
A Novel Sphingosine Kinase Inhibitor Suppresses Chikungunya Virus Infection

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Compounds, and Virus
2.2. Sphingosine Kinase Inhibitor Studies
2.3. Immunofluorescence
2.4. Quantitative RT-PCR
2.5. Plaque Assay
2.6. Statistics
3. Results
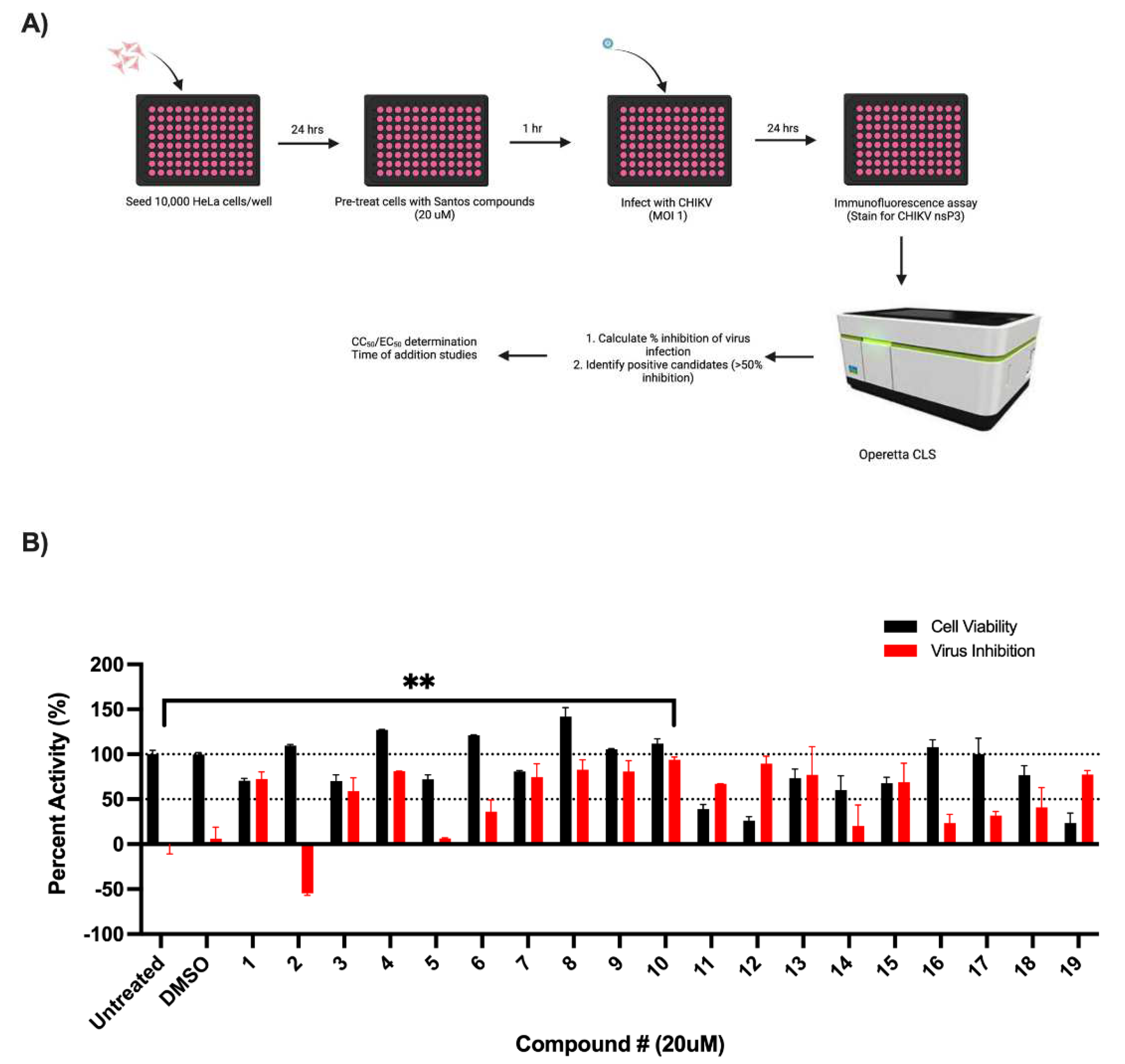
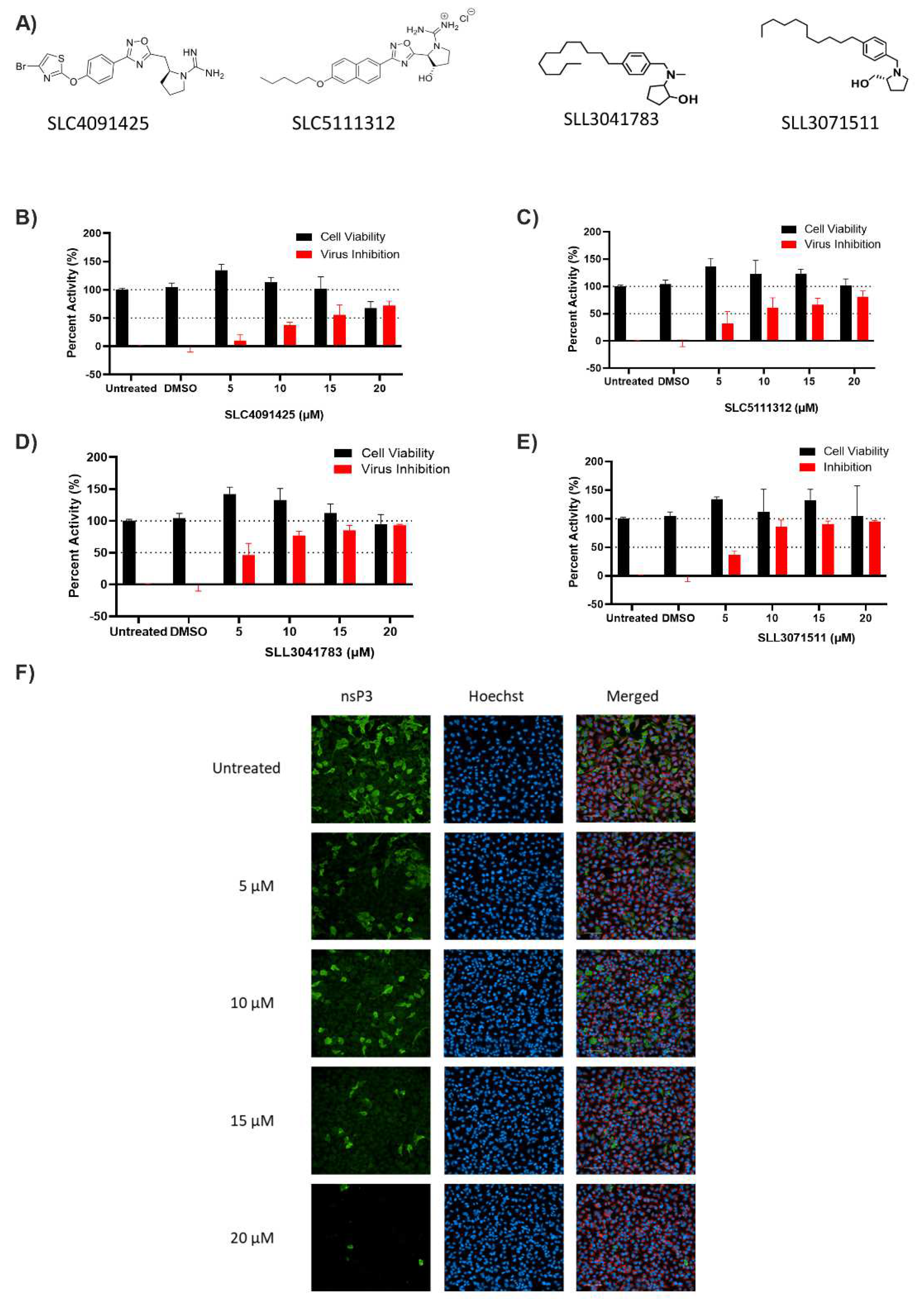
3.1. Screening of SK Inhibitors
3.2. Dose–Response and Time-Course of SLL3071511 Treatment
3.3. SLL3071511 Effect on CHIKV Gene Expression and Release
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, A.Y.; Martins, K.A.O.; Encinales, L.; Reid, S.P.; Acuna, M.; Encinales, C.; Matranga, C.B.; Pacheco, N.; Cure, C.; Shukla, B.; et al. Chikungunya Arthritis Mechanisms in the Americas: A Cross-Sectional Analysis of Chikungunya Arthritis Patients Twenty-Two Months After Infection Demonstrating No Detectable Viral Persistence in Synovial Fluid. Arthritis Rheumatol. 2018, 70, 585–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.M.O.; Kikuti, M.; Anjos, R.O.; Portilho, M.M.; Santos, V.C.; Goncalves, T.S.F.; Tauro, L.B.; Moreira, P.S.S.; Jacob-Nascimento, L.C.; Santana, P.M.; et al. Risk of chronic arthralgia and impact of pain on daily activities in a cohort of patients with chikungunya virus infection from Brazil. Int. J. Infect. Dis. 2021, 105, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Sissoko, D.; Malvy, D.; Ezzedine, K.; Renault, P.; Moscetti, F.; Ledrans, M.; Pierre, V. Post-epidemic Chikungunya disease on Reunion Island: Course of rheumatic manifestations and associated factors over a 15-month period. PLoS Negl. Trop. Dis. 2009, 3, e389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilte, C.; Staikowsky, F.; Couderc, T.; Madec, Y.; Carpentier, F.; Kassab, S.; Albert, M.L.; Lecuit, M.; Michault, A. Chikungunya virus-associated long-term arthralgia: A 36-month prospective longitudinal study. PLoS Negl. Trop. Dis. 2013, 7, e2137. [Google Scholar] [CrossRef]
- Ross, R.W. The Newala epidemic. III. The virus: Isolation, pathogenic properties and relationship to the epidemic. J. Hyg. 1956, 54, 177–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrel, R.N.; de Lamballerie, X.; Raoult, D. Chikungunya outbreaks—The globalization of vectorborne diseases. N. Engl. J. Med. 2007, 356, 769–771. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Lecuit, M. Chikungunya virus and the global spread of a mosquito-borne disease. N. Engl. J. Med. 2015, 372, 1231–1239. [Google Scholar] [CrossRef] [Green Version]
- Zeller, H.; Van Bortel, W.; Sudre, B. Chikungunya: Its History in Africa and Asia and Its Spread to New Regions in 2013–2014. J. Infect Dis. 2016, 214, S436–S440. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef]
- Zhang, H.; Desai, N.N.; Olivera, A.; Seki, T.; Brooker, G.; Spiegel, S. Sphingosine-1-phosphate, a novel lipid, involved in cellular proliferation. J. Cell Biol. 1991, 114, 155–167. [Google Scholar] [CrossRef]
- Olivera, A.; Spiegel, S. Sphingosine-1-phosphate as second messenger in cell proliferation induced by PDGF and FCS mitogens. Nature 1993, 365, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Olivera, A.; Spiegel, S. Sphingosine kinase: A mediator of vital cellular functions. Prostaglandins Other Lipid Mediat. 2001, 64, 123–134. [Google Scholar] [CrossRef]
- Cuvillier, O.; Pirianov, G.; Kleuser, B.; Vanek, P.G.; Coso, O.A.; Gutkind, S.; Spiegel, S. Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nature 1996, 381, 800–803. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Wang, L.; Gamble, J.R.; Vadas, M.A. Activation of sphingosine kinase by tumor necrosis factor-alpha inhibits apoptosis in human endothelial cells. J. Biol. Chem. 1999, 274, 34499–34505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivera, A.; Kohama, T.; Edsall, L.; Nava, V.; Cuvillier, O.; Poulton, S.; Spiegel, S. Sphingosine kinase expression increases intracellular sphingosine-1-phosphate and promotes cell growth and survival. J. Cell Biol. 1999, 147, 545–558. [Google Scholar] [CrossRef] [Green Version]
- Wattenberg, B.W. Role of sphingosine kinase localization in sphingolipid signaling. World J. Biol. Chem. 2010, 1, 362–368. [Google Scholar] [CrossRef]
- Siow, D.L.; Anderson, C.D.; Berdyshev, E.V.; Skobeleva, A.; Natarajan, V.; Pitson, S.M.; Wattenberg, B.W. Sphingosine kinase localization in the control of sphingolipid metabolism. Adv. Enzym. Regul. 2011, 51, 229–244. [Google Scholar] [CrossRef] [Green Version]
- Van Brocklyn, J.R.; Lee, M.-J.; Menzeleev, R.; Olivera, A.; Edsall, L.; Cuvillier, O.; Thomas, D.M.; Coopman, P.J.P.; Thangada, S.; Liu, C.H.; et al. Dual Actions of Sphingosine-1-Phosphate: Extracellular through the Gi-coupled Receptor Edg-1 and Intracellular to Regulate Proliferation and Survival. J. Cell Biol. 1998, 142, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Van Brocklyn, J.R.; Thangada, S.; Liu, C.H.; Hand, A.R.; Menzeleev, R.; Spiegel, S.; Hla, T. Sphingosine-1-phosphate as a ligand for the G protein coupled receptor EDG-1. Science 1998, 279, 1552–1555. [Google Scholar] [CrossRef]
- Hait, N.C.; Bellamy, A.; Milstien, S.; Kordula, T.; Spiegel, S. Sphingosine kinase type 2 activation by ERK-mediated phosphorylation. J. Biol. Chem. 2007, 282, 12058–12065. [Google Scholar] [CrossRef] [Green Version]
- Ding, G.; Sonoda, H.; Yu, H.; Kajimoto, T.; Goparaju, S.K.; Jahangeer, S.; Okada, T.; Nakamura, S. Protein kinase D-mediated phosphorylation and nuclear export of sphingosine kinase 2. J. Biol. Chem. 2007, 282, 27493–27502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igarashi, N.; Okada, T.; Hayashi, S.; Fujita, T.; Jahangeer, S.; Nakamura, S. Sphingosine kinase 2 is a nuclear protein and inhibits DNA synthesis. J. Biol. Chem. 2003, 278, 46832–46839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hait, N.C.; Allegood, J.; Maceyka, M.; Strub, G.M.; Harikumar, K.B.; Singh, S.K.; Luo, C.; Marmorstein, R.; Kordula, T.; Milstien, S.; et al. Regulation of histone acetylation in the nucleus by sphingosine-1-phosphate. Science 2009, 325, 1254–1257. [Google Scholar] [CrossRef] [Green Version]
- Strub, G.M.; Paillard, M.; Liang, J.; Gomez, L.; Allegood, J.C.; Hait, N.C.; Maceyka, M.; Price, M.M.; Chen, Q.; Simpson, D.C.; et al. Sphingosine-1-phosphate produced by sphingosine kinase 2 in mitochondria interacts with prohibitin 2 to regulate complex IV assembly and respiration. FASEB J. 2011, 25, 600–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nat. Rev. Cancer 2018, 18, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Jin, H.K.; Bae, J.S. Sphingolipids in neuroinflammation: A potential target for diagnosis and therapy. BMB Rep. 2019, 53, 28–34. [Google Scholar] [CrossRef]
- Ng, M.L.; Wadham, C.; Sukocheva, O.A. The role of sphingolipid signalling in diabetes associated pathologies (Review). Int J Mol. Med. 2017, 39, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Maia, L.P.; Santos, P.S.; Alves, P.T.; Rodrigues, C.M.; Araujo, T.G.; Maia, Y.C.P.; Camara, A.T.F.; Santos, D.W.; Goulart, L.R. Altered Leukocyte Sphingolipid Pathway in Breast Cancer. Int. J. Mol. Sci. 2017, 18, 2521. [Google Scholar] [CrossRef] [Green Version]
- Bryan, P.F.; Karla, C.; Edgar Alejandro, M.T.; Sara Elva, E.P.; Gemma, F.; Luz, C. Sphingolipids as Mediators in the Crosstalk between Microbiota and Intestinal Cells: Implications for Inflammatory Bowel Disease. Mediat. Inflamm. 2016, 2016, 9890141. [Google Scholar] [CrossRef] [Green Version]
- Carr, J.M.; Kua, T.; Clarke, J.N.; Calvert, J.K.; Zebol, J.R.; Beard, M.R.; Pitson, S.M. Reduced sphingosine kinase 1 activity in dengue virus type-2 infected cells can be mediated by the 3′ untranslated region of dengue virus type-2 RNA. J. Gen. Virol. 2013, 94, 2437–2448. [Google Scholar] [CrossRef]
- Seo, Y.J.; Pritzl, C.J.; Vijayan, M.; Bomb, K.; McClain, M.E.; Alexander, S.; Hahm, B. Sphingosine kinase 1 serves as a pro-viral factor by regulating viral RNA synthesis and nuclear export of viral ribonucleoprotein complex upon influenza virus infection. PLoS ONE 2013, 8, e75005. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.J.; Blake, C.; Alexander, S.; Hahm, B. Sphingosine 1-phosphate-metabolizing enzymes control influenza virus propagation and viral cytopathogenicity. J. Virol. 2010, 84, 8124–8131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayan, M.; Seo, Y.J.; Pritzl, C.J.; Squires, S.A.; Alexander, S.; Hahm, B. Sphingosine kinase 1 regulates measles virus replication. Virology 2014, 450–451, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.; Plaisance-Bonstaff, K.; Voelkel-Johnson, C.; Smith, C.D.; Ogretmen, B.; Qin, Z.; Parsons, C. Sphingosine kinase-2 maintains viral latency and survival for KSHV-infected endothelial cells. PLoS ONE 2014, 9, e102314. [Google Scholar] [CrossRef] [PubMed]
- Yamane, D.; Zahoor, M.A.; Mohamed, Y.M.; Azab, W.; Kato, K.; Tohya, Y.; Akashi, H. Inhibition of sphingosine kinase by bovine viral diarrhea virus NS3 is crucial for efficient viral replication and cytopathogenesis. J. Biol. Chem. 2009, 284, 13648–13659. [Google Scholar] [CrossRef] [Green Version]
- Reid, S.P.; Tritsch, S.R.; Kota, K.; Chiang, C.Y.; Dong, L.; Kenny, T.; Brueggemann, E.E.; Ward, M.D.; Cazares, L.H.; Bavari, S. Sphingosine kinase 2 is a chikungunya virus host factor co-localized with the viral replication complex. Emerg. Microbes Infect. 2015, 4, e61. [Google Scholar] [CrossRef]
- Gao, P.; Peterson, Y.K.; Smith, R.A.; Smith, C.D. Characterization of isoenzyme-selective inhibitors of human sphingosine kinases. PLoS ONE 2012, 7, e44543. [Google Scholar] [CrossRef]
- French, K.J.; Zhuang, Y.; Maines, L.W.; Gao, P.; Wang, W.; Beljanski, V.; Upson, J.J.; Green, C.L.; Keller, S.N.; Smith, C.D. Pharmacology and antitumor activity of ABC294640, a selective inhibitor of sphingosine kinase-2. J. Pharmacol. Exp. Ther. 2010, 333, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Congdon, M.D.; Kharel, Y.; Brown, A.M.; Lewis, S.N.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Structure-Activity Relationship Studies and Molecular Modeling of Naphthalene-Based Sphingosine Kinase 2 Inhibitors. ACS Med. Chem. Lett. 2016, 7, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Childress, E.S.; Kharel, Y.; Brown, A.M.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Transforming Sphingosine Kinase 1 Inhibitors into Dual and Sphingosine Kinase 2 Selective Inhibitors: Design, Synthesis, and in Vivo Activity. J. Med. Chem. 2017, 60, 3933–3957. [Google Scholar] [CrossRef]
- Vettorazzi, M.; Insuasty, D.; Lima, S.; Gutierrez, L.; Nogueras, M.; Marchal, A.; Abonia, R.; Andujar, S.; Spiegel, S.; Cobo, J.; et al. Design of new quinolin-2-one-pyrimidine hybrids as sphingosine kinases inhibitors. Bioorg. Chem. 2019, 94, 103414. [Google Scholar] [CrossRef] [PubMed]
- Sibley, C.D.; Morris, E.A.; Kharel, Y.; Brown, A.M.; Huang, T.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Discovery of a Small Side Cavity in Sphingosine Kinase 2 that Enhances Inhibitor Potency and Selectivity. J. Med. Chem. 2020, 63, 1178–1198. [Google Scholar] [CrossRef] [PubMed]
- Congdon, M.; Fritzemeier, R.G.; Kharel, Y.; Brown, A.M.; Serbulea, V.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Probing the substitution pattern of indole-based scaffold reveals potent and selective sphingosine kinase 2 inhibitors. Eur. J. Med. Chem. 2021, 212, 113121. [Google Scholar] [CrossRef]
- Kharel, Y.; Morris, E.A.; Congdon, M.D.; Thorpe, S.B.; Tomsig, J.L.; Santos, W.L.; Lynch, K.R. Sphingosine Kinase 2 Inhibition and Blood Sphingosine 1-Phosphate Levels. J. Pharmacol. Exp. Ther. 2015, 355, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Sibley, C.D.; Kharel, Y.; Huang, T.; Brown, A.M.; Wonilowicz, L.G.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Lipophilic tail modifications of 2-(hydroxymethyl)pyrrolidine scaffold reveal dual sphingosine kinase 1 and 2 inhibitors. Bioorg. Med. Chem. 2021, 30, 115941. [Google Scholar] [CrossRef] [PubMed]
- Kharel, Y.; Agah, S.; Huang, T.; Mendelson, A.J.; Eletu, O.T.; Barkey-Bircann, P.; Gesualdi, J.; Smith, J.S.; Santos, W.L.; Lynch, K.R. Saccharomyces cerevisiae as a platform for assessing sphingolipid lipid kinase inhibitors. PLoS ONE 2018, 13, e0192179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.L.; Coates, E.E.; Plummer, S.H.; Carter, C.A.; Berkowitz, N.; Conan-Cibotti, M.; Cox, J.H.; Beck, A.; O′Callahan, M.; Andrews, C.; et al. Effect of a Chikungunya Virus-Like Particle Vaccine on Safety and Tolerability Outcomes: A Randomized Clinical Trial. JAMA 2020, 323, 1369–1377. [Google Scholar] [CrossRef]
- Wolf, J.J.; Studstill, C.J.; Hahm, B. Emerging Connections of S1P-Metabolizing Enzymes with Host Defense and Immunity During Virus Infections. Viruses 2019, 11, 1097. [Google Scholar] [CrossRef] [Green Version]
- Morchang, A.; Lee, R.C.H.; Yenchitsomanus, P.T.; Sreekanth, G.P.; Noisakran, S.; Chu, J.J.H.; Limjindaporn, T. RNAi screen reveals a role of SPHK2 in dengue virus-mediated apoptosis in hepatic cell lines. PLoS ONE 2017, 12, e0188121. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Seo, Y.J.; Studstill, C.J.; Vijayan, M.; Wolf, J.J.; Hahm, B. Transient inhibition of sphingosine kinases confers protection to influenza A virus infected mice. Antivir. Res. 2018, 158, 171–177. [Google Scholar] [CrossRef]
- Monick, M.M.; Cameron, K.; Powers, L.S.; Butler, N.S.; McCoy, D.; Mallampalli, R.K.; Hunninghake, G.W. Sphingosine kinase mediates activation of extracellular signal-related kinase and Akt by respiratory syncytial virus. Am. J. Respir. Cell Mol. Biol. 2004, 30, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.P.; Xiao, Z.L.; Yang, Z.; Li, J.; Feng, G.X.; Chen, F.Q.; Li, Y.H.; Feng, J.Y.; Gao, Y.E.; Ye, L.H.; et al. Hepatitis B virus X protein promotes human hepatoma cell growth via upregulation of transcription factor AP2alpha and sphingosine kinase 1. Acta Pharmacol. Sin. 2015, 36, 1228–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitson, S.M. Regulation of sphingosine kinase and sphingolipid signaling. Trends Biochem. Sci. 2011, 36, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Bektas, M.; Jolly, P.S.; Muller, C.; Eberle, J.; Spiegel, S.; Geilen, C.C. Sphingosine kinase activity counteracts ceramide-mediated cell death in human melanoma cells: Role of Bcl-2 expression. Oncogene 2005, 24, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Evangelisti, C.; Evangelisti, C.; Buontempo, F.; Lonetti, A.; Orsini, E.; Chiarini, F.; Barata, J.T.; Pyne, S.; Pyne, N.J.; Martelli, A.M. Therapeutic potential of targeting sphingosine kinases and sphingosine 1-phosphate in hematological malignancies. Leukemia 2016, 30, 2142–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, P.; Gamble, J.R.; Wang, L.; Pitson, S.M.; Moretti, P.A.; Wattenberg, B.W.; D′Andrea, R.J.; Vadas, M.A. An oncogenic role of sphingosine kinase. Curr. Biol. 2000, 10, 1527–1530. [Google Scholar] [CrossRef]
- Rex, K.; Jeffries, S.; Brown, M.L.; Carlson, T.; Coxon, A.; Fajardo, F.; Frank, B.; Gustin, D.; Kamb, A.; Kassner, P.D.; et al. Sphingosine kinase activity is not required for tumor cell viability. PLoS ONE 2013, 8, e68328. [Google Scholar] [CrossRef] [PubMed]
- Kharel, Y.; Raje, M.; Gao, M.; Gellett, A.M.; Tomsig, J.L.; Lynch, K.R.; Santos, W.L. Sphingosine kinase type 2 inhibition elevates circulating sphingosine 1-phosphate. Biochem. J. 2012, 447, 149–157. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Compound ID | SK1 Ki (µM) | SK2 Ki (µM) | EC50 (µM) |
|---|---|---|---|
| SLC4091425 | > 3 | >3 | 3.35 |
| SLC5111312 | 0.36 ± 0.09 | 0.33 ± 0.07 | 11.84 |
| SLL3041783 | 2.4 ± 0.61 | >3 | 3.33 |
| SLL3071511 | 0.27 ± 0.03 | 0.66 ± 0.11 | 2.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyewole, O.O.; Dunnavant, K.; Bhattarai, S.; Kharel, Y.; Lynch, K.R.; Santos, W.L.; Reid, S.P. A Novel Sphingosine Kinase Inhibitor Suppresses Chikungunya Virus Infection. Viruses 2022, 14, 1123. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061123
Oyewole OO, Dunnavant K, Bhattarai S, Kharel Y, Lynch KR, Santos WL, Reid SP. A Novel Sphingosine Kinase Inhibitor Suppresses Chikungunya Virus Infection. Viruses. 2022; 14(6):1123. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061123
Chicago/Turabian StyleOyewole, Opeoluwa O., Kyle Dunnavant, Shaurav Bhattarai, Yugesh Kharel, Kevin R. Lynch, Webster L. Santos, and St. Patrick Reid. 2022. "A Novel Sphingosine Kinase Inhibitor Suppresses Chikungunya Virus Infection" Viruses 14, no. 6: 1123. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061123