
Streptococcus thermophilus Phages in Whey Derivatives: From Problem to Application in the Dairy Industry


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Phage Isolation
2.3. Phage Characterization
2.3.1. Host Range
2.3.2. DNA Extraction and Restriction Enzyme Analysis
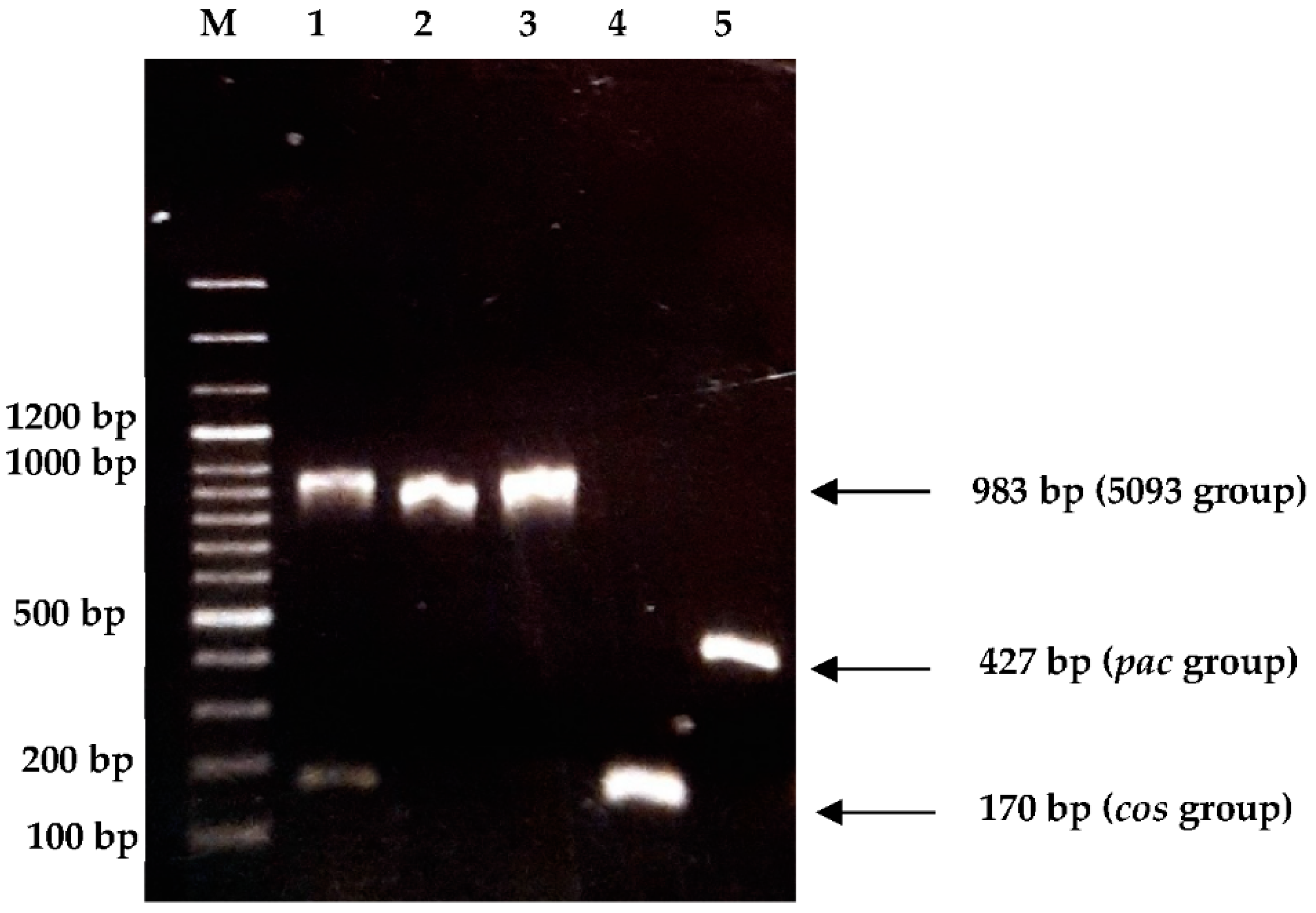
2.3.3. Determination of Genetic Group
2.3.4. Thermal Resistance
3. Results
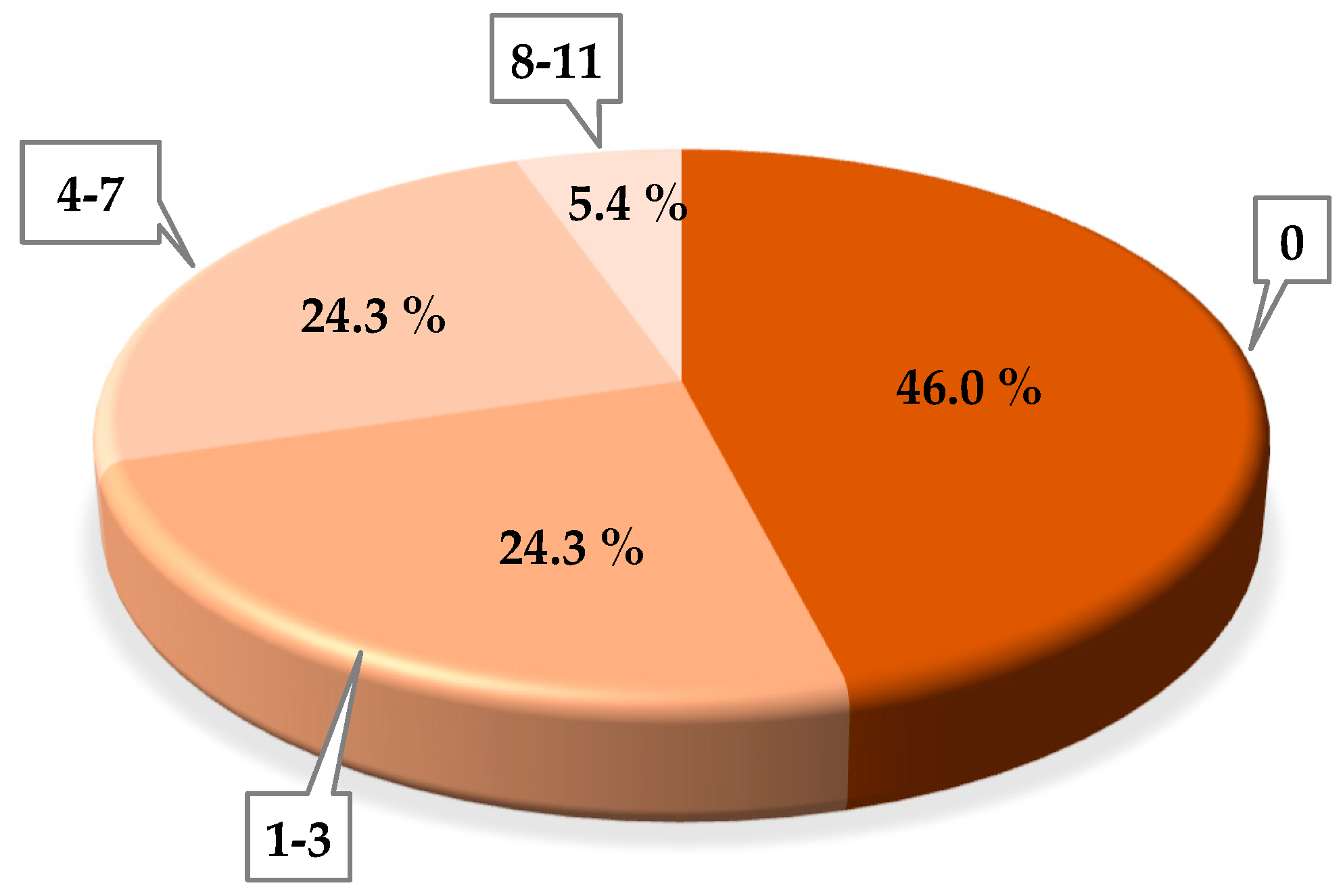
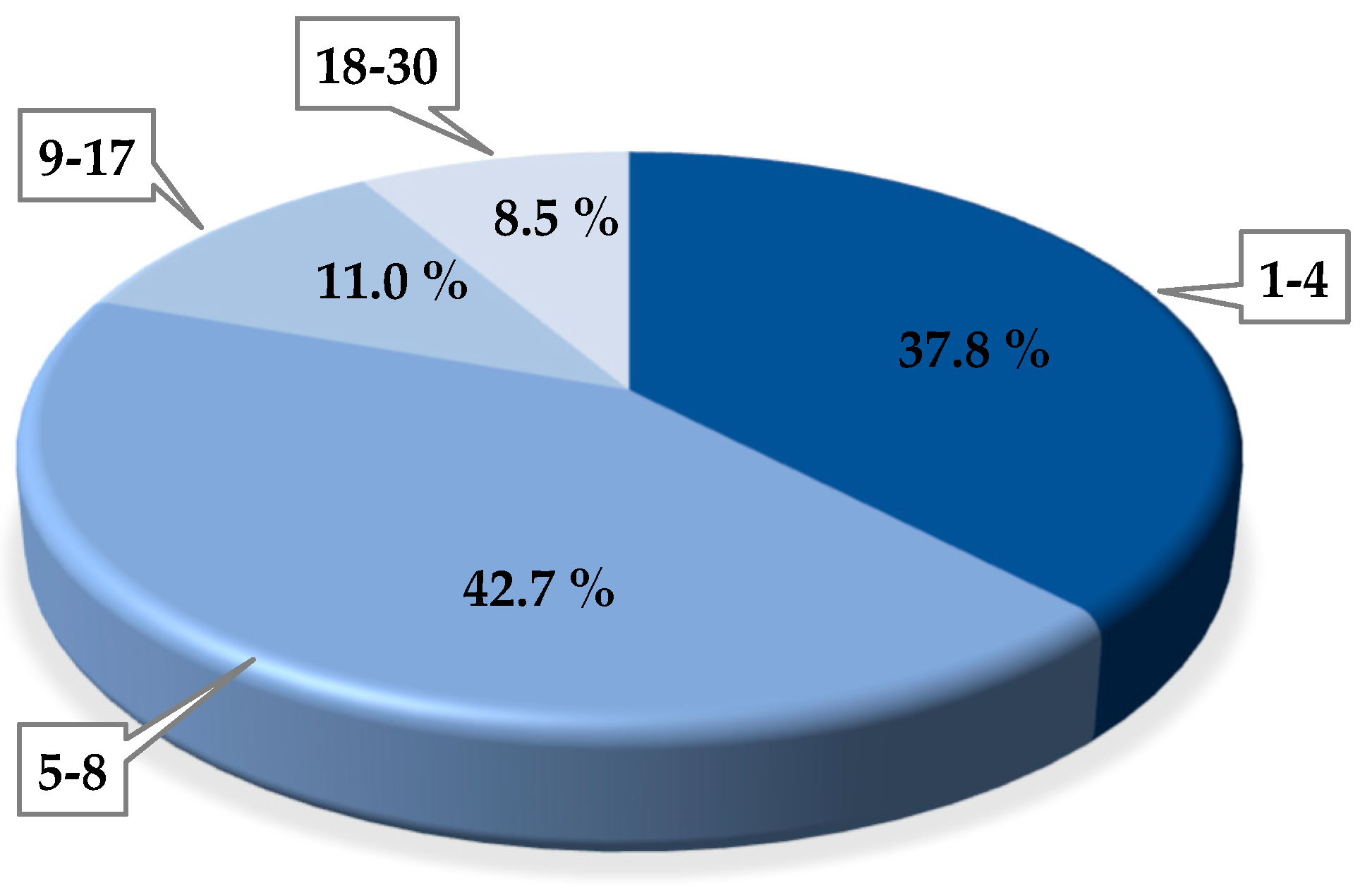
3.1. Phage Isolation
3.2. Phage Concentration
3.3. Phage Characterization
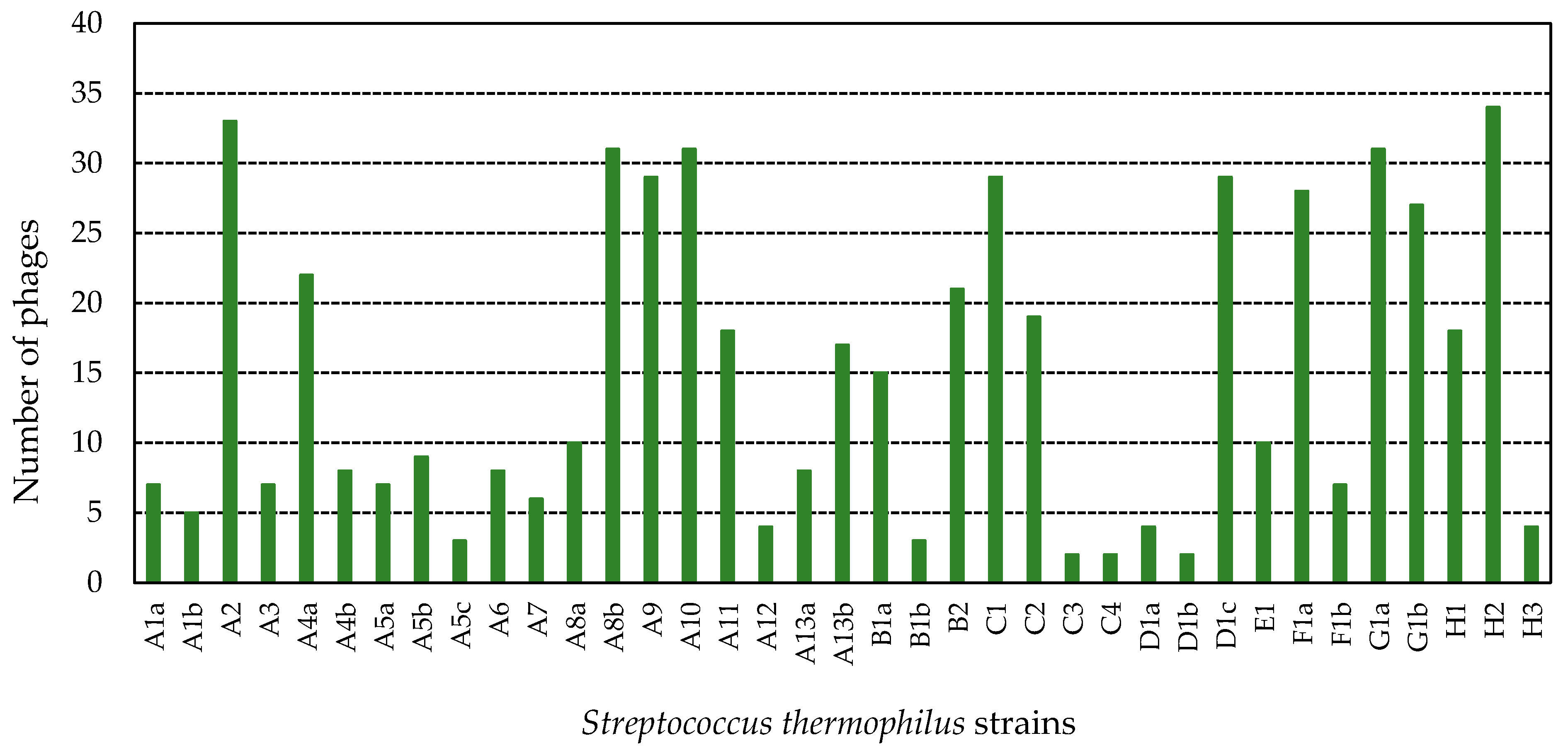
3.3.1. Host Range
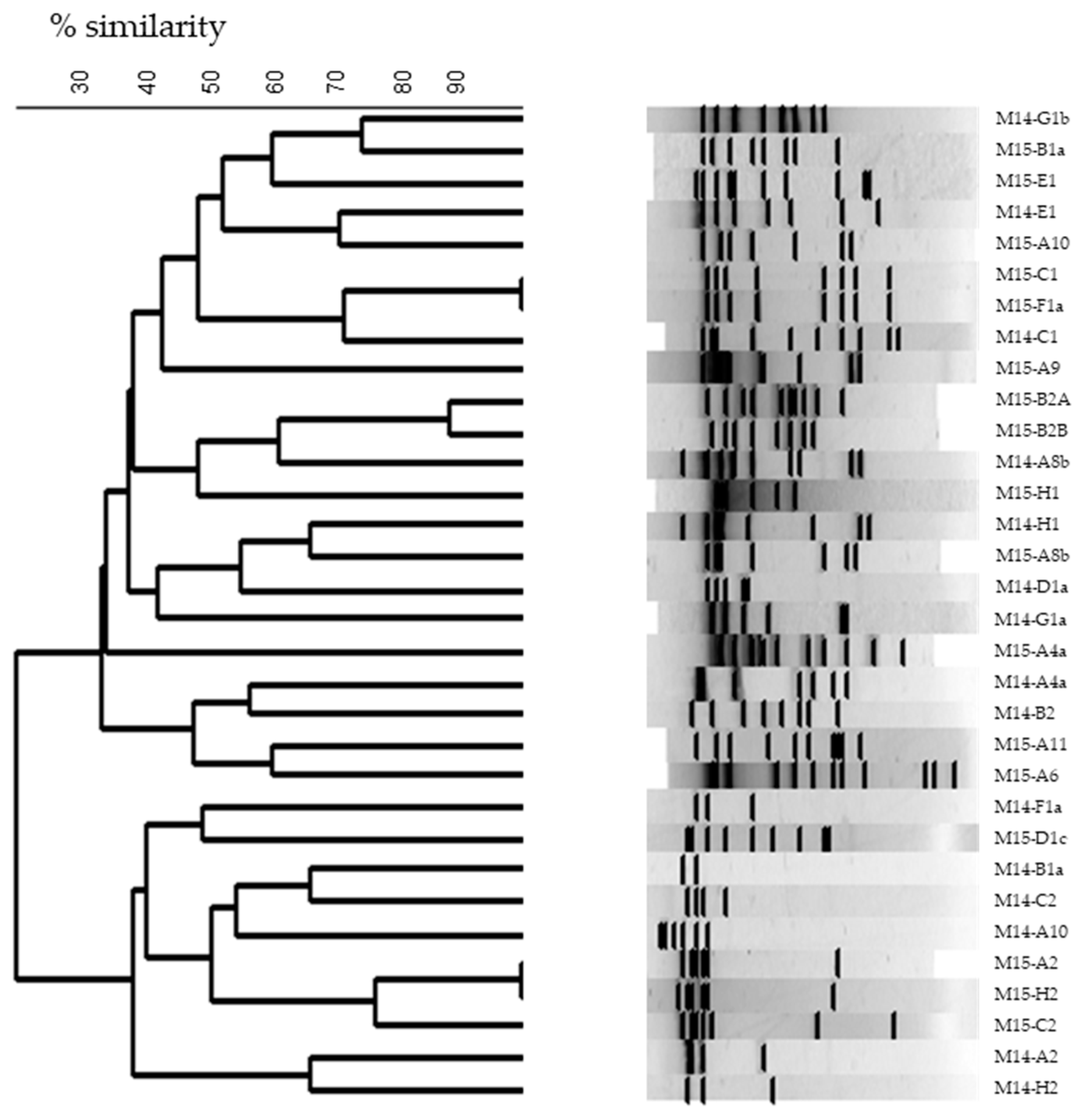
3.3.2. Restriction Enzyme Analysis
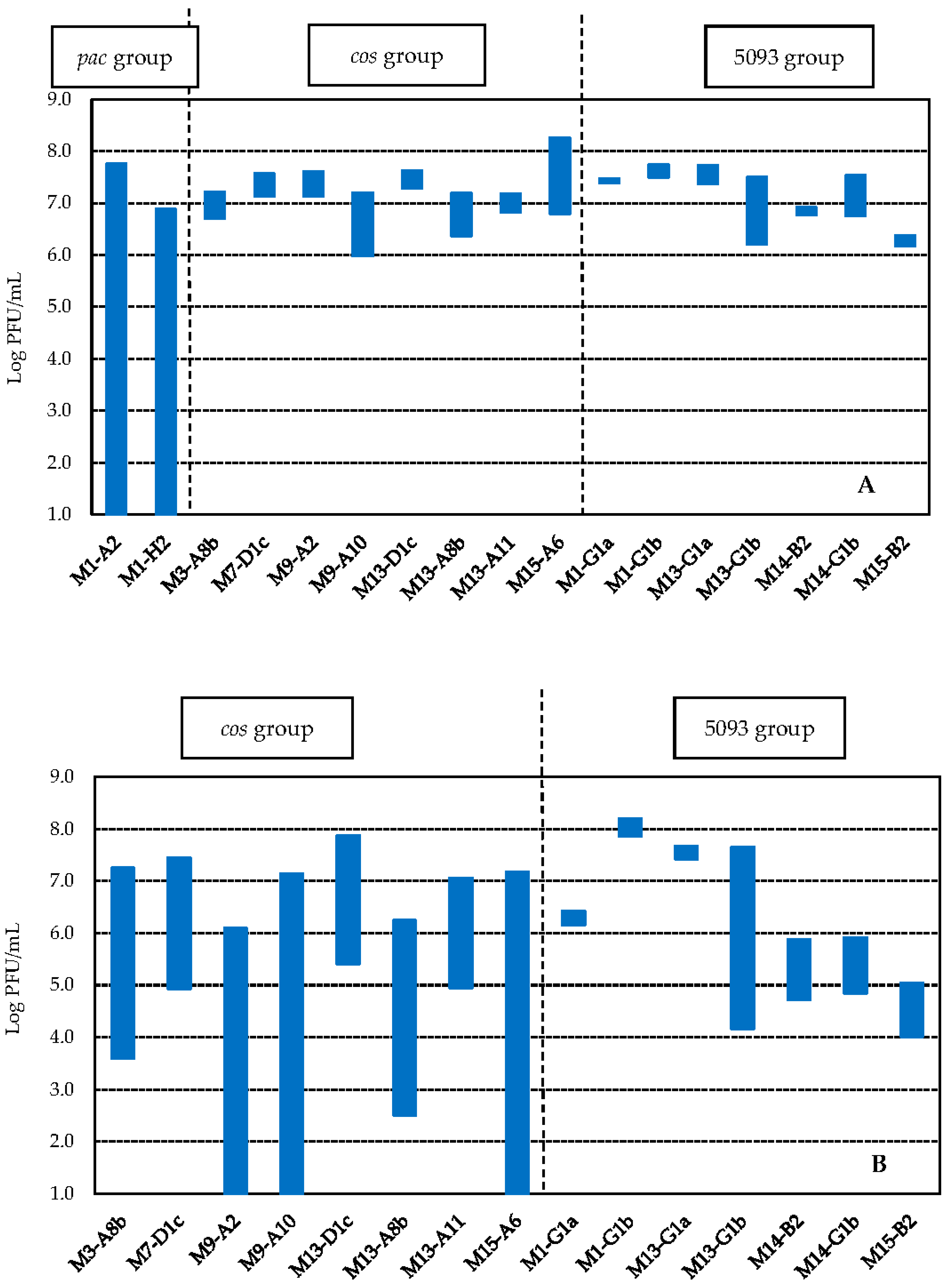
3.3.3. Determination of Genetic Group
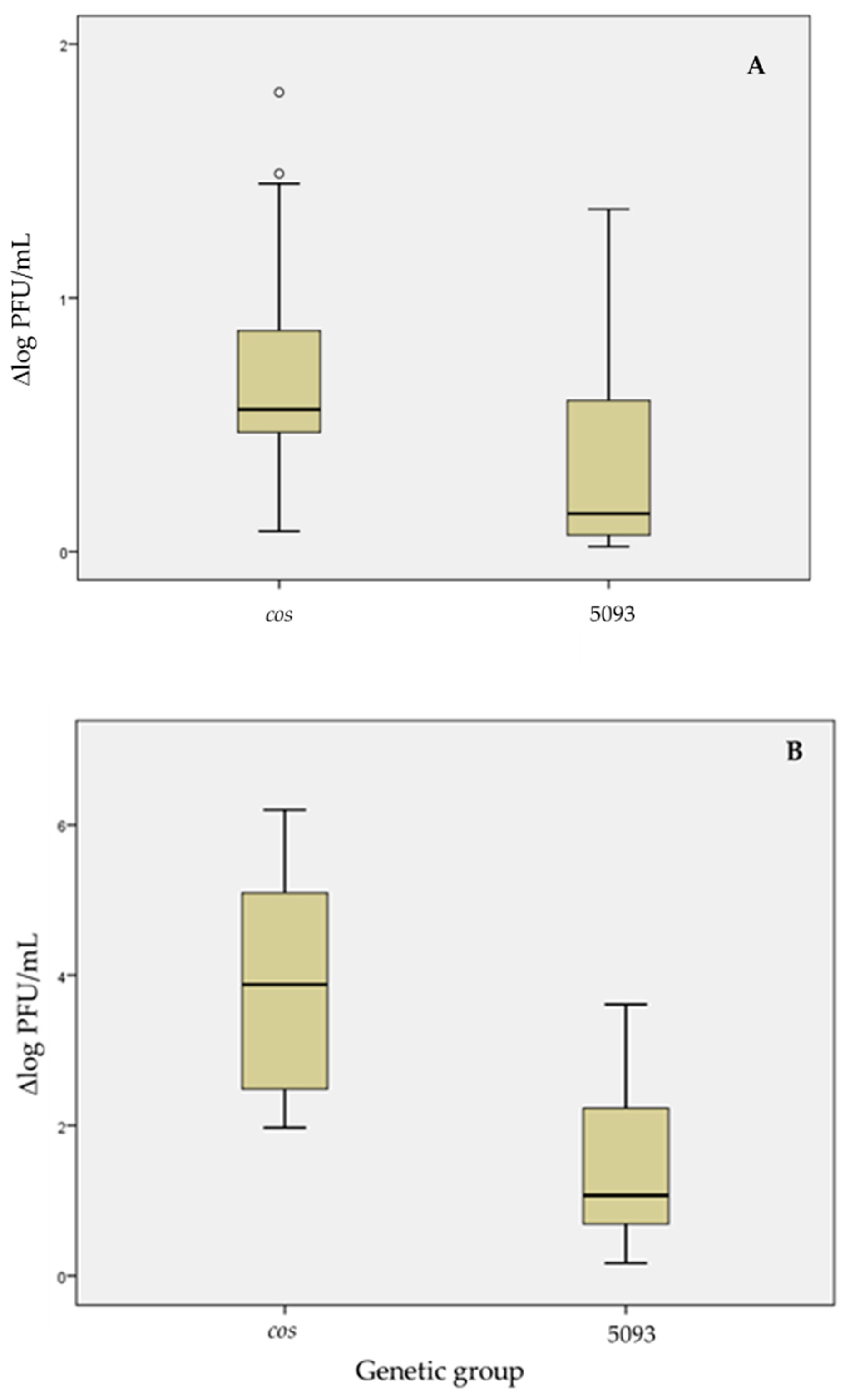
3.3.4. Thermal Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Yadav, J.S.S.; Yan, S.; Pilli, S.; Kumar, L.; Tyagi, R.D.; Surampalli, R.Y. Cheese whey: A potential resource to transform into bioprotein, functional/nutritional proteins and bioactive peptides. Biotechnol. Adv. 2015, 33, 756–774. [Google Scholar] [CrossRef] [PubMed]
- Masotti, F.; Cattaneo, S.; Stuknytė, M.; De Noni, I. Technological tools to include whey proteins in cheese: Current status and perspectives. Trends Food Sci. Technol. 2017, 64, 102–114. [Google Scholar] [CrossRef]
- Atamer, Z.; Meike, S.; Neve, H.; Heller, K.J.; Hinrichs, J. Review: Elimination of bacteriophages in whey and whey products. Front. Microbiol. 2013, 4, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, N.; Brinks, E.; Samtlebe, M.; Hinrichs, J.; Atamer, Z.; Kot, W.; Franz, C.M.A.P.; Neve, H.; Heller, K.J. Whey powders are a rich source and excellent storage matrix for dairy bacteriophages. Int. J. Food Microbiol. 2017, 241, 308–317. [Google Scholar] [CrossRef]
- Briggiler Marcó, M.; Suárez, V.B.; Quiberoni, A.; Pujato, S.A. Inactivation of dairy bacteriophages by thermal and chemical treatments. Viruses 2019, 11, 480. [Google Scholar] [CrossRef] [Green Version]
- Atamer, Z.; Ali, Y.; Neve, H.; Heller, K.J.; Hinrichs, J. Thermal resistance of bacteriophages attacking flavour-producing dairy Leuconostoc starter cultures. Int. Dairy J. 2011, 21, 327–333. [Google Scholar] [CrossRef]
- Lavelle, K.; Martinez, I.; Neve, H.; Lugli, G.A.; Franz, C.M.A.P.; Ventura, M.; Dal Bello, F.; van Sinderen, D.; Mahony, J. Biodiversity of Streptococcus thermophilus phages in global dairy fermentations. Viruses 2018, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Samtlebe, M.; Wagner, N.; Neve, H.; Heller, K.J.; Hinrichs, J.; Atamer, Z. Application of a membrane technology to remove bacteriophages from whey. Int. Dairy J. 2015, 48, 38–45. [Google Scholar] [CrossRef]
- Geagea, H.; Gomaa, A.I.; Remondetto, G.; Moineau, S.; Subirade, M. Investigation of the protective effect of whey proteins on lactococcal phages during heat treatment at various pH. Int. J. Food Microbiol. 2015, 210, 33–41. [Google Scholar] [CrossRef]
- Reinheimer, J.A.; Binetti, A.G.; Quiberoni, A.; Bailo, N.B.; Rubiolo, A.C.; Giraffa, G. Natural milk cultures for the production of Argentinian cheeses. J. Food Prot. 1997, 60, 59–63. [Google Scholar] [CrossRef]
- Svensson, U.; Christiansson, A. Methods for phage monitoring. FILIDF Bull. 1991, 263, 29–39. [Google Scholar]
- Neviani, E.; Carminati, D.; Giraffa, G. Selection of some bacteriophage and lysozyme-Resistant variants of Lactobacillus helveticus CNRZ 892. J. Dairy Sci. 1992, 75, 905–913. [Google Scholar] [CrossRef]
- Pujato, S.A.; Guglielmotti, D.M.; Ackermann, H.W.; Patrignani, F.; Lanciotti, R.; Reinheimer, J.A.; Quiberoni, A. Leuconostoc bacteriophages from blue cheese manufacture: Long-term survival, resistance to thermal treatments, high pressure homogenization and chemical biocides of industrial application. Int. J. Food Microbiol. 2014, 177, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Vauterin, L.; Vauterin, P. Computer-aided objective comparison of electrophoresis patterns for grouping and identification of microorganisms. Eur. Microbiol. 1992, 1, 37–42. [Google Scholar]
- Philippe, C.; Levesque, S.; Dion, M.B.; Tremblay, D.M.; Horvath, P.; Lüth, N.; Cambillau, C.; Franz, C.; Neve, H.; Fremaux, C.; et al. Novel genus of phages infecting Streptococcus thermophilus: Genomic and morphological characterization. Appl. Environ. Microbiol. 2020, 86, e00227-20. [Google Scholar] [CrossRef] [PubMed]
- Quiberoni, A.; Tremblay, D.; Ackermann, H.W.; Moineau, S.; Reinheimer, J.A. Diversity of Streptococcus thermophilus phages in a large-production cheese factory in Argentina. J. Dairy Sci. 2006, 89, 3791–3799. [Google Scholar] [CrossRef]
- McDonnell, B.; Mahony, J.; Hanemaaijer, L.; Neve, H.; Noben, J.P.; Lugli, G.A.; Ventura, M.; Kouwen, T.R.; van Sinderen, D. Global survey and genome exploration of bacteriophages infecting the lactic acid bacterium Streptococcus thermophilus. Front. Microbiol. 2017, 8, 1754. [Google Scholar] [CrossRef]
- Tayyarcan, E.K.; Acar Soykut, E.; Boyaci, I.H. A Raman-spectroscopy-based approach for detection and discrimination of Streptococcus thermophilus and Lactobacillus bulgaricus phages at low titer in raw milk. Folia Microbiol. 2018, 63, 627–636. [Google Scholar] [CrossRef]
- Moineau, S.; Lévesque, C. Control of Bacteriophages in Industrial Fermentations. In Bacteriophages: Biology and Applications; Kutter, E., Sulakvelidze, A., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 285–296. [Google Scholar]
- Atamer, Z.; Dietrich, J.; Müller-Merbach, M.; Neve, H.; Heller, K.J.; Hinrichs, J. Screening for and characterization of Lactococcus lactis bacteriophages with high thermal resistance. Int. Dairy J. 2009, 19, 228–235. [Google Scholar] [CrossRef]
- Murphy, J.; Royer, B.; Mahony, J.; Hoyles, L.; Heller, K.; Neve, H.; Bonestroo, M.; Nauta, A.; van Sinderen, D. Biodiversity of lactococcal bacteriophages isolated from 3 Gouda-type cheese-producing plants. J. Dairy Sci. 2013, 96, 4945–4957. [Google Scholar] [CrossRef] [Green Version]
- Lavelle, K.; Murphy, J.; Fitzgerald, B.; Lugli, G.A.; Zomer, A.; Neve, H.; Ventura, M.; Franz, C.M.; Cambillau, C.; van Sinderen, D.; et al. A decade of Streptococcus thermophilus phage evolution in an Irish dairy plant. Appl. Environ. Microbiol. 2018, 84, e02855-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, S.; Griffin, C.; O’Sullivan, O.; Coffey, A.; McAuliffe, O.E.; Meijer, W.C.; Serrano, L.M.; Ross, R.P. A new phage on the “Mozzarella” block: Bacteriophage 5093 shares a low level of homology with other Streptococcus thermophilus phages. Int. Dairy J. 2011, 21, 963–969. [Google Scholar] [CrossRef]
- McDonnell, B.; Mahony, J.; Neve, H.; Hanemaaijer, L.; Noben, J.P.; Kouwen, T.; van Sinderen, D. Identification and analysis of a novel group of bacteriophages infecting the lactic acid bacterium Streptococcus thermophilus. Appl. Environ. Microbiol. 2016, 82, 5153–5165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binetti, A.G.; Del Río, B.; Cruz Martín, M.; Álvarez, M.A. Detection and characterization of Streptococcus thermophilus bacteriophages by use of the antireceptor gene sequence. Appl. Environ. Microbiol. 2005, 71, 6096–6103. [Google Scholar] [CrossRef] [Green Version]
- Binetti, A.G.; Reinheimer, J.A. Thermal and chemical inactivation of indigenous Streptococcus thermophilus bacteriophages isolated from Argentinian dairy plants. J. Food Prot. 2000, 63, 509–515. [Google Scholar] [CrossRef]
- Capra, M.L.; Neve, H.; Sorati, P.C.; Atamer, Z.; Hinrichs, J.; Heller, K.J.; Quiberoni, A. Extreme thermal resistance of phages isolated from dairy samples: Updating traditional phage detection methodologies. Int. Dairy J. 2013, 30, 59–63. [Google Scholar] [CrossRef]
- Wagner, N.; Samtlebe, M.; Franz, C.M.A.P.; Neve, H.; Heller, K.J.; Hinrichs, J.; Atamer, Z. Dairy bacteriophages isolated from whey powder: Thermal inactivation and kinetic characterisation. Int. Dairy J. 2017, 68, 95–104. [Google Scholar] [CrossRef]
- Atamer, Z.; Dietrich, J.; Neve, H.; Heller, K.J.; Hinrichs, J. Influence of the suspension media on the thermal treatment of mesophilic lactococcal bacteriophages. Int. Dairy J. 2010, 20, 408–414. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Type d | Lysed Streptococcus thermophilus Strains (Number) | Range of Phage Concentration (PFU/g) |
|---|---|---|---|
| M1a | WPC 35 | 5 | <1.0 × 102–2.2 × 104 |
| M2 a | WPC 35 | 3 | <1.0 × 102–1.0 × 103 |
| M3 a | WPC 35 | 4 | <1.0 × 102–4.3 × 103 |
| M4 a | WPC 35 | 0 | nd |
| M5 a | WPC 35 | 1 | 5.0 × 103 |
| M6 a | WPC 35 | 2 | 3.5 × 103–2.6 × 104 |
| M7 a | WPC 35 | 4 | <1.0 × 102–1.5 × 103 |
| M8 a | WPC 35 | 8 | <1.0 × 102–1.6 × 105 |
| M9 a | WPC 35 | 7 | <1.0 × 102–3.3 × 105 |
| M10 a | WPC 35 | 3 | 1.0 × 102–5.0 × 102 |
| M11 a | WPC 35 | 0 | nd |
| M12 b | WPC 50 | 0 | nd |
| M13 a | WPC 80 | 18 | <1.0 × 102–1.1 × 105 |
| M14 c | WPC 35 | 15 | <1.0 × 102–7.0 × 104 |
| M15 c | WPC 35 | 17 | <1.0 × 102–1.9 × 104 |
| Genetic Group | Primer Name | Sequence | Expected Size | Reference |
|---|---|---|---|---|
| cos | cos FOR | 5′-ggttcacgtgtttatgaaaaatgg-3′ | 170 bp | [17] |
| cos REV | 5′-agcagaatcagcaagcaagctgtt-3′ | |||
| pac | pac FOR | 5′-gaagctatgcgtatgcaagt-3′ | 427 bp | [17] |
| pac REV | 5′-ttagggataagagtcaagtg-3′ | |||
| 5093 | 5093 FOR | 5′-ctggctcttggtggtcttgc-3′ | 983 bp | [18] |
| 5093 REV | 5′-gcggcaaccatcttagaccag-3′ | |||
| 987 | 987 FOR | 5′-ctaagcgtttgccactgtcag-3′ | 707 bp | [18] |
| 987 REV | 5′-gctgccgttgtttgaaaac-3′ |
| Genetic Group | Δlog PFU/mL * for Thermal Treatment ( ± δ): | |
|---|---|---|
| 75 °C for 5 min ** | 80 °C for 5 min ** | |
| cos | ||
| M3-A8b | 0.53 ± 0.04 a,b | 3.67 ± 0.09 b |
| M7-D1c | 0.44 ± 0.02 a | 2.52 ± 0.11 a |
| M9-A2 | 0.41 ± 0.34 a | 5.10 ± 0.02 c |
| M9-A10 | 1.38 ± 0.46 b,c | 5.22 ± 0.30 c |
| M13-D1c | 0.40 ± 0.28 a | 2.47 ± 0.06 a |
| M13-A8b | 0.84 ± 0.03 a,b,c | 4.19 ± 0.24 b |
| M13-A11 | 0.47 ± 0.19 a | 2.21 ± 0.35 a |
| M15-A6 | 1.47 ± 0.02 c | 6.18 ± 0.04 d |
| 5093 | ||
| M1-G1a | 0.07 ± 0.04 a | 0.28 ± 0.07 a |
| M1-G1b | 0.24 ± 0.13 a | 0.54 ± 0.23 a |
| M13-G1a | 0.26 ± 0.31 a | 1.29 ± 1.22 a |
| M13-G1b | 1.29 ± 0.08 b | 3.56 ± 0.06 b |
| M14-B2 | 0.09 ± 0.09 a | 0.86 ± 0.26 a |
| M14-G1b | 0.80 ± 0.11 b | 1.38 ± 0.51 a |
| M15-B2(B) | 0.14 ± 0.14 a | 1.12 ± 0.14 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briggiler Marcó, M.; Machado, N.; Quiberoni, A.; Suárez, V. Streptococcus thermophilus Phages in Whey Derivatives: From Problem to Application in the Dairy Industry. Viruses 2022, 14, 810. https://0-doi-org.brum.beds.ac.uk/10.3390/v14040810
Briggiler Marcó M, Machado N, Quiberoni A, Suárez V. Streptococcus thermophilus Phages in Whey Derivatives: From Problem to Application in the Dairy Industry. Viruses. 2022; 14(4):810. https://0-doi-org.brum.beds.ac.uk/10.3390/v14040810
Chicago/Turabian StyleBriggiler Marcó, Mariángeles, Nicolás Machado, Andrea Quiberoni, and Viviana Suárez. 2022. "Streptococcus thermophilus Phages in Whey Derivatives: From Problem to Application in the Dairy Industry" Viruses 14, no. 4: 810. https://0-doi-org.brum.beds.ac.uk/10.3390/v14040810