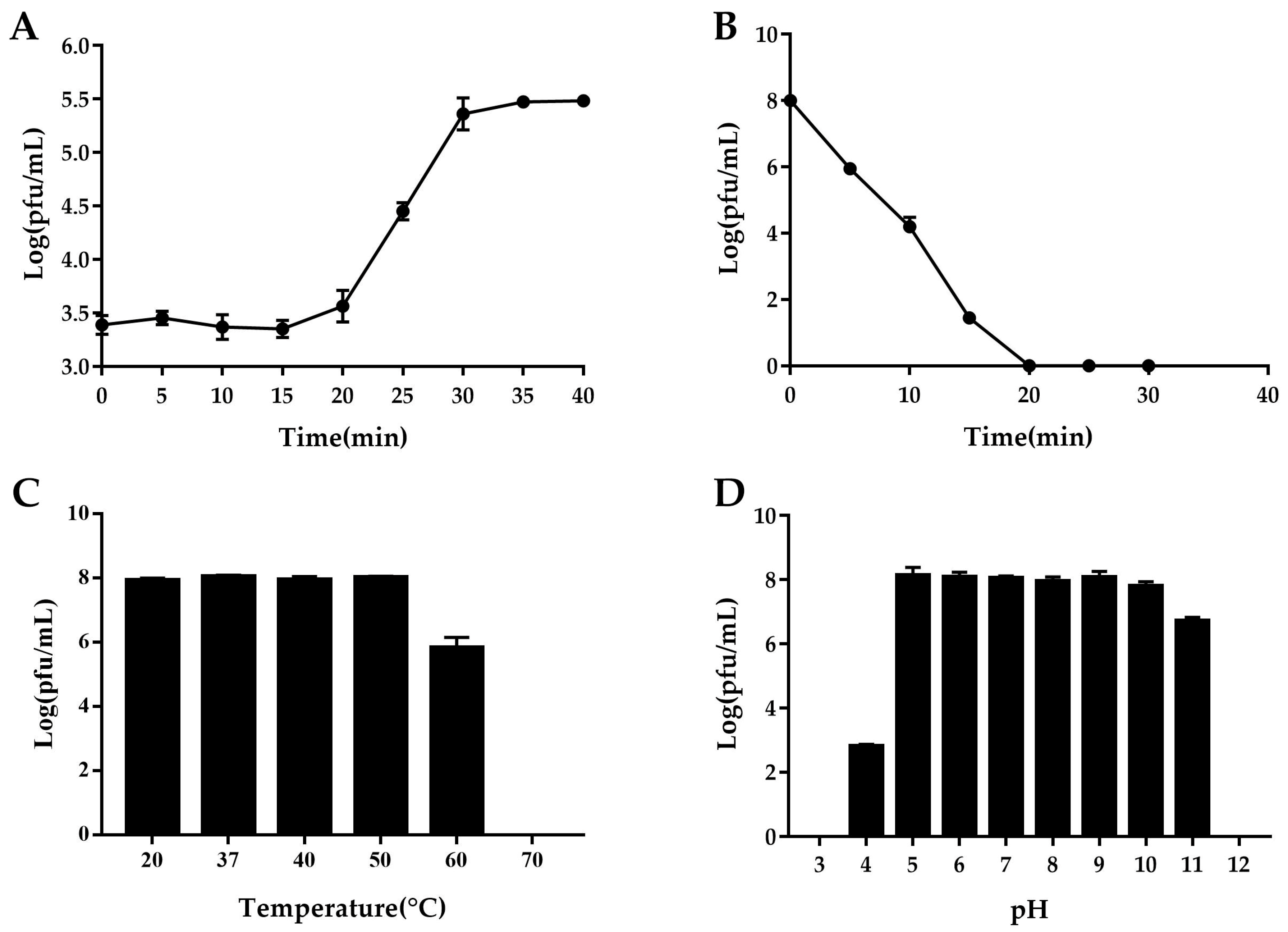
3.4. One-Step Growth Curve
3.5. Determination of Phage Stability
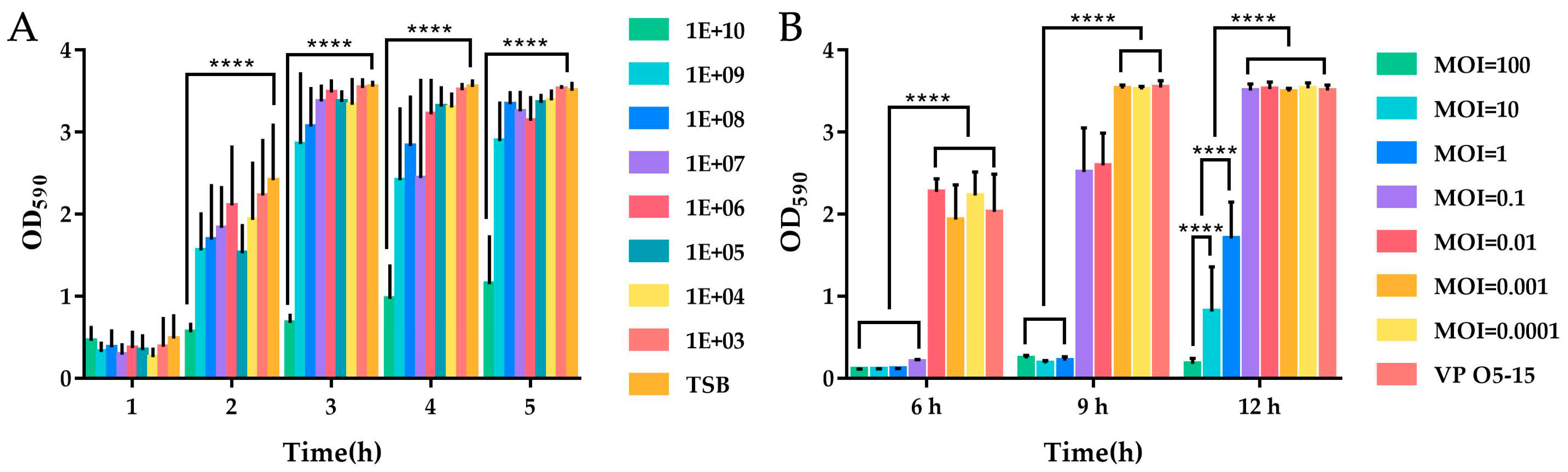
3.6. Effect of Phage on Formed Biofilm
3.7. Effect of Phage on Biofilm Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matsuda, S.; Hiyoshi, H.; Tandhavanant, S.; Kodama, T. Advances on Vibrio Parahaemolyticus Research in the Postgenomic Era. Microbiol. Immunol. 2020, 64, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Spaur, M.; Davis, B.J.K.; Kivitz, S.; DePaola, A.; Bowers, J.C.; Curriero, F.C.; Nachman, K.E. A Systematic Review of Post-Harvest Interventions for Vibrio Parahaemolyticus in Raw Oysters. Sci. Total Environ. 2020, 745, 140795. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Hsu, K.; Wang, H. Structural Insights into the Cytotoxic Mechanism of Vibrio Parahaemolyticus PirA(vp) and PirB(vp) Toxins. Mar. Drugs 2017, 15, 373. [Google Scholar] [CrossRef] [Green Version]
- Nair, G.B.; Ramamurthy, T.; Bhattacharya, S.K.; Dutta, B.; Takeda, Y.; Sack, D.A. Global Dissemination of Vibrio Parahaemolyticus Serotype O3: K6 and its Serovariants. Clin. Microbiol. Rev. 2007, 20, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, X.; Lu, L.; Xu, D. Progress in Research on Acute Hepatopancreatic Necrosis Disease (AHPND). Aquacult. Int. 2016, 24, 577–593. [Google Scholar] [CrossRef]
- Song, X.; Ma, Y.; Fu, J.; Zhao, A.; Guo, Z.; Malakar, P.K.; Pan, Y.; Zhao, Y. Effect of Temperature on Pathogenic and Non-Pathogenic Vibrio Parahaemolyticus Biofilm Formation. Food Control 2017, 73, 485–491. [Google Scholar] [CrossRef]
- Elexson, N.; Afsah-Hejri, L.; Rukayadi, Y.; Soopna, P.; Lee, H.Y.; Zainazor, T.C.T.; Ainy, M.N.; Nakaguchi, Y.; Mitsuaki, N.; Son, R. Effect of Detergents as Antibacterial Agents on Biofilm of Antibiotics-Resistant Vibrio Parahaemolyticus Isolates. Food Control 2014, 35, 378–385. [Google Scholar] [CrossRef]
- Xie, T.; Wu, Q.; Xu, X.; Zhang, J.; Guo, W. Prevalence and Population Analysis of Vibrio Parahaemolyticus in Aquatic Products from South China Markets. FEMS Microbiol. Lett. 2015, 362, fnv178. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Xu, X.; Wu, Q.; Zhang, J.; Cheng, J. Prevalence, Molecular Characterization, and Antibiotic Susceptibility of Vibrio Parahaemolyticus from Ready-to-Eat Foods in China. Front. Microbiol. 2016, 7, 549. [Google Scholar] [CrossRef]
- Hendrix, R.W. Bacteriophages: Evolution of the Majority. Theor. Popul. Biol. 2002, 61, 471–480. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, Z.; Li, Z.; Wang, L.; Li, H.; Wu, F.; Jin, L.; Li, X.; Li, S.; Xu, Y. Effect of Bacteriophages on Vibrio Alginolyticus Infection in the Sea Cucumber, Apostichopus Japonicus (Selenka). J. World Aquacult. Soc. 2015, 46, 149–158. [Google Scholar] [CrossRef]
- Li, Z.; Ren, H.; Li, Q.; Murtaza, B.; Li, X.; Zhang, J.; Xu, Y. Exploring the Effects of Phage Cocktails in Preventing Vibrio Infections in Juvenile Sea Cucumber (Apostichopus Japonicus) Farming. Aquaculture 2020, 515, 734599. [Google Scholar] [CrossRef]
- Higuera, G.; Bastias, R.; Tsertsvadze, G.; Romero, J.; Espejo, R.T. Recently Discovered Vibrio Anguillarum Phages can Protect Against Experimentally Induced Vibriosis in Atlantic Salmon, Salmo Salar. Aquaculture 2013, 392, 128–133. [Google Scholar] [CrossRef]
- Kim, S.G.; Jun, J.W.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.W.; Kang, J.W.; Han, S.J.; Jeong, D.; Park, S.C. Isolation and Characterisation of pVa-21, a Giant Bacteriophage with Anti-Biofilm Potential Against Vibrio Alginolyticus. Sci. Rep. 2019, 9, 6284. [Google Scholar] [CrossRef]
- Sasikala, D.; Srinivasan, P. Characterization of Potential Lytic Bacteriophage Against Vibrio Alginolyticus and its Therapeutic Implications on Biofilm Dispersal. Microb. Pathog. 2016, 101, 24–35. [Google Scholar] [CrossRef]
- Van Twest, R.; Kropinski, A.M. Bacteriophage Enrichment from Water and Soil. Methods Mol. Biol. 2009, 501, 15–21. [Google Scholar] [CrossRef]
- Ajuebor, J.; Buttimer, C.; Arroyo-Moreno, S.; Chanishvili, N.; Gabriel, E.M.; O’Mahony, J.; McAuliffe, O.; Neve, H.; Franz, C.; Coffey, A. Comparison of Staphylococcus Phage K with Close Phage Relatives Commonly Employed in Phage Therapeutics. Antibiotics 2018, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, P. Purification of Bacteriophages and SDS-PAGE Analysis of Phage Structural Proteins from Ghost Particles. Methods Mol. Biol. 2009, 502, 227–238. [Google Scholar] [CrossRef]
- Bian, S.; Jia, Y.; Zhan, Q.; Wong, N.; Hu, Q.; Zhang, W.; Zhang, Y.; Li, L. VPsero: Rapid Serotyping of Vibrio Parahaemolyticus Using Serogroup-Specific Genes Based on Whole-Genome Sequencing Data. Front. Microbiol. 2021, 12, 2446. [Google Scholar] [CrossRef]
- Lu, S.; Le, S.; Tan, Y.; Zhu, J.; Li, M.; Rao, X.; Zou, L.; Li, S.; Wang, J.; Jin, X.; et al. Genomic and Proteomic Analyses of the Terminally Redundant Genome of the Pseudomonas aeruginosa Phage PaP1: Establishment of Genus PaP1-Like Phages. PLoS ONE 2013, 8, e62933. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, M.; Karimi Torshizi, M.A.; Rahimi, S.; Dennehy, J.J. Prophylactic Bacteriophage Administration More Effective than Post-infection Administration in Reducing Salmonella Enterica Serovar Enteritidis Shedding in Quail. Front. Microbiol. 2016, 7, 1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garneau, J.R.; Depardieu, F.; Fortier, L.; Bikard, D.; Monot, M. PhageTerm: A Tool for Fast and Accurate Determination of Phage Termini and Packaging Mechanism Using Next-Generation Sequencing Data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and Taxonomy in Diagnostics for Food Security: Soft-Rotting Enterobacterial Plant Pathogens. Anal. Methods 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.H.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.; Chang, H.; Daugherty, L.; Fraser, M.; Hunter, S.; Lopez, R.; McAnulla, C.; McMenamin, C.; Nuka, G.; Pesseat, S.; et al. The InterPro Protein Families Database: The Classification Resource After 15 Years. Nucleic Acids Res. 2015, 43, D213–D221. [Google Scholar] [CrossRef]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred Interactive Server for Protein Homology Detection and Structure Prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis Across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bolduc, B.; Jang, H.B.; Doulcier, G.; You, Z.; Roux, S.; Sullivan, M.B. VConTACT: An iVirus Tool to Classify Double-Stranded DNA Viruses that Infect Archaea and Bacteria. PeerJ 2017, 5, e3243. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M. Practical Advice on the One-Step Growth Curve. Methods Mol. Biol. 2018, 1681, 41–47. [Google Scholar] [CrossRef]
- Yang, M.; Liang, Y.; Huang, S.; Zhang, J.; Wang, J.; Chen, H.; Ye, Y.; Gao, X.; Wu, Q.; Tan, Z. Isolation and Characterization of the Novel Phages vB_VpS_BA3 and vB_VpS_CA8 for Lysing Vibrio parahaemolyticus. Front. Microbiol. 2020, 11, 259. [Google Scholar] [CrossRef]
- Imam, M.; Alrashid, B.; Patel, F.; Dowah, A.S.A.; Brown, N.; Millard, A.; Clokie, M.R.J.; Galyov, E.E. VB_PaeM_MIJ3, a Novel Jumbo Phage Infecting Pseudomonas aeruginosa, Possesses Unusual Genomic Features. Front. Microbiol. 2019, 10, 2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endersen, L.; Buttimer, C.; Nevin, E.; Coffey, A.; Neve, H.; Oliveira, H.; Lavigne, R.; O’Mahony, J. Investigating the Biocontrol and Anti-Biofilm Potential of a Three Phage Cocktail Against Cronobacter Sakazakii in Different Brands of Infant Formula. Int. J. Food Microbiol. 2017, 253, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knezevic, P.; Petrovic, O. A Colorimetric Microtiter Plate Method for Assessment of Phage Effect on Pseudomonas Aeruginosa Biofilm. J. Microbiol. Meth. 2008, 74, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Qu, K.; Tan, D.; Li, X.; Wan, L.; Cong, C.; Xiu, Z.; Xu, Y. Isolation and Characterization of a Bacteriophage and its Potential to Disrupt Multi-Drug Resistant Pseudomonas Aeruginosa Biofilms. Microb. Pathog. 2019, 128, 329–336. [Google Scholar] [CrossRef]
- Adriaenssens, E.M.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Lomeli-Ortega, C.O.; Martinez-Diaz, S.F. Phage Therapy Against Vibrio Parahaemolyticus Infection in the Whiteleg Shrimp (Litopenaeus Vannamei) Larvae. Aquaculture 2014, 434, 208–211. [Google Scholar] [CrossRef]
- Stalin, N.; Srinivasan, P. Characterization of Vibrio Parahaemolyticus and its Specific Phage from Shrimp Pond in Palk Strait, South East Coast of India. Biologicals 2016, 44, 526–533. [Google Scholar] [CrossRef]
- Han, N.; Mizan, M.F.R.; Jahid, I.K.; Ha, S. Biofilm Formation by Vibrio Parahaemolyticus on Food and Food Contact Surfaces Increases with Rise in Temperature. Food Control 2016, 70, 161–166. [Google Scholar] [CrossRef]
- Lopatek, M.; Wieczorek, K.; Osek, J. Antimicrobial Resistance, Virulence Factors, and Genetic Profiles of Vibrio Parahaemolyticus from Seafood. Appl. Environ. Microb. 2018, 84, e00537-18. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.; Ab Mutalib, N.; Law, J.W.; Wong, S.H.; Letchumanan, V. Discovery on Antibiotic Resistance Patterns of Vibrio Parahaemolyticus in Selangor Reveals Carbapenemase Producing Vibrio Parahaemolyticus in Marine and Freshwater Fish. Front. Microbiol. 2018, 9, 2513. [Google Scholar] [CrossRef]
- Popova, K.B.; Valsamatzi-Panagiotou, A.; Penchovsky, R. New Drug Discovery Strategies for Targeting Drug-Resistant Bacteria. Environ. Chem. Lett. 2021, 19, 1995–2004. [Google Scholar] [CrossRef]
- Pereira, C.; Costa, P.; Duarte, J.; Balcao, V.M.; Almeida, A. Phage Therapy as a Potential Approach in the Biocontrol of Pathogenic Bacteria Associated with Shellfish Consumption. Int. J. Food Microbiol. 2021, 338, 108995. [Google Scholar] [CrossRef]
- Tian, F.; Li, J.; Nazir, A.; Tong, Y. Bacteriophage—A Promising Alternative Measure for Bacterial Biofilm Control. Infect. Drug Resist. 2021, 14, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zhang, H.; Gu, D.; Ma, Y.; Zhou, X. Identification of a Novel Bacterial Receptor that Binds Tail Tubular Proteins and Mediates Phage Infection of Vibrio Parahaemolyticus. Emerg. Microbes Infect. 2020, 9, 855–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Jin, J.; Choi, Y.; Kim, J. LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity. Viruses 2020, 12, 1340. [Google Scholar] [CrossRef]
- Son, B.; Kong, M.; Cha, Y.; Bai, J.; Ryu, S. Simultaneous Control of Staphylococcus Aureus and Bacillus Cereus Using a Hybrid Endolysin LysB4EAD-LysSA11. Antibiotics 2020, 9, 906. [Google Scholar] [CrossRef]
- Abdelrahman, F.; Easwaran, M.; Daramola, O.I.; Ragab, S.; Lynch, S.; Oduselu, T.J.; Khan, F.M.; Ayobami, A.; Adnan, F.; Torrents, E.; et al. Phage-Encoded Endolysins. Antibiotics 2021, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.C.B.; Chen, X.; Ho, M.K.Y.; Xia, J.; Leung, S.S.Y. Bacteriophage-Derived Endolysins to Target Gram-Negative Bacteria. Int. J. Pharmaceut. 2020, 589, 119833. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Lee, N.; Chun, H.; Chang, H. Characterization of a Novel Endolysin From Bacteriophage infecting Vibrio Parahaemolyticus, vB_VpaP_KF2. Appl. Biol. Chem. 2020, 63, 40. [Google Scholar] [CrossRef]
- Barr, J.J.; Auro, R.; Furlan, M.; Whiteson, K.L.; Erb, M.L.; Pogliano, J.; Stotland, A.; Wolkowicz, R.; Cutting, A.S.; Doran, K.S.; et al. Bacteriophage Adhering to Mucus Provide a Non-Host-Derived Immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 10771–10776. [Google Scholar] [CrossRef] [Green Version]
- Lomelí-Ortega, C.O.; Martínez-Sández, A.J.; Barajas-Sandoval, D.R.; Reyes, A.G.; Magallón-Barajas, F.; Veyrand-Quíros, B.; Gannon, L.; Harrison, C.; Michniewski, S.; Millard, A.; et al. Isolation and Characterization of Vibrio phage vB_Vc_SrVc9: An Effective Agent in Preventing Vibrio Campbellii Infections in Brine Shrimp Nauplii (Artemia Franciscana). J. Appl. Microbiol. 2021, 131, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Latka, A.; Drulis-Kawa, Z. Advantages and Limitations of Microtiter Biofilm Assays in the Model of Antibiofilm Activity of Klebsiella Phage KP34 and its Depolymerase. Sci. Rep. 2020, 10, 20338. [Google Scholar] [CrossRef] [PubMed]
- Mangieri, N.; Foschino, R.; Picozzi, C. Application of Bacteriophages on Shiga Toxin-Producing Escherichia Coli (STEC) Biofilm. Antibiotics 2021, 10, 1423. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Giri, S.S.; Yun, S.; Kim, S.W.; Han, S.J.; Kwon, J.; Oh, W.T.; Lee, S.B.; Park, Y.H.; Park, S.C. Two Novel Bacteriophages Control Multidrug- and Methicillin-Resistant Staphylococcus Pseudintermedius Biofilm. Front. Med. 2021, 8, 359. [Google Scholar] [CrossRef]
- Matamp, N.; Bhat, S.G. Genome Characterization of Novel Lytic Myoviridae Bacteriophage ϕVP-1 Enhances its Applicability Against MDR-biofilm-forming Vibrio Parahaemolyticus. Arch. Virol. 2020, 165, 387–396. [Google Scholar] [CrossRef]
- Yin, Y.; Ni, P.; Liu, D.; Yang, S.; Almeida, A.; Guo, Q.; Zhang, Z.; Deng, L.; Wang, D. Bacteriophage Potential Against Vibrio Parahaemolyticus Biofilms. Food Control 2019, 98, 156–163. [Google Scholar] [CrossRef]
- Ning, H.; Cong, Y.; Lin, H.; Wang, J. Development of Cationic Peptide Chimeric Lysins Based on Phage Lysin Lysqdvp001 and their Antibacterial Effects Against Vibrio Parahaemolyticus: A Preliminary Study. Int. J. Food Microbiol. 2021, 358, 109396. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VP Strains | Serotype | FE11 | VP Strains | Serotype | FE11 | VP Strains | Serotype | FE11 | VP Strains | Serotype | FE11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| O1–1 | O1 | − | O2–91 | O2 | + | O4–112 | O4 | − | O10–127 | O10 | − |
| O1–2 | O1 | + | O2–92 | O2 | − | O4–113 | O4 | + | O10–128 | O10 | − |
| O1–3 | O1 | + | O2–93 | O2 | − | O4–114 | O4 | + | O10–129 | O10 | − |
| O1–4 | O1 | − | O2–94 | O2 | − | O4–115 | O4 | − | O10–130 | O10 | −− |
| O1–36 | O1 | − | O2–95 | O2 | − | O5–15 | O5 | + | O10–131 | O10 | − |
| O1–37 | O1 | − | O2–96 | O2 | − | O5–16 | O5 | − | O10–132 | O10 | − |
| O1–38 | O1 | − | O2–97 | O2 | + | O5–17 | O5 | − | O11–29 | O11 | − |
| O1–62 | O1 | − | O2–98 | O2 | − | O5–49 | O5 | − | O11–30 | O11 | + |
| O1–63 | O1 | + | O2–99 | O2 | − | O5–50 | O5 | − | O11–31 | O11 | + |
| O1–64 | O1 | + | O2–100 | O2 | − | O5–116 | O5 | − | O11–56 | O11 | − |
| O1–65 | O1 | − | O2–101 | O2 | + | O5–117 | O5 | − | O11–57 | O11 | − |
| O1–66 | O1 | − | O2–102 | O2 | − | O5–118 | O5 | − | O11–58 | O11 | − |
| O1–67 | O1 | + | O2–103 | O2 | − | O6–18 | O6 | + | O11–133 | O11 | + |
| O1–68 | O1 | − | O3–8 | O2 | − | O6–19 | O6 | + | O11–134 | O11 | + |
| O1–69 | O1 | − | O3–9 | O2 | − | O6–20 | O6 | + | O11–135 | O11 | + |
| O1–70 | O1 | − | O3–10 | O3 | − | O6–120 | O6 | − | O11–136 | O11 | − |
| O1–71 | O1 | − | O3–11 | O3 | − | O8–21 | O8 | + | O11–137 | O11 | + |
| O1–72 | O1 | + | O3–42 | O3 | − | O8–22 | O8 | − | O11–138 | O11 | − |
| O1–73 | O1 | − | O3–43 | O3 | − | O8–51 | O8 | − | O12–32 | O12 | − |
| O2–5 | O2 | − | O3–44 | O3 | − | O8–52 | O8 | − | O12–33 | O12 | + |
| O2–6 | O2 | − | O3–104 | O3 | + | O8–53 | O8 | − | O12–34 | O12 | − |
| O2–7 | O2 | − | O3–105 | O3 | − | O8–121 | O8 | − | O12–35 | O12 | − |
| O2–39 | O2 | − | O3–106 | O3 | + | O8–122 | O8 | + | O12–60 | O12 | − |
| O2–40 | O2 | − | O3–107 | O3 | + | O8–123 | O8 | − | O12–61 | O12 | − |
| O2–41 | O2 | − | O4–12 | O4 | + | O8–124 | O8 | − | O12–140 | O12 | − |
| O2–82 | O2 | − | O4–14 | O4 | + | O8–125 | O8 | − | O12–141 | O12 | − |
| O2–83 | O2 | − | O4–45 | O4 | − | O8–126 | O8 | − | O12–143 | O12 | − |
| O2–84 | O2 | − | O4–46 | O4 | − | O9–24 | O9 | − | O12–144 | O12 | − |
| O2–85 | O2 | − | O4–47 | O4 | + | O10–25 | O10 | − | O12–145 | O12 | − |
| O2–86 | O2 | − | O4–48 | O4 | − | O10–26 | O10 | − | O12–146 | O12 | + |
| O2–87 | O2 | − | O4–108 | O4 | − | O10–27 | O10 | + | O12–147 | O12 | − |
| O2–88 | O2 | − | O4–109 | O4 | + | O10–28 | O10 | − | |||
| O2–89 | O2 | − | O4–110 | O4 | − | O10–54 | O10 | + | |||
| O2–90 | O2 | − | O4–111 | O4 | + | O10–55 | O10 | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Chen, H.; Huang, Q.; Xie, Z.; Liu, Z.; Zhang, J.; Ding, Y.; Chen, M.; Xue, L.; Wu, Q.; et al. Characterization of the Novel Phage vB_VpaP_FE11 and Its Potential Role in Controlling Vibrio parahaemolyticus Biofilms. Viruses 2022, 14, 264. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020264
Yang M, Chen H, Huang Q, Xie Z, Liu Z, Zhang J, Ding Y, Chen M, Xue L, Wu Q, et al. Characterization of the Novel Phage vB_VpaP_FE11 and Its Potential Role in Controlling Vibrio parahaemolyticus Biofilms. Viruses. 2022; 14(2):264. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020264
Chicago/Turabian StyleYang, Meiyan, Hanfang Chen, Qiaolan Huang, Zhuanbei Xie, Zekun Liu, Jumei Zhang, Yu Ding, Moutong Chen, Liang Xue, Qingping Wu, and et al. 2022. "Characterization of the Novel Phage vB_VpaP_FE11 and Its Potential Role in Controlling Vibrio parahaemolyticus Biofilms" Viruses 14, no. 2: 264. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020264