
Genome-Wide Identification and Gene Expression Analysis of the OTU DUB Family in Oryza sativa

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Identification of OTU Family Genes in Rice, Arabidopsis, and Maize
2.2. Multiple Sequence Alignment and Phylogenetic Analysis
2.3. Analysis of Conserved Gene Domains and Motifs
2.4. Gene Structure and Chromosomal Distribution of rice OTU Gene Analysis
2.5. Prediction Cis-Acting Elements of Rice OTU Genes
2.6. Plant Materials Acquisition and Hormone Treatments
2.7. Rice Viruses Infect Plant Materials
2.8. RNA Extraction and RT-qPCR
3. Results
3.1. Identification and Analysis of OTU Family in Rice
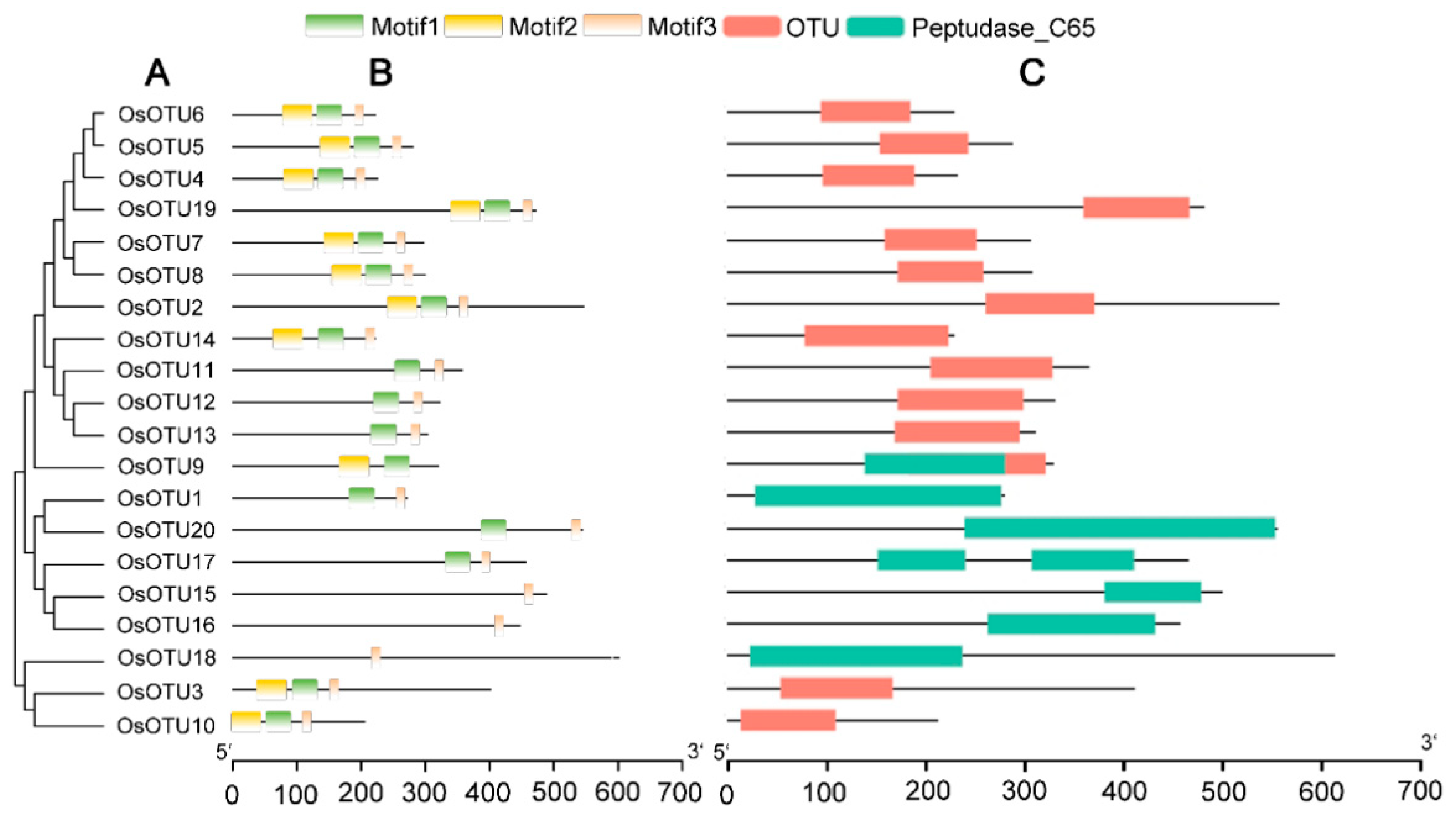
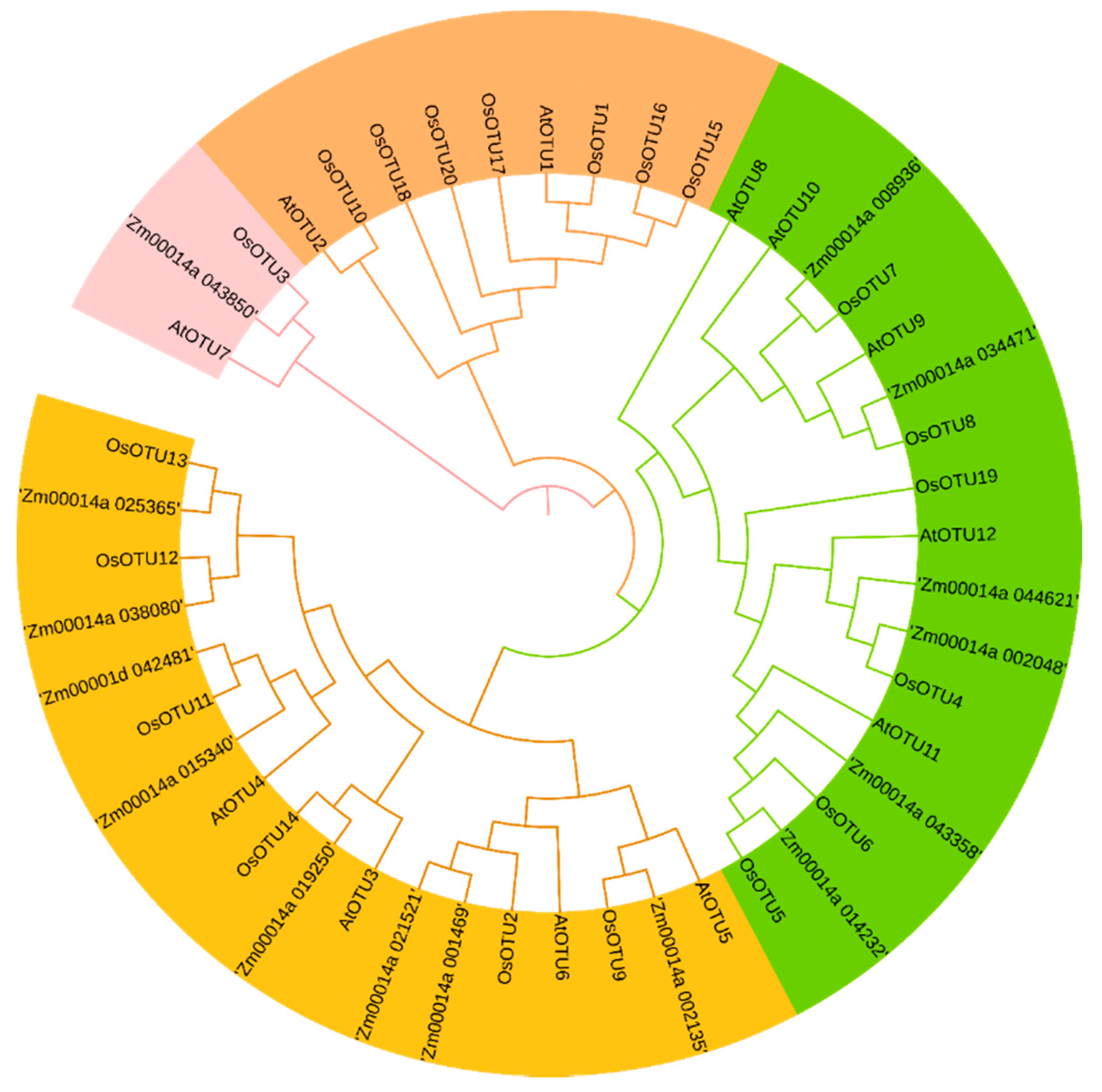
3.2. Analysis of Domain Organization, Conserved Motifs, and Phylogenetics of OTU
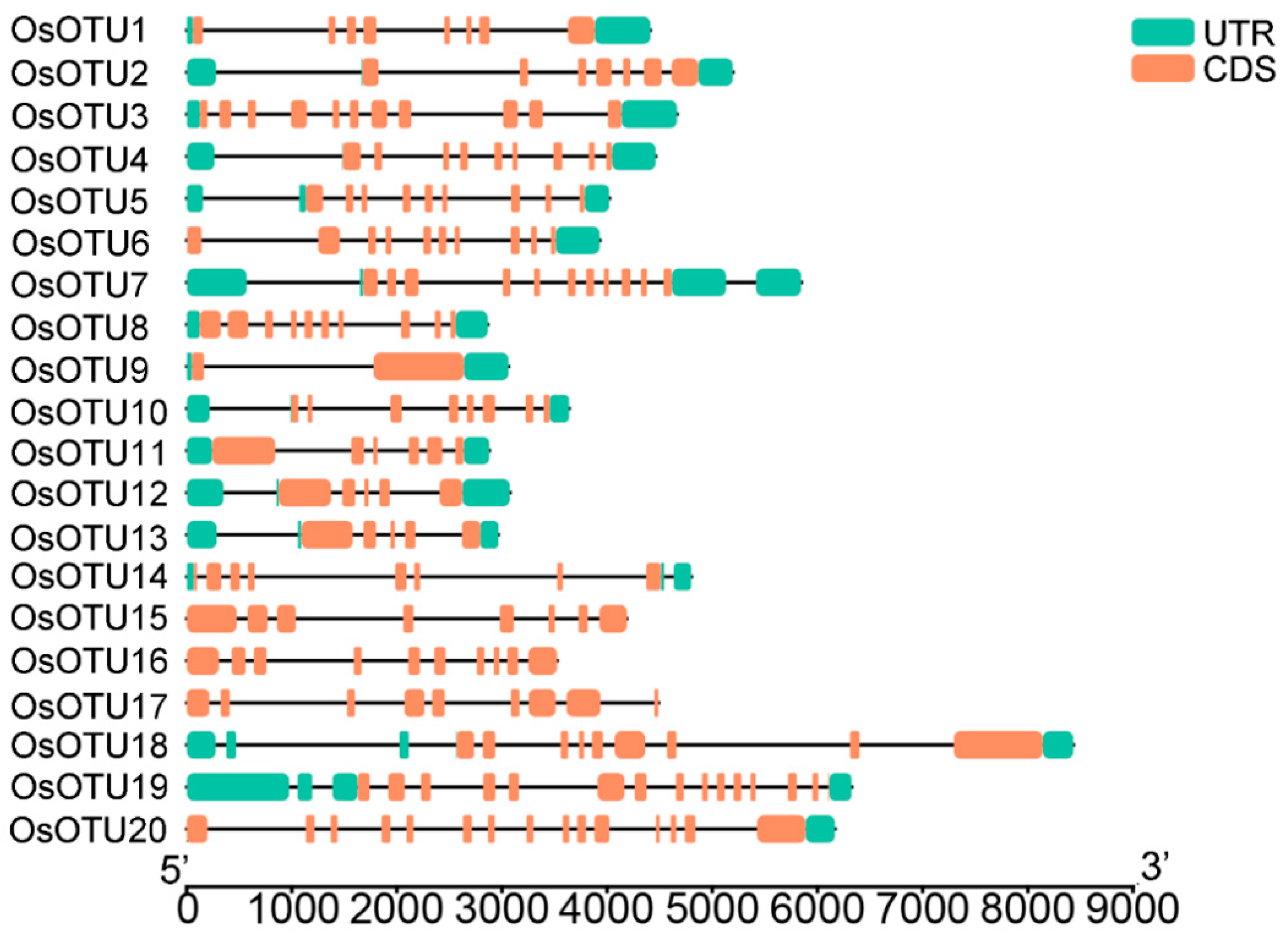
3.3. Gene Structure and Chromosomal Distribution Analysis of Rice OTU Genes
3.4. Prediction of OsOTU Cis-Acting Elements
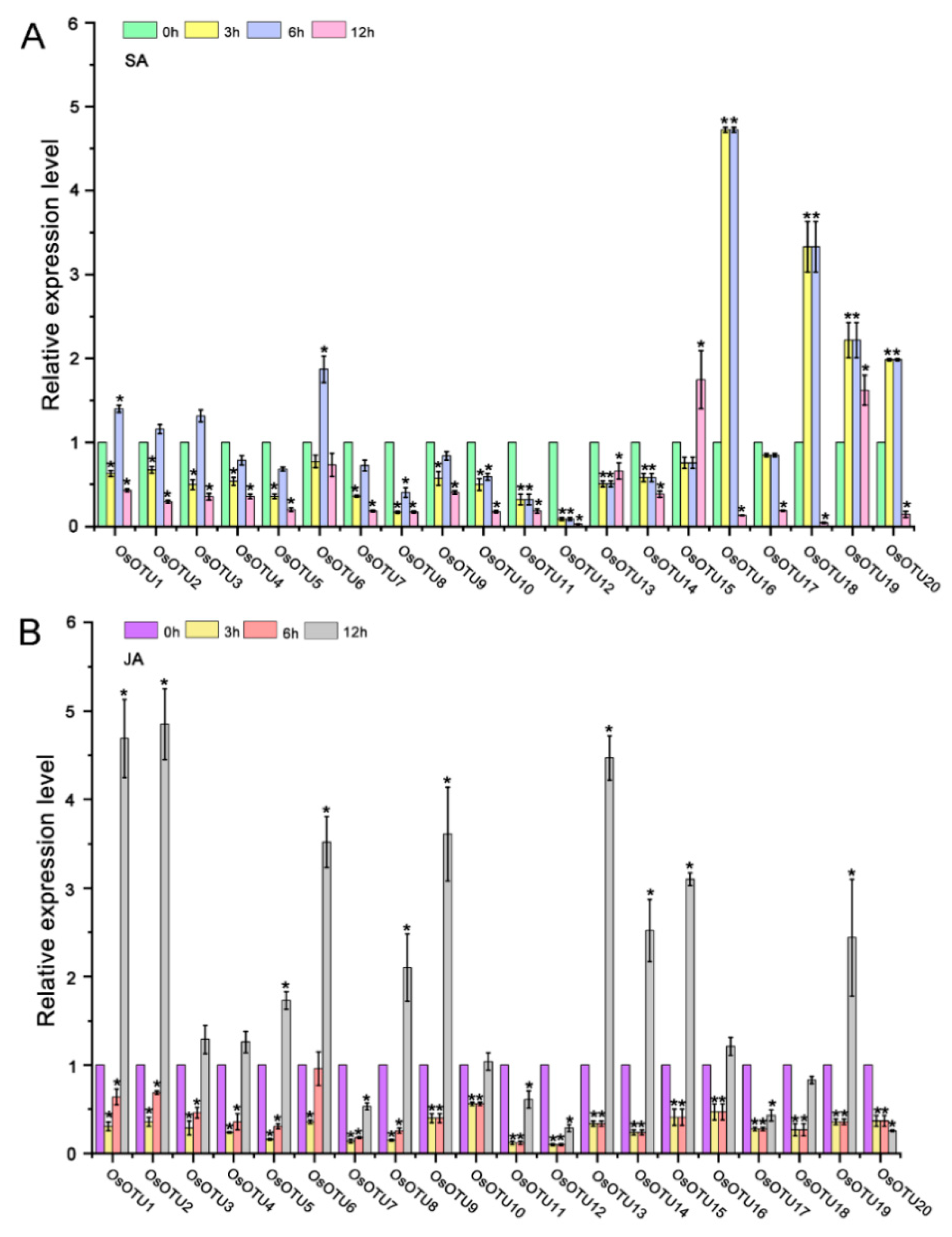
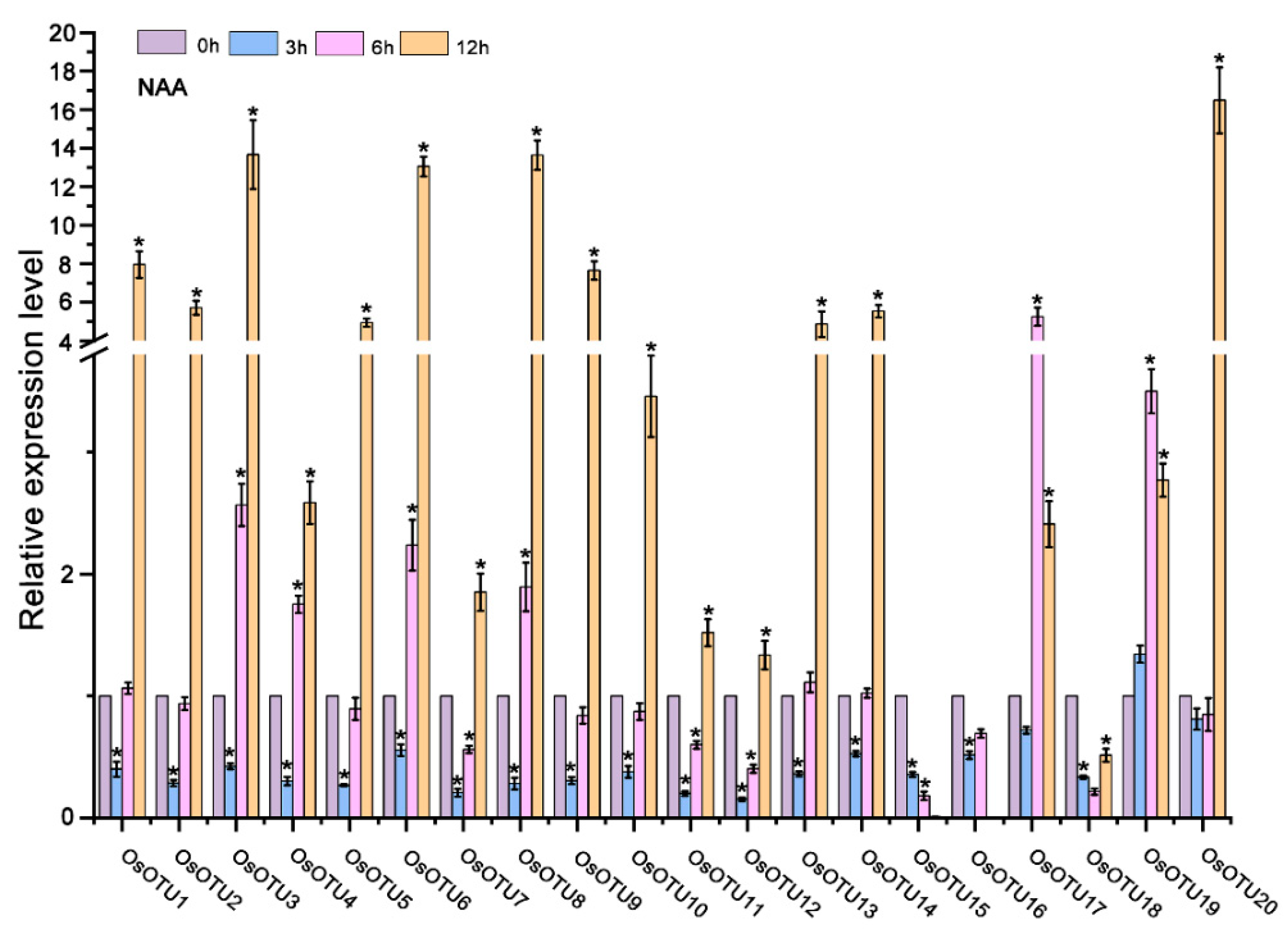
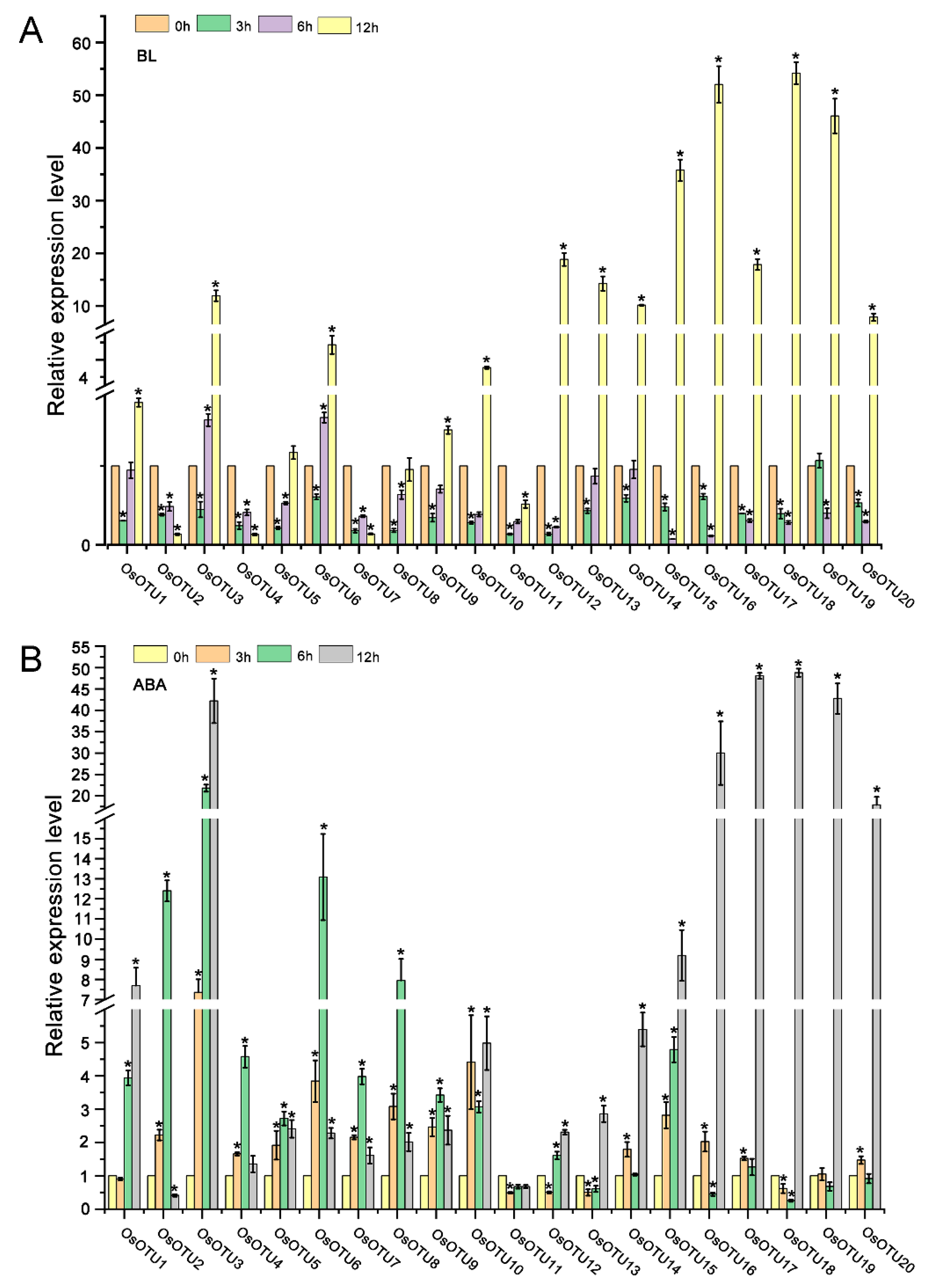
3.5. Expression Patterns of Rice OTU Genes under Hormone Treatments
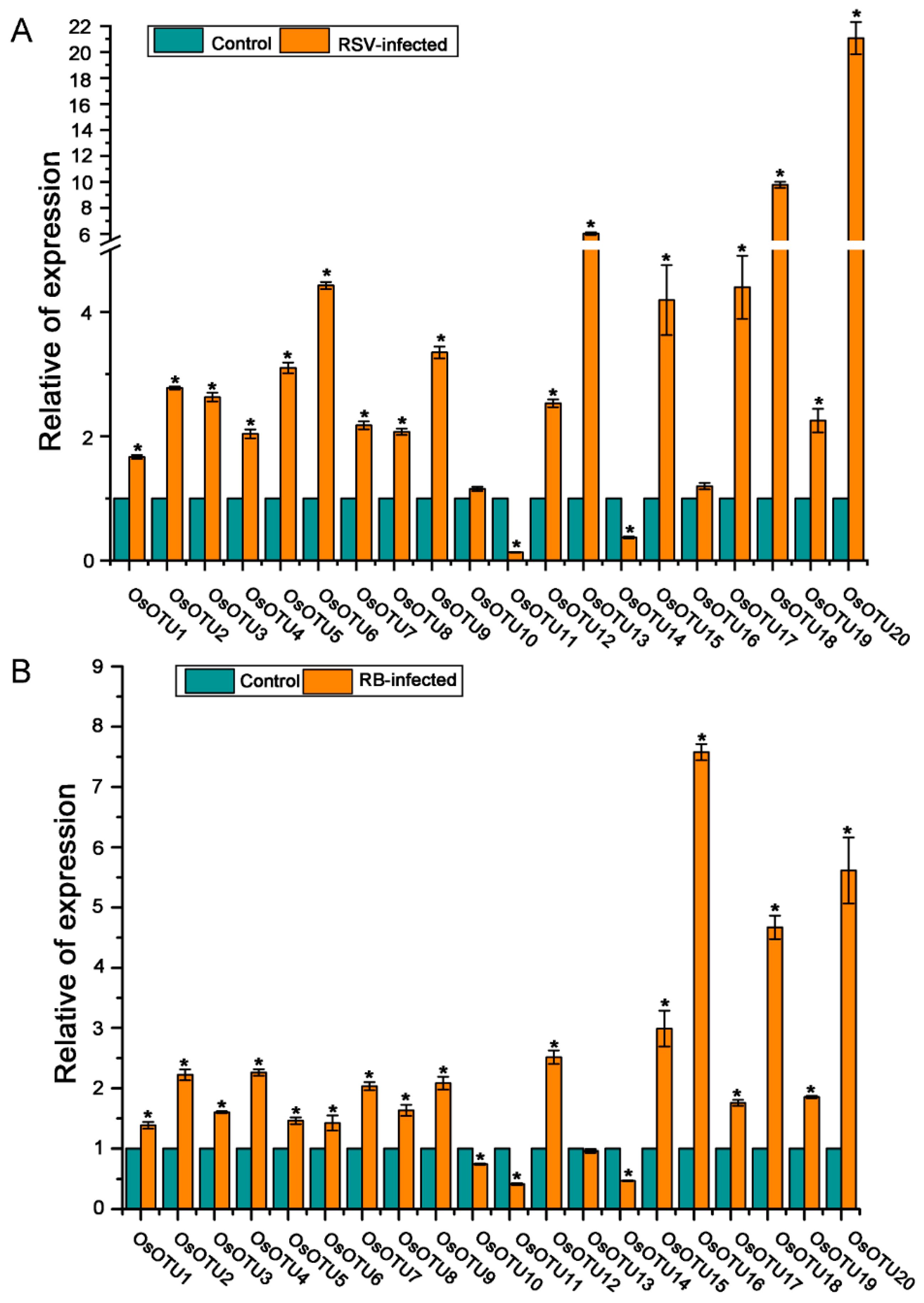
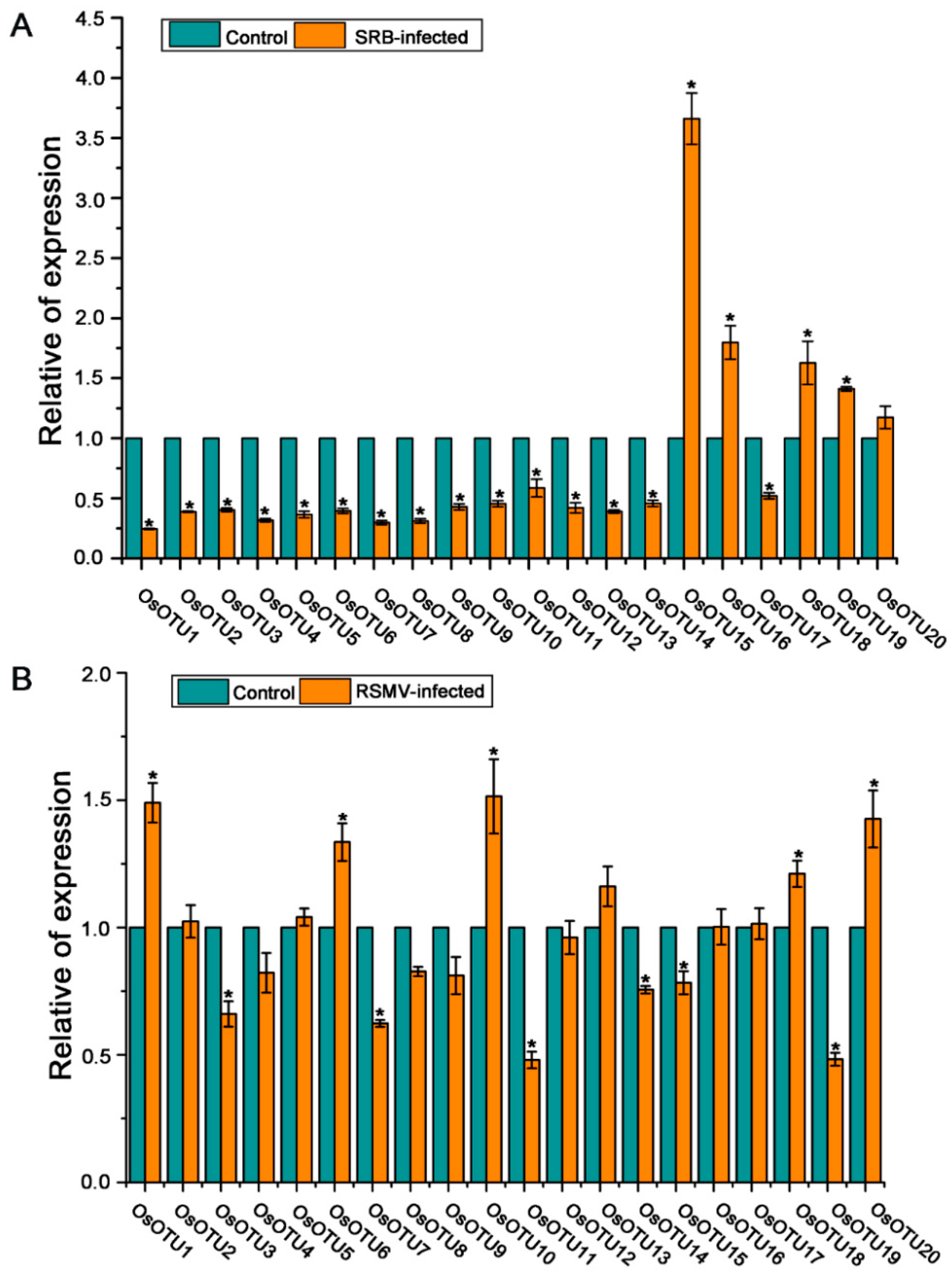
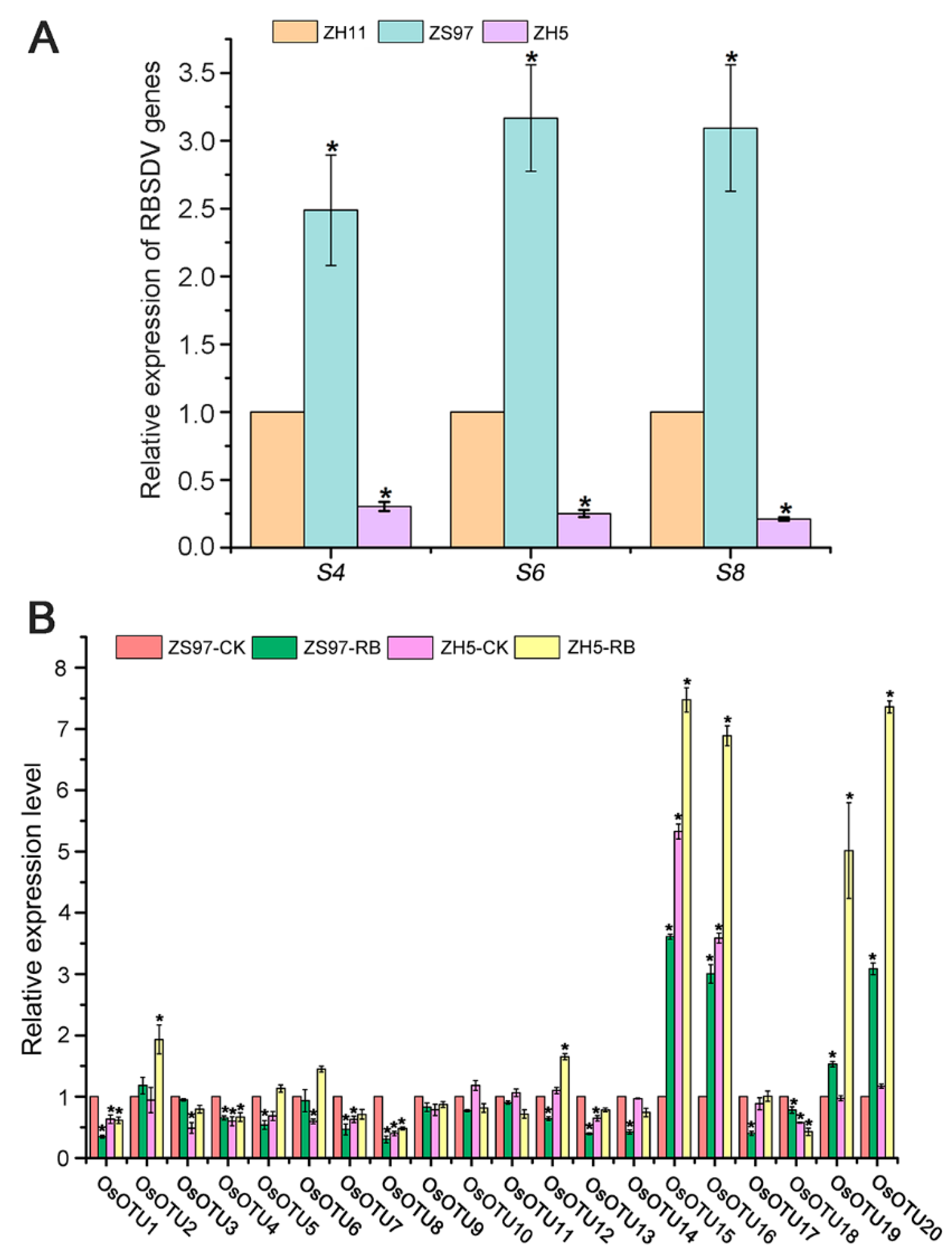
3.6. Analysis of Rice OTU Genes Expression Profiling after RSV, RBSDV, SRBSDV, and RSMV Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Goldstein, G.; Scheid, M.; Hammerling, U.; Schlesinger, D.H.; Niall, H.D.; Boyse, E.A. Isolation of a polypeptide that has lymphocyte-differentiating properties and is probably represented universally in living cells. Proc. Natl. Acad. Sci. USA 1975, 72, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershko, A.; Ciechanover, A. The Ubiquitin System. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Luo, Z.-Q. Post-translational regulation of ubiquitin signaling. J. Cell Biol. 2019, 218, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Rape, M. Reverse the curse—The role of deubiquitination in cell cycle control. Curr. Opin. Cell Biol. 2008, 20, 156–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guterman, A.; Glickman, M.H. Deubiquitinating enzymes are IN (trinsic to proteasome function). Curr. Protein Pept. Sci. 2004, 5, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Komada, M. Controlling receptor downregulation by ubiquitination and deubiquitination. Curr. Drug Discov. Technol. 2008, 5, 78–84. [Google Scholar] [CrossRef]
- Daniel, J.A.; Grant, P.A. Multi-tasking on chromatin with the SAGA coactivator complexes. Mutat. Res. Mol. Mech. Mutagen. 2007, 618, 135–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, R.D.; D’Andrea, A.D. The Fanconi Anemia/BRCA pathway: New faces in the crowd. Genes Dev. 2005, 19, 2925–2940. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, A.; Xu, M.; Chen, Z. Ubiquitin-mediated activation of TAK1 and IKK. Oncogene 2007, 26, 3214–3226. [Google Scholar] [CrossRef] [Green Version]
- Rytkönen, A.; Holden, D.W. Bacterial interference of ubiquitination and deubiquitination. Cell Host Microbe 2007, 1, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, N. Ubiquitin System. Int. J. Mol. Sci. 2018, 19, 1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balakirev, M.Y.; Tcherniuk, S.O.; Jaquinod, M.; Chroboczek, J. Otubains: A new family of cysteine proteases in the ubiquitin pathway. EMBO Rep. 2003, 4, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Fu, L.; Sui, Y.; Zhang, L. The function and regulation of OTU deubiquitinases. Front. Med. 2020, 14, 542–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelmann, M.J.; Iphöfer, A.; Akutsu, M.; Altun, M.; Di Gleria, K.; Kramer, H.B.; Fiebiger, E.; Dhe-Paganon, S.; Kessler, B.M. Structural basis and specificity of human otubain 1-mediated deubiquitination. Biochem. J. 2009, 418, 379–390. [Google Scholar] [CrossRef]
- Wang, T.; Yin, L.; Cooper, E.M.; Lai, M.-Y.; Dickey, S.; Pickart, C.M.; Fushman, D.; Wilkinson, K.D.; Cohen, R.E.; Wolberger, C. Evidence for bidentate substrate binding as the basis for the K48 linkage specificity of otubain 1. J. Mol. Biol. 2009, 386, 1011–1023. [Google Scholar] [CrossRef] [Green Version]
- Wiener, R.; Zhang, X.; Wang, T.; Wolberger, C. The mechanism of OTUB1-mediated inhibition of ubiquitination. Nature 2012, 483, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Radjacommare, R.; Usharani, R.; Kuo, C.-H.; Fu, H. Distinct phylogenetic relationships and biochemical properties of Arabidopsis ovarian tumor-related deubiquitinases support their functional differentiation. Front. Plant Sci. 2014, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Schubert, A.F.; Nguyen, J.V.; Franklin, T.G.; Geurink, P.P.; Roberts, C.G.; Sanderson, D.J.; Miller, L.N.; Ovaa, H.; Hofmann, K.; Pruneda, J.N.; et al. Identification and characterization of diverse OTU deubiquitinases in bacteria. EMBO J. 2020, 39, e105127. [Google Scholar] [CrossRef]
- Zhao, S.; Gu, X.; Li, J.; Liang, C. The N-terminal cysteine protease domain of rice stripe tenuivirus Pc1 possesses deubiquitinating enzyme activity. Virus Genes 2021, 57, 117–120. [Google Scholar] [CrossRef]
- Capodagli, G.C.; McKercher, M.A.; Baker, E.A.; Masters, E.M.; Brunzelle, J.S.; Pegan, S.D. Structural analysis of a viral ovarian tumor domain protease from the Crimean-Congo hemorrhagic fever virus in complex with covalently bonded ubiquitin. J. Virol. 2011, 85, 3621–3630. [Google Scholar] [CrossRef] [Green Version]
- Frias-Staheli, N.; Giannakopoulos, N.V.; Kikkert, M.; Taylor, S.L.; Bridgen, A.; Paragas, J.; Richt, J.A.; Rowland, R.R.; Schmaljohn, C.S.; Lenschow, D.J. Ovarian tumor domain-containing viral proteases evade ubiquitin-and ISG15-dependent innate immune responses. Cell Host Microbe 2007, 2, 404–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, J.-I.; Nonomura, K.-I.; Ikeda, K.; Yamaki, S.; Inukai, Y.; Yamagishi, H.; Kitano, H.; Nagato, Y. Rice Plant Development: From Zygote to Spikelet. Plant Cell Physiol. 2005, 46, 23–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2011, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37 (Suppl. 2), W202–W208. [Google Scholar] [CrossRef]
- Ouyang, S.; Zhu, W.; Hamilton, J.; Lin, H.; Campbell, M.; Childs, K.; Thibaud-Nissen, F.; Malek, R.L.; Lee, Y.; Zheng, L.; et al. The TIGR Rice Genome Annotation Resource: Improvements and new features. Nucleic Acids Res. 2006, 35 (Suppl. 1), D883–D887. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Rombauts, S.; Déhais, P.; Van Montagu, M.; Rouzé, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Li, L.; He, Y.; Qin, Q.; Chen, C.; Wei, Z.; Tan, X.; Xie, K.; Zhang, R.; Hong, G.; et al. Distinct modes of manipulation of rice auxin response factor OsARF17 by different plant RNA viruses for infection. Proc. Natl. Acad. Sci. USA 2020, 117, 9112. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hong, G.; Zhang, H.; Tan, X.; Li, L.; Kong, Y.; Sang, T.; Xie, K.; Wei, J.; Li, J.; et al. The OsGSK2 Kinase Integrates Brassinosteroid and Jasmonic Acid Signaling by Interacting with OsJAZ4. Plant Cell 2020, 32, 2806–2822. [Google Scholar] [CrossRef] [PubMed]
- Bachman, J. Chapter Two—Reverse-Transcription PCR (RT-PCR). Method Enzymol. 2013, 530, 67–74. [Google Scholar]
- Bustin, S.A.; Mueller, R. Real-time reverse transcription PCR (qRT-PCR) and its potential use in clinical diagnosis. Clin. Sci. 2005, 109, 365–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, P.; Shivhare, R.; Kaur, A.; Bansal, S.; Sonah, H.; Deshmukh, R.; Giri, J.; Lata, C.; Ram, H. Reference gene identification for gene expression analysis in rice under different metal stress. J. Biotechnol. 2021, 332, 83–93. [Google Scholar] [CrossRef]
- Abtahi, S.H.; Mohammadi, M.H.; Farsani, M.A.; Aghelan, Z.; Salari, S. Evaluation of Sestrin 2, Adiponectin, AMPK, and mTOR Genes Expression in Acute Myeloid Leukemia Patients. Iran. J. Biotechnol. 2021, 19, e2860. [Google Scholar]
- Yan, J.; Ma, Z.; Xu, X.; Guo, A.-Y. Evolution, functional divergence and conserved exon–intron structure of bHLH/PAS gene family. Mol. Genet. Genom. 2014, 289, 25–36. [Google Scholar] [CrossRef]
- Zheng, Z.-M. Split genes and their expression in Kaposi’s sarcoma-associated herpesvirus. Rev. Med. Virol. 2003, 13, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, C.M.; Finer, J.J. A novel cis-acting element in the GmERF3 promoter contributes to inducible gene expression in soybean and tobacco after wounding. Plant Cell Rep. 2016, 35, 303–316. [Google Scholar] [CrossRef]
- Cho, W.K.; Lian, S.; Kim, S.-M.; Park, S.-H.; Kim, K.-H. Current Insights into Research on Rice stripe virus. Plant Pathol. J. 2013, 29, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Xiao, D.; Wu, J.; Jia, D.; Yuan, Z.; Liu, Y.; Hu, L.; Han, Z.; Wei, T.; Lin, Q.; et al. P2 of Rice stripe virus (RSV) interacts with OsSGS3 and is a silencing suppressor. Mol. Plant Pathol. 2011, 12, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Toriyama, S.; Takahashi, M.; Sano, Y.; Shimizu, T.; Ishihama, A. Nucleotide sequence of RNA 1, the largest genomic segment of rice stripe virus, the prototype of the tenuiviruses. J. Gen. Virol. 1994, 75, 3569–3579. [Google Scholar] [CrossRef]
- Hamamatsu, C.; Toriyama, S.; Toyoda, T.; Ishihama, A. Ambisense coding strategy of the rice stripe virus genome: In vitro translation studies. J. Gen. Virol. 1993, 74, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Shikata, E.; Kitagawa, Y. Rice black-streaked dwarf virus: Its properties, morphology and intracellular localization. Virology 1977, 77, 826–842. [Google Scholar] [CrossRef]
- Fang, S.; Yu, J.; Feng, J.; Han, C.; Li, D.; Liu, Y. Identification of rice black-streaked dwarf fijivirus in maize with rough dwarf disease in China. Arch. Virol. 2001, 146, 167–170. [Google Scholar] [CrossRef]
- Wang, Z.-H.; Fang, S.-G.; Xu, J.-L.; Sun, L.-Y.; Li, D.-W.; Yu, J.-L. Sequence Analysis of the Complete Genome of Rice Black-Streaked Dwarf Virus Isolated from Maize with Rough Dwarf Disease. Virus Genes 2003, 27, 163–168. [Google Scholar] [CrossRef]
- Zhang, H.-M.; Yang, J.; Chen, J.-P.; Adams, M.J. A black-streaked dwarf disease on rice in China is caused by a novel fijivirus. Arch. Virol. 2008, 153, 1893–1898. [Google Scholar] [CrossRef]
- Zhou, G.; Wen, J.; Cai, D.; Li, P.; Xu, D.; Zhang, S. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Yang, X.; Huang, J.; Liu, C.; Chen, B.; Zhang, T.; Zhou, G. Rice Stripe Mosaic Virus, a Novel Cytorhabdovirus Infecting Rice via Leafhopper Transmission. Front. Microbiol. 2017, 7, 2140. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Li, Y. Rice reoviruses in insect vectors. Annu. Rev. Phytopathol. 2016, 54, 99–120. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, H.; Sun, Z.; Li, J.; Hong, G.; Zhu, Q.; Zhou, X.; MacFarlane, S.; Yan, F.; Chen, J. Jasmonic acid-mediated defense suppresses brassinosteroid-mediated susceptibility to Rice black streaked dwarf virus infection in rice. New Phytol. 2017, 214, 388–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhang, H.; Chen, C.; Huang, H.; Tan, X.; Wei, Z.; Li, J.; Yan, F.; Zhang, C.; Chen, J.; et al. A class of independently evolved transcriptional repressors in plant RNA viruses facilitates viral infection and vector feeding. Proc. Natl. Acad. Sci. USA 2021, 118, e2016673118. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Gong, Y.; Wang, Q.; Guo, H.; Xiao, W. Arabidopsis OTU1, a linkage-specific deubiquitinase, is required for endoplasmic reticulum-associated protein degradation. Plant J. 2020, 101, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Li, Y.; Tang, G.; Hui, S.; Yang, Z.; Zhao, J.; Liu, H.; Cao, J.; Yuan, M. Dynamic phytohormone profiling of rice upon rice black-streaked dwarf virus invasion. J. Plant Physiol. 2018, 228, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Li, L.; Zhang, H.; Wang, R.; Tan, X.; He, Y.; Hong, G.; Li, J.; Ming, F.; Yao, X.; et al. Abscisic acid negatively modulates plant defence against rice black-streaked dwarf virus infection by suppressing the jasmonate pathway and regulating reactive oxygen species levels in rice. Plant Cell Environ. 2018, 41, 2504–2514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tan, X.; Li, L.; He, Y.; Hong, G.; Li, J.; Lin, L.; Cheng, Y.; Yan, F.; Chen, J.; et al. Suppression of auxin signalling promotes rice susceptibility to Rice black streaked dwarf virus infection. Mol. Plant Pathol. 2019, 20, 1093–1104. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Gene Name | Gene ID Number | Amino Acid Residues | OTU Domain | Peptidase C65 Domain | PI |
|---|---|---|---|---|---|---|
| 1 | OsOTU1 | >LOC_Os08g42540.1 | 274 | 22–273 | 4.76 | |
| 2 | OsOTU2 | >LOC_Os04g33780.1 | 550 | 255–366 | 5.03 | |
| 3 | OsOTU3 | >LOC_Os04g57480.1 | 404 | 51–169 | 6.69 | |
| 4 | OsOTU4 | >LOC_Os04g32970.1 | 228 | 93–208 | 8.93 | |
| 5 | OsOTU5 | >LOC_Os02g07210.2 | 224 | 91–206 | 8.28 | |
| 6 | OsOTU6 | >LOC_Os06g45850.1 | 283 | 150–267 | 9.14 | |
| 7 | OsOTU7 | >LOC_Os02g57410.2 | 300 | 156–270 | 5.67 | |
| 8 | OsOTU8 | >LOC_Os03g64219.1 | 302 | 168–282 | 5.11 | |
| 9 | OsOTU9 | >LOC_Os04g52850.1 | 323 | 180–317 | 132–312 | 5.89 |
| 10 | OsOTU10 | >LOC_Os02g06890.1 | 208 | 11–123 | 5.08 | |
| 11 | OsOTU11 | >LOC_Os01g67490.1 | 360 | 202–330 | 10.68 | |
| 12 | OsOTU12 | >LOC_Os08g39560.1 | 325 | 169–297 | 8.60 | |
| 13 | OsOTU13 | >LOC_Os09g31280.1 | 306 | 166–292 | 9.10 | |
| 14 | OsOTU14 | >LOC_Os03g15930.2 | 224 | 76–220 | 8.71 | |
| 15 | OsOTU15 | >LOC_Os02g32180.1 | 492 | 355–473 | 5.80 | |
| 16 | OsOTU16 | >LOC_Os02g32190.1 | 450 | 248–427 | 9.58 | |
| 17 | OsOTU17 | >LOC_Os02g32280.1 | 459 | 129–269 289–406 | 6.96 | |
| 18 | OsOTU18 | >LOC_Os02g30974.1 | 605 | 5–234 | 8.13 | |
| 19 | OsOTU19 | >LOC_Os03g39230.1 | 475 | 354–469 | 8.54 | |
| 20 | OsOTU20 | >LOC_Os04g55840.1 | 548 | 228–547 | 4.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Yan, T.; Tan, X.; Wei, Z.; Li, Y.; Sun, Z.; Zhang, H.; Chen, J. Genome-Wide Identification and Gene Expression Analysis of the OTU DUB Family in Oryza sativa. Viruses 2022, 14, 392. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020392
Liu Q, Yan T, Tan X, Wei Z, Li Y, Sun Z, Zhang H, Chen J. Genome-Wide Identification and Gene Expression Analysis of the OTU DUB Family in Oryza sativa. Viruses. 2022; 14(2):392. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020392
Chicago/Turabian StyleLiu, Qiannan, Tingyun Yan, Xiaoxiang Tan, Zhongyan Wei, Yanjun Li, Zongtao Sun, Hehong Zhang, and Jianping Chen. 2022. "Genome-Wide Identification and Gene Expression Analysis of the OTU DUB Family in Oryza sativa" Viruses 14, no. 2: 392. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020392