
Methamphetamine Induces the Release of Proadhesive Extracellular Vesicles and Promotes Syncytia Formation: A Potential Role in HIV-1 Neuropathogenesis

 ,
, 
 and
and 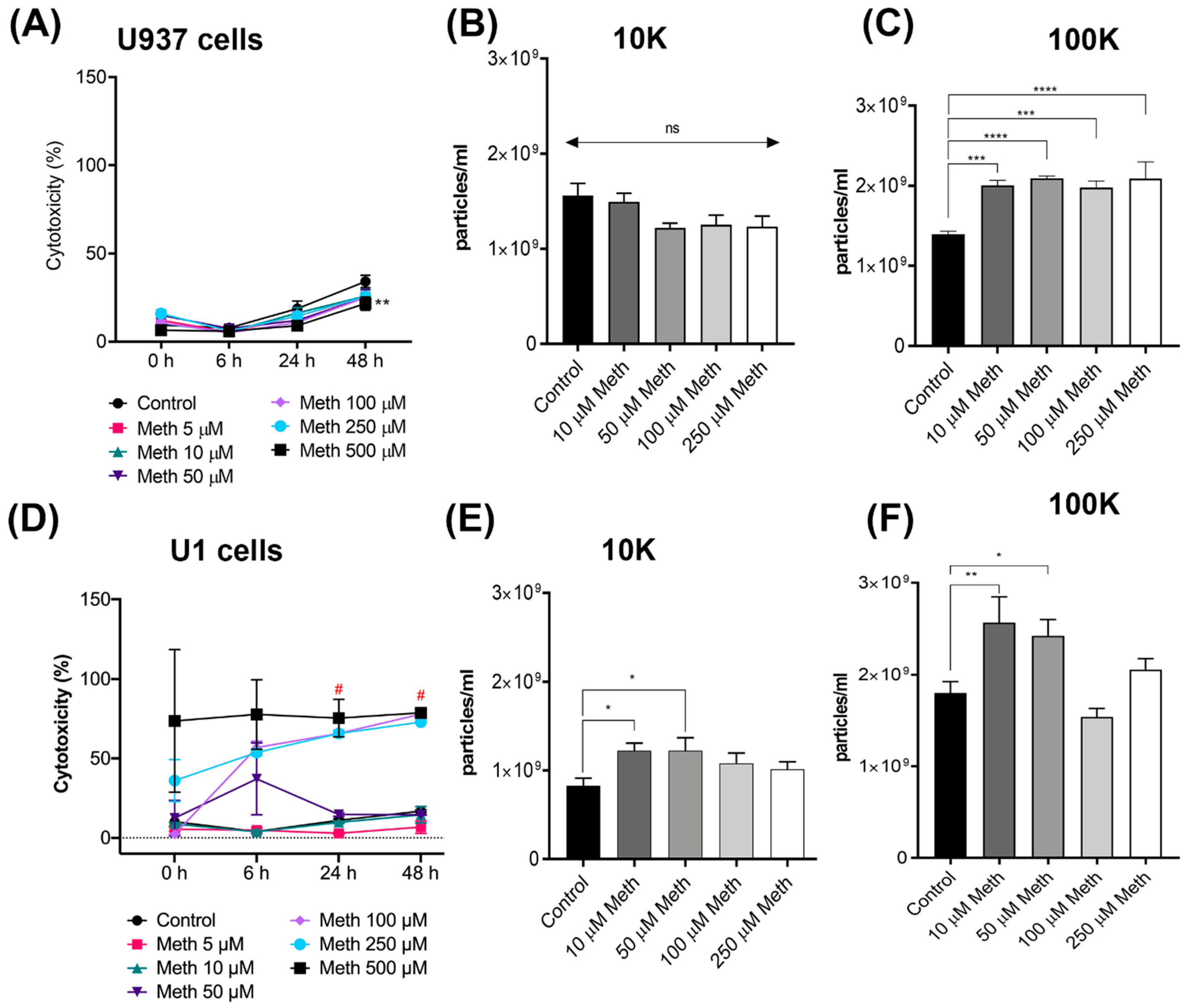{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Meth and cART Treatment
2.3. Cell Fixing and Staining
2.4. Extracellular Vesicles Isolations
2.5. Nanoparticle Tracking Analysis (NTA)
2.6. Western Blot
2.7. Labeling of Extracellular Vesicles and Confocal Microscopy
2.8. Cell Contact Inhibition Assay
2.9. Cell Toxicity Assay
2.10. RNA Isolation and qPCR
2.11. Syncytia Count
2.12. Statistics
3. Results
3.1. Impact of Meth on EVs Biogenesis in HIV Infected Macrophages
3.2. Meth Alters Genes in ESCRT-Dependent and Independent Pathways
3.3. Role of Meth on EVs Cargo and Expression of Viral Protein
3.4. Role of Meth and EVs in Syncytia and HIV Pathogenesis
3.5. Antibody Treatment against ICAM-1 and LFA-1 Antibodies Prevented Meth-Induced Syncytia Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EV(s) | Extracellular vesicle(s) |
| HIV | Human immunodeficiency virus-1 |
| Meth | Methamphetamine |
| cART | Combinational antiretroviral therapy |
| HAND | HIV associated neurological disorders |
| STDs | Sexually transmitted diseases |
| ICAM-1 | Intercellular adhesion molecule 1 |
| LFA-1 | Lymphocyte function-associated antigen 1 |
| Abs | Antibodies |
| CNS | Central nervous system |
| IL-1b | Interleukin-1b |
| EVs | Extracellular vesicles |
| HTLV-1 | Human T-cell leukemia virus-1 |
| MDMs | Monocyte-derived macrophages |
| VCAM | Vascular cell adhesion molecule |
| NTA | Nanoparticle tracking analysis |
| LDH | Lactate dehydrogenase |
| ESCRT | Endosomal sorting complexes required for transport |
| TEM | Transmission electron microscopy |
| cDNA | Complementary DNA |
| NIDA | National Institute on Drug Abuse |
| Env | Envelope |
| PMA | Phorbol 12-myristate 13-acetate |
| M-CSF | Macrophage colony-stimulating factor |
| DIC | Differential interference contrast |
| CERS | Ceramide synthases |
| MNC | Multinucleated cells |
References
- Sacktor, N.; Skolasky, R.L.; Seaberg, E.; Munro, C.; Becker, J.T.; Martin, E.; Ragin, A.; Levine, A.; Miller, E. Prevalence of HIV-associated neurocognitive disorders in the Multicenter AIDS Cohort Study. Neurology 2015, 86, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, S.P.; Moore, D.J.; Weber, E.; Grant, I. Cognitive Neuropsychology of HIV-Associated Neurocognitive Disorders. Neuropsychol. Rev. 2009, 19, 152–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Carbonell, D.; Ye, F.; Ramanath, N.; Garcia-Mesa, Y.; Knapp, P.E.; Hauser, K.F.; Karn, J. Cross-talk between microglia and neurons regulates HIV latency. PLOS Pathog. 2019, 15, e1008249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, I. Interrogating the impact of combination antiretroviral therapies on HIV-associated neurocognitive disorders. HIV Med. 2021, 22, 783–790. [Google Scholar] [CrossRef]
- Tomkins, A.; George, R.; Kliner, M. Sexualised drug taking among men who have sex with men: A systematic review. Perspect. Public Health 2019, 139, 23–33. [Google Scholar] [CrossRef]
- Bourne, A.; Reid, D.; Hickson, F.; Torres-Rueda, S.; Steinberg, P.; Weatherburn, P. “Chemsex” and harm reduction need among gay men in South London. Int. J. Drug Policy 2015, 26, 1171–1176. [Google Scholar] [CrossRef]
- Howarth, A.; Apea, V.; Michie, S.; Morris, S.; Sachikonye, M.; Mercer, C.; Evans, A.; Delpech, V.; Sabin, C.; Burns, F. The association between use of chemsex drugs and HIV clinic attendance among gay and bisexual men living with HIV in London. HIV Med. 2021, 22, 641–649. [Google Scholar] [CrossRef]
- Vosburgh, H.W.; Mansergh, G.; Sullivan, P.; Purcell, D. A Review of the Literature on Event-Level Substance Use and Sexual Risk Behavior among Men Who Have Sex with Men. AIDS Behav. 2012, 16, 1394–1410. [Google Scholar] [CrossRef]
- Rosińska, M.; Gios, L.; Nöstlinger, C.; Berghe, W.V.; Marcus, U.; Schink, S.; Sherriff, N.; Jones, A.-M.; Folch, C.; Dias, S.; et al. Prevalence of drug use during sex amongst MSM in Europe: Results from a multi-site bio-behavioural survey. Int. J. Drug Policy 2018, 55, 231–241. [Google Scholar] [CrossRef]
- Schecke, H.; Lea, T.; Bohn, A.; Köhler, T.; Sander, D.; Scherbaum, N.; Deimel, D. Crystal Methamphetamine Use in Sexual Settings Among German Men Who Have Sex with Men. Front. Psychiatry 2019, 10, 886. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.; Tiwari, S.K.; Agrawal, K.; Hui, H.; Qin, Y.; Rana, T.M. Glial cell diversity and methamphetamine-induced neuroinflammation in human cerebral organoids. Mol. Psychiatry 2021, 26, 1194–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.M.; Compton, W.M.; Mustaquim, D. Patterns and Characteristics of Methamphetamine Use Among Adults—United States, 2015–2018. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.B.; Kepler, K.L.; Irvine, M.K.; Thomas, J.A. Associations between drug use patterns and viral load suppression among HIV-positive individuals who use support services in New York City. Drug Alcohol Depend. 2019, 197, 15–21. [Google Scholar] [CrossRef] [PubMed]
- de Guglielmo, G.; Fu, Y.; Chen, J.; Larrosa, E.; Hoang, I.; Kawamura, T.; Lorrai, I.; Zorman, B.; Bryant, J.; George, O.; et al. Increases in compulsivity, inflammation, and neural injury in HIV transgenic rats with escalated methamphetamine self-administration under extended-access conditions. Brain Res. 2020, 1726, 146502. [Google Scholar] [CrossRef]
- Ladak, F.; Socias, E.; Nolan, S.; Dong, H.; Kerr, T.; Wood, E.; Montaner, J.; Milloy, M.-J. Substance use patterns and HIV-1 RNA viral load rebound among HIV-Positive illicit drug users in a Canadian setting. Antivir. Ther. 2018, 24, 19–25. [Google Scholar] [CrossRef]
- Moore, D.J.; Blackstone, K.; Woods, S.P.; Ellis, R.; Atkinson, J.H.; Heaton, R.K.; Grant, I.; the HNRC Group; the TMARC Group. Methamphetamine use and neuropsychiatric factors are associated with antiretroviral non-adherence. AIDS Care 2012, 24, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Lai, H.-H.; Kuo, Y.-C.; Kuo, C.-J.; Lai, Y.-J.; Chen, M.; Chen, Y.-T.; Chen, C.-C.; Yen, M.-Y.; Hu, B.-S.; Wang, T.-H.; et al. Methamphetamine Use Associated with Non-adherence to Antiretroviral Treatment in Men Who Have Sex with Men. Sci. Rep. 2020, 10, 7131. [Google Scholar] [CrossRef]
- Marcondes, M.C.G.; Flynn, C.; Watry, D.D.; Zandonatti, M.; Fox, H.S. Methamphetamine Increases Brain Viral Load and Activates Natural Killer Cells in Simian Immunodeficiency Virus-Infected Monkeys. Am. J. Pathol. 2010, 177, 355–361. [Google Scholar] [CrossRef]
- Anderson, M.R.; Kashanchi, F.; Jacobson, S. Exosomes in Viral Disease. Neurotherapeutics 2016, 13, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Pleet, M.L.; Erickson, J.; DeMarino, C.; A Barclay, R.; Cowen, M.; Lepene, B.; Liang, J.; Kuhn, J.H.; Prugar, L.; Stonier, S.W.; et al. Ebola Virus VP40 Modulates Cell Cycle and Biogenesis of Extracellular Vesicles. J. Infect. Dis. 2018, 218, S365–S387. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, J.; Aslan, C.; Ahmadi, M.; Zolbanin, N.M.; Kashanchi, F.; Jafari, R. The versatile role of exosomes in human retroviral infections: From immunopathogenesis to clinical application. Cell Biosci. 2021, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, C.; Li, R.; Chen, H.; Chen, D.; Li, W. Exosomes in HIV infection. Curr. Opin. HIV AIDS 2021, 16, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Arthos, J.; Cicala, C.; Martinelli, E.; Macleod, K.; Van Ryk, D.; Wei, D.; Xiao, Z.; Veenstra, T.D.; Conrad, T.P.; Lempicki, R.A.; et al. HIV-1 envelope protein binds to and signals through integrin alpha4beta7, the gut mucosal homing receptor for peripheral T cells. Nat. Immunol. 2008, 9, 301–309. [Google Scholar] [CrossRef]
- Chand, S.; Messina, E.L.; AlSalmi, W.; Ananthaswamy, N.; Gao, G.; Uritskiy, G.; Padilla-Sanchez, V.; Mahalingam, M.; Peachman, K.K.; Robb, M.L.; et al. Glycosylation and oligomeric state of envelope protein might influence HIV-1 virion capture by alpha4beta7 integrin. Virology 2017, 508, 199–212. [Google Scholar] [CrossRef]
- Arakelyan, A.; Fitzgerald, W.; Zicari, S.; Vanpouille, C.; Margolis, L. Extracellular vesicles carry HIV Env and facilitate Hiv infection of human lymphoid tissue. Sci. Rep. 2017, 7, 1695. [Google Scholar] [CrossRef] [Green Version]
- Barclay, R.A.; Mensah, G.A.; Cowen, M.; DeMarino, C.; Kim, Y.; Pinto, D.O.; Erickson, J.; Kashanchi, F. Extracellular Vesicle Activation of Latent HIV-1 Is Driven by EV-Associated c-Src and Cellular SRC-1 via the PI3K/AKT/mTOR Pathway. Viruses 2020, 12, 665. [Google Scholar] [CrossRef]
- Sampey, G.C.; Meyering, S.S.; Zadeh, M.A.; Saifuddin, M.; Hakami, R.M.; Kashanchi, F. Exosomes and their role in CNS viral infections. J. Neuro Virol. 2014, 20, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Barclay, R.A.; Schwab, A.; DeMarino, C.; Akpamagbo, Y.; Lepene, B.; Kassaye, S.; Iordanskiy, S.; Kashanchi, F. Exosomes from uninfected cells activate transcription of latent HIV-1. J. Biol. Chem. 2017, 292, 11682–11701. [Google Scholar] [CrossRef] [Green Version]
- Marques de Menezes, E.G.; Jang, K.; George, A.F.; Nyegaard, M.; Neidleman, J.; Inglis, H.C.; Danesh, A.; Deng, X.; Afshari, A.; Kim, Y.H.; et al. Seminal Plasma-Derived Extracellular-Vesicle Fractions from HIV-Infected Men Exhibit Unique MicroRNA Signatures and Induce a Proinflammatory Response in Cells Isolated from the Female Reproductive Tract. J. Virol. 2020, 94, 16. [Google Scholar] [CrossRef]
- Olivetta, E.; Chiozzini, C.; Arenaccio, C.; Manfredi, F.; Ferrantelli, F.; Federico, M. Extracellular vesicle-mediated intercellular communication in HIV-1 infection and its role in the reservoir maintenance. Cytokine Growth Factor Rev. 2019, 51, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, V.; Fitzgerald, W.; Molodtsov, I.; Margolis, L. Persistent Immune Activation in HIV-1–Infected Ex Vivo Model Tissues Subjected to Antiretroviral Therapy: Soluble and Extracellular Vesicle-Associated Cytokines. JAIDS J. Acquir. Immune Defic. Syndr. 2020, 84, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Duette, G.; Gerber, P.P.; Rubione, J.; Perez, P.S.; Landay, A.L.; Crowe, S.M.; Liao, Z.; Witwer, K.W.; Holgado, M.P.; Salido, J.; et al. Induction of HIF-1α by HIV-1 Infection in CD4 + T Cells Promotes Viral Replication and Drives Extracellular Vesicle-Mediated Inflammation. mBio 2018, 9, e00757-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dargent, J.L.; Lespagnard, L.; Kornreich, A.; Hermans, P.; Clumeck, N.; Verhest, A. HIV-associated multinucleated giant cells in lymphoid tissue of the Waldeyer’s ring: A detailed study. Mod. Pathol. 2000, 13, 1293–1299. [Google Scholar] [CrossRef] [Green Version]
- Ryzhova, E.V.; Crinoa, P.; Shawvera, L.; Westmoreland, S.V.; Lackner, A.A.; González-Scarano, F. Simian Immunodeficiency Virus Encephalitis: Analysis of Envelope Sequences from Individual Brain Multinucleated Giant Cells and Tissue Samples. Virology 2002, 297, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, S.S.; Wenig, B.M.; Burke, A.P.; Mannan, P.; Thompson, L.D.R.; Abbondanzo, S.L.; Nelson, A.M.; Pope, M.; Steinman, R.M. Replication of HIV-1 in Dendritic Cell-Derived Syncytia at the Mucosal Surface of the Adenoid. Science 1996, 272, 115–117. [Google Scholar] [CrossRef]
- Najera, J.A.; Bustamante, E.A.; Bortell, N.; Morsey, B.; Fox, H.S.; Ravasi, T.; Marcondes, M.C.G. Methamphetamine abuse affects gene expression in brain-derived microglia of SIV-infected macaques to enhance inflammation and promote virus targets. BMC Immunol. 2016, 17, 7. [Google Scholar] [CrossRef] [Green Version]
- DeMarino, C.; Pleet, M.L.; Cowen, M.; Barclay, R.A.; Akpamagbo, Y.; Erickson, J.; Ndembi, N.; Charurat, M.; Jumare, J.; Bwala, S.; et al. Publisher Correction: Antiretroviral Drugs Alter the Content of Extracellular Vesicles from HIV-1-Infected Cells. Sci. Rep. 2018, 8, 14303. [Google Scholar] [CrossRef]
- Kim, Y.; Mensah, G.; Al Sharif, S.; Pinto, D.; Branscome, H.; Yelamanchili, S.; Cowen, M.; Erickson, J.; Khatkar, P.; Mahieux, R.; et al. Extracellular Vesicles from Infected Cells Are Released Prior to Virion Release. Cells 2021, 10, 781. [Google Scholar] [CrossRef]
- Jaworski, E.; Saifuddin, M.; Sampey, G.; Shafagati, N.; Van Duyne, R.; Iordanskiy, S.; Kehn-Hall, K.; Liotta, L.; Petricoin, E.; Young, M.; et al. The Use of Nanotrap Particles Technology in Capturing HIV-1 Virions and Viral Proteins from Infected Cells. PLoS ONE 2014, 9, e96778. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.; Meays, B.M.; Madduri, L.S.V.; Shahjin, F.; Chand, S.; Niu, M.; Albahrani, A.; Guda, C.; Pendyala, G.; Fox, H.S.; et al. Downregulation of an Evolutionary Young miR-1290 in an iPSC-Derived Neural Stem Cell Model of Autism Spectrum Disorder. Stem Cells Int. 2019, 2019, 8710180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chand, S.; Jo, A.; Vellichirammal, N.N.; Gowen, A.; Guda, C.; Schaal, V.; Odegaard, K.; Lee, H.; Pendyala, G.; Yelamanchili, S.V. Comprehensive Characterization of Nanosized Extracellular Vesicles from Central and Peripheral Organs: Implications for Preclinical and Clinical Applications. ACS Appl. Nano Mater. 2020, 3, 8906–8919. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; DeMarino, C.; Pleet, M.L.; Cowen, M.; Branscome, H.; Al Sharif, S.; Jones, J.; Dutartre, H.; Lepene, B.; Liotta, L.A.; et al. HTLV-1 Extracellular Vesicles Promote Cell-to-Cell Contact. Front. Microbiol. 2019, 10, 2147. [Google Scholar] [CrossRef]
- Yelamanchili, S.V.; Lamberty, B.G.; Rennard, D.A.; Morsey, B.M.; Hochfelder, C.G.; Meays, B.M.; Levy, E.; Fox, H.S. MiR-21 in Extracellular Vesicles Leads to Neurotoxicity via TLR7 Signaling in SIV Neurological Disease. PLoS Pathog. 2015, 11, e1005032. [Google Scholar]
- Harrison, E.B.; Hochfelder, C.G.; Lamberty, B.G.; Meays, B.M.; Morsey, B.M.; Kelso, M.L.; Fox, H.S.; Yelamanchili, S.V. Traumatic brain injury increases levels of miR-21 in extracellular vesicles: Implications for neuroinflammation. FEBS Open Bio 2016, 6, 835–846. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Wang, X.; Chen, H.; Song, L.; Ye, L.; Wang, S.-H.; Wang, Y.-J.; Zhou, L.; Ho, W.-Z. Methamphetamine Enhances HIV Infection of Macrophages. Am. J. Pathol. 2008, 172, 1617–1624. [Google Scholar] [CrossRef]
- Potula, R.; Persidsky, Y. Adding Fuel to the Fire: Methamphetamine Enhances HIV Infection. Am. J. Pathol. 2008, 172, 1467–1470. [Google Scholar] [CrossRef] [Green Version]
- Nair, M.P.N.; Saiyed, Z.M.; Nair, N.; Gandhi, N.H.; Rodriguez, J.W.; Boukli, N.; Provencio-Vásquez, E.; Malow, R.M.; Míguez-Burbano, M.J. Methamphetamine Enhances HIV-1 Infectivity in Monocyte Derived Dendritic Cells. J. Neuroimmune Pharmacol. 2008, 4, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Kadiu, I.; Narayanasamy, P.; Dash, P.K.; Zhang, W.; Gendelman, H.E. Biochemical and Biologic Characterization of Exosomes and Microvesicles as Facilitators of HIV-1 Infection in Macrophages. J. Immunol. 2012, 189, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Chand, S.; Gowen, A.; Savine, M.; Moore, D.; Clark, A.; Huynh, W.; Wu, N.; Odegaard, K.; Weyrich, L.; Bevins, R.A.; et al. A comprehensive study to delineate the role of an extracellular vesicle-associated microRNA-29a in chronic methamphetamine use disorder. J. Extracell. Vesicles 2021, 10, e12177. [Google Scholar] [CrossRef]
- Poli, G. U1 and OM10.1. Myeloid Cell Lines as Surrogate Models of Reversible Proviral Latency. Methods Pharmacol. Toxicol. 2022, 2407, 17–28. [Google Scholar] [CrossRef]
- Ash, M.; Al-Harthi, L.; Schneider, J. HIV in the Brain: Identifying Viral Reservoirs and Addressing the Challenges of an HIV Cure. Vaccines 2021, 9, 867. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.E.; Sneider, A.; Witwer, K.W.; Bergese, P.; Bhattacharyya, S.N.; Cocks, A.; Cocucci, E.; Erdbrügger, U.; Falcon-Perez, J.M.; Freeman, D.W.; et al. Biological membranes in EV biogenesis, stability, uptake, and cargo transfer: An ISEV position paper arising from the ISEV membranes and EVs workshop. J. Extracell. Vesicles 2019, 8, 1684862. [Google Scholar] [CrossRef] [Green Version]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neuro-Oncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Teng, F.; Fussenegger, M. Shedding Light on Extracellular Vesicle Biogenesis and Bioengineering. Adv. Sci. 2021, 8, 2003505. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Latifkar, A.; Hur, Y.H.; Sanchez, J.C.; Cerione, R.A.; Antonyak, M.A. New insights into extracellular vesicle biogenesis and function. J. Cell Sci. 2019, 132, jcs222406. [Google Scholar] [CrossRef] [Green Version]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT Pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Babst, M. MVB vesicle formation: ESCRT-dependent, ESCRT-independent and everything in between. Curr. Opin. Cell Biol. 2011, 23, 452–457. [Google Scholar] [CrossRef] [Green Version]
- Harris, D.S.; Boxenbaum, H.; Everhart, E.T.; Ms, G.S.; Mendelson, J.E.; Jones, R.T. The bioavailability of intranasal and smoked methamphetamine. Clin. Pharmacol. Ther. 2003, 74, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Moita, C.; van Niel, G.; Kowal, J.; Vigneron, J.; Benaroch, P.; Manel, N.; Moita, L.F.; Théry, C.; Raposo, G. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. 2013, 126 Pt 24, 5553–5565. [Google Scholar] [CrossRef] [Green Version]
- Arenaccio, C.; Anticoli, S.; Manfredi, F.; Chiozzini, C.; Olivetta, E.; Federico, M. Latent HIV-1 is activated by exosomes from cells infected with either replication-competent or defective HIV-1. Retrovirology 2015, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenaccio, C.; Chiozzini, C.; Columba-Cabezas, S.; Manfredi, F.; Federico, M. Cell activation and HIV-1 replication in unstimulated CD4+T lymphocytes ingesting exosomes from cells expressing defective HIV-1. Retrovirology 2014, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patters, B.J.; Kumar, S. The role of exosomal transport of viral agents in persistent HIV pathogenesis. Retrovirology 2018, 15, 79. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Place, A.T.; Chen, Z.; Brovkovych, V.M.; Vogel, S.M.; Muller, W.A.; Skidgel, R.A.; Malik, A.B.; Minshall, R.D. ICAM-1–activated Src and eNOS signaling increase endothelial cell surface PECAM-1 adhesivity and neutrophil transmigration. Blood 2012, 120, 1942–1952. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, J.C.; Starkey, J.; Zhang, D.; Sarfo, A.; Chadha, P.; Wills, J.W.; Han, J. Glycoprotein D of HSV-1 is dependent on tegument protein UL16 for packaging and contains a motif that is differentially required for syncytia formation. Virology 2019, 527, 64–76. [Google Scholar] [CrossRef]
- Burton, C.; Bartee, E. Syncytia Formation in Oncolytic Virotherapy. Mol. Ther. Oncolytics 2019, 15, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Compton, A.A.; Schwartz, O. They Might Be Giants: Does Syncytium Formation Sink or Spread HIV Infection? PLOS Pathog. 2017, 13, e1006099. [Google Scholar] [CrossRef]
- Higuchi, H.; Bronk, S.F.; Bateman, A.; Harrington, K.; Vile, R.G.; Gores, G.J. Viral fusogenic membrane glycoprotein expression causes syncytia formation with bioenergetic cell death: Implications for gene therapy. Cancer Res. 2000, 60, 6396–6402. [Google Scholar]
- Hassan, J.; Browne, K.; De Gascun, C. HIV-1 in Monocytes and Macrophages: An Overlooked Reservoir? Viral Immunol. 2016, 29, 532–533. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.E.; Jaworowski, A.; Hearps, A.C. The HIV Reservoir in Monocytes and Macrophages. Front. Immunol. 2019, 10, 1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiner, B.C.; Keblesh, J.P.; Xiong, H. Methamphetamine abuse, HIV infection, and neurotoxicity. Int. J. Physiol. Pathophysiol. Pharmacol. 2009, 1, 162–179. [Google Scholar] [PubMed]
- Henne, W.M.; Stenmark, H.; Emr, S.D. Molecular Mechanisms of the Membrane Sculpting ESCRT Pathway. Cold Spring Harb. Perspect. Biol. 2013, 5, a016766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Tang, S.; Han, X.; Jiang, Z.; Dong, L.; Liu, C.; Liang, X.; Dong, J.; Qiu, C.; Wang, Y.; et al. KIBRA controls exosome secretion via inhibiting the proteasomal degradation of Rab27a. Nat. Commun. 2019, 10, 1639. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Overbye, A.; Brech, A.; Torgersen, M.L.; Jakobsen, I.S.; Sandvig, K.; Llorente, A. PIKfyve inhibition increases exosome release and induces secretory autophagy. Cell Mol. Life Sci. 2016, 73, 4717–4737. [Google Scholar] [CrossRef]
- Kajimoto, T.; Okada, T.; Miya, S.; Zhang, L.; Nakamura, S.-I. Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes. Nat. Commun. 2013, 4, 2712. [Google Scholar] [CrossRef] [Green Version]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brügger, B.; Simons, M. Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, D.; Jin, F.; Bian, Z.; Li, L.; Liang, H.; Li, M.; Shi, L.; Pan, C.; Zhu, D.; et al. Pyruvate kinase type M2 promotes tumour cell exosome release via phosphorylating synaptosome-associated protein 23. Nat. Commun. 2017, 8, 14041. [Google Scholar] [CrossRef] [Green Version]
- Baixauli, F.; Lopez-Otin, C.; Mittelbrunn, M. Exosomes and Autophagy: Coordinated Mechanisms for the Maintenance of Cellular Fitness. Front. Immunol. 2014, 5, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Deep, G. Hypoxia in tumor microenvironment regulates exosome biogenesis: Molecular mechanisms and translational opportunities. Cancer Lett. 2020, 479, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Kita, S.; Koyama, Y.; Fukuda, S.; Takeda, H.; Takahashi, M.; Fujishima, Y.; Nagao, H.; Masuda, S.; Tanaka, Y.; et al. Adiponectin/T-cadherin system enhances exosome biogenesis and decreases cellular ceramides by exosomal release. JCI Insight 2018, 3, e99680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astarita, G.; Avanesian, A.; Grimaldi, B.; Realini, N.; Justinova, Z.; Panlilio, L.V.; Basit, A.; Goldberg, S.R.; Piomelli, D. Methamphetamine Accelerates Cellular Senescence through Stimulation of De Novo Ceramide Biosynthesis. PLoS ONE 2015, 10, e0116961. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Zhang, G.; Han, C.; Ma, K.; Guo, X.; Wan, F.; Kou, L.; Yin, S.; Liu, L.; Huang, J.; et al. Microglia as modulators of exosomal alpha-synuclein transmission. Cell Death Dis. 2019, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Croese, T.; Furlan, R. Extracellular vesicles in neurodegenerative diseases. Mol. Asp. Med. 2018, 60, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Glass, J.D.; Fedor, H.; Wesselingh, S.L.; McArthur, J.C. Immunocytochemical quantitation of human immunodeficiency virus in the brain: Correlations with dementia. Ann. Neurol. 1995, 38, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Borgmann, K.; Ghorpade, A. HIV-1, methamphetamine and astrocytes at neuroinflammatory Crossroads. Front. Microbiol. 2015, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, A.; Collman, R.G. CNS Inflammation and Macrophage/Microglial Biology Associated with HIV-1 Infection. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2009, 4, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Sami Saribas, A.; Cicalese, S.; Ahooyi, T.M.; Khalili, K.; Amini, S.; Sariyer, I.K. HIV-1 Nef is released in extracellular vesicles derived from astrocytes: Evidence for Nef-mediated neurotoxicity. Cell Death Dis. 2017, 8, e2542. [Google Scholar] [CrossRef] [Green Version]
- Lopez, P.; Koh, W.H.; Hnatiuk, R.; Murooka, T.T. HIV Infection Stabilizes Macrophage-T Cell Interactions To Promote Cell-Cell HIV Spread. J. Virol. 2019, 93, e00805-19. [Google Scholar] [CrossRef] [Green Version]
- Verollet, C.; Cabec, V.E.; Emaridonneau-Parini, I. HIV-1 Infection of T Lymphocytes and Macrophages Affects Their Migration via Nef. Front. Immunol. 2015, 6, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallet, C.; De Rovere, M.; Van Assche, J.; Daouad, F.; De Wit, S.; Gautier, V.; Mallon, P.W.G.; Marcello, A.; Van Lint, C.; Rohr, O.; et al. Microglial Cells: The Main HIV-1 Reservoir in the Brain. Front. Cell. Infect. Microbiol. 2019, 9, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, A.; Kang, G.; Hattler, J.B.; Galadima, H.; Zhang, J.; Li, Q.; Kim, W.-K. Macrophages but not Astrocytes Harbor HIV DNA in the Brains of HIV-1-Infected Aviremic Individuals on Suppressive Antiretroviral Therapy. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2019, 14, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saylor, D.; Dickens, A.; Sacktor, N.; Haughey, N.; Slusher, B.; Pletnikov, M.; Mankowski, J.L.; Brown, A.; Volsky, D.J.; McArthur, D. HIV-associated neurocognitive disorder—Pathogenesis and prospects for treatment. Nat. Rev. Neurol. 2016, 12, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Gannon, P.; Khan, M.Z.; Kolson, D.L. Current understanding of HIV-associated neurocognitive disorders pathogenesis. Curr. Opin. Neurol. 2011, 24, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaul, M.; Zheng, J.; Okamoto, S.; Gendelman, H.E.; Lipton, S.A. HIV-1 infection and AIDS: Consequences for the central nervous system. Cell Death Differ. 2005, 12 (Suppl. 1), 878–892. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chand, S.; DeMarino, C.; Gowen, A.; Cowen, M.; Al-Sharif, S.; Kashanchi, F.; Yelamanchili, S.V. Methamphetamine Induces the Release of Proadhesive Extracellular Vesicles and Promotes Syncytia Formation: A Potential Role in HIV-1 Neuropathogenesis. Viruses 2022, 14, 550. https://0-doi-org.brum.beds.ac.uk/10.3390/v14030550
Chand S, DeMarino C, Gowen A, Cowen M, Al-Sharif S, Kashanchi F, Yelamanchili SV. Methamphetamine Induces the Release of Proadhesive Extracellular Vesicles and Promotes Syncytia Formation: A Potential Role in HIV-1 Neuropathogenesis. Viruses. 2022; 14(3):550. https://0-doi-org.brum.beds.ac.uk/10.3390/v14030550
Chicago/Turabian StyleChand, Subhash, Catherine DeMarino, Austin Gowen, Maria Cowen, Sarah Al-Sharif, Fatah Kashanchi, and Sowmya V. Yelamanchili. 2022. "Methamphetamine Induces the Release of Proadhesive Extracellular Vesicles and Promotes Syncytia Formation: A Potential Role in HIV-1 Neuropathogenesis" Viruses 14, no. 3: 550. https://0-doi-org.brum.beds.ac.uk/10.3390/v14030550