
Isolation of Genetically Diverse H5N8 Avian Influenza Viruses in Poultry in Egypt, 2019–2021


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Detection and Isolation
2.2. Sequence of the HA and NA Genes
2.3. Phylogenetic Analysis
3. Results
3.1. Surveillance
3.2. HA Phylogenetic Analysis
3.3. NA Phylogenetic Analysis
3.4. Comparison of HA Protein with Commercially Licensed H5 Vaccines in Poultry in Egypt
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaplan, B.S.; Webby, R.J. The avian and mammalian host range of highly pathogenic avian H5N1 influenza. Virus Res. 2013, 178, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.J. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.J.; Donis, R.O. Nomenclature updates resulting from the evolution of avian influenza A(H5) virus clades 2.1.3.2a, 2.2.1, and 2.3.4 during 2013–2014. Influenza Respir. Viruses 2015, 9, 271–276. [Google Scholar]
- Antigua, K.J.C.; Choi, W.S.; Baek, Y.H.; Song, M.S. The Emergence and Decennary Distribution of Clade 2.3.4.4 HPAI H5Nx. Microorganisms 2019, 7, 156. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Bertran, K.; Kwon, J.H.; Swayne, D.E. Evolution, global spread, and pathogenicity of highly pathogenic avian influenza H5Nx clade 2.3.4.4. J. Vet. Sci. 2017, 18, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Floyd, T.; Banyard, A.C.; Lean, F.Z.X.; Byrne, A.M.P.; Fullick, E.; Whittard, E.; Mollett, B.C.; Bexton, S.; Swinson, V.; Macrelli, M.; et al. Encephalitis and Death in Wild Mammals at a Rehabilitation Center after Infection with Highly Pathogenic Avian Influenza A(H5N8) Virus, United Kingdom. Emerg. Infect. Dis. 2021, 27, 2856–2863. [Google Scholar] [CrossRef] [PubMed]
- Pyankova, O.G.; Susloparov, I.M.; Moiseeva, A.A.; Kolosova, N.P.; Onkhonova, G.S.; Danilenko, A.V.; Vakalova, E.V.; Shendo, G.L.; Nekeshina, N.N.; Noskova, L.N.; et al. Isolation of clade 2.3.4.4b A(H5N8), a highly pathogenic avian influenza virus, from a worker during an outbreak on a poultry farm, Russia, December 2020. Eurosurveillance 2021, 26, 2100439. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.L.; Siebert, U.; Lakemeyer, J.; Grilo, M.; Pawliczka, I.; Wu, N.H.; Valentin-Weigand, P.; Haas, L.; Herrler, G. Highly Pathogenic Avian Influenza A(H5N8) Virus in Gray Seals, Baltic Sea. Emerg. Infect. Dis. 2019, 25, 2295–2298. [Google Scholar] [CrossRef] [Green Version]
- Oliver, I.; Roberts, J.; Brown, C.S.; Byrne, A.M.; Mellon, D.; Hansen, R.; Banyard, A.C.; James, J.; Donati, M.; Porter, R.; et al. A case of avian influenza A(H5N1) in England, January 2022. Eurosurveillance 2022, 27, 2200061. [Google Scholar] [CrossRef]
- Zheng, Z.; Lu, Y.; Short, K.R.; Lu, J. One health insights to prevent the next HxNy viral outbreak: Learning from the epidemiology of H7N9. BMC Infect. Dis. 2019, 19, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H. H5N1 avian influenza in China. Sci. China C Life Sci. 2009, 52, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Bu, Z.; Chen, H. Avian influenza vaccines against H5N1 ‘bird flu’. Trends Biotechnol. 2014, 32, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Jadhao, S.J.; Lee, C.W.; Sylte, M.; Suarez, D.L. Comparative efficacy of North American and antigenically matched reverse genetics derived H5N9 DIVA marker vaccines against highly pathogenic Asian H5N1 avian influenza viruses in chickens. Vaccine 2009, 27, 6247–6260. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Deng, G.; Ma, S.; Zeng, X.; Yin, X.; Li, M.; Zhang, B.; Cui, P.; Chen, Y.; Yang, H.; et al. Rapid Evolution of H7N9 Highly Pathogenic Viruses that Emerged in China in 2017. Cell Host Microbe 2018, 24, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Swayne, D.E.; Kapczynski, D.R. Vaccines and vaccination for avian influenza in poultry. In Animal Influenza; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 378–434. [Google Scholar]
- Zeng, X.; Tian, G.; Shi, J.; Deng, G.; Li, C.; Chen, H. Vaccination of poultry successfully eliminated human infection with H7N9 virus in China. Sci. China Life Sci. 2018, 61, 1465–1473. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Chen, X.; Ma, S.; Wu, J.; Bao, H.; Pan, S.; Liu, Y.; Deng, G.; Shi, J.; Chen, P.; et al. Protective efficacy of an H5/H7 trivalent inactivated vaccine produced from Re-11, Re-12, and H7-Re2 strains against challenge with different H5 and H7 viruses in chickens. J. Integr. Agric. 2020, 19, 2294–2300. [Google Scholar] [CrossRef]
- Hafez, M.H.; Arafa, A.; Abdelwhab, E.M.; Selim, A.; Khoulosy, S.G.; Hassan, M.K.; Aly, M.M. Avian influenza H5N1 virus infections in vaccinated commercial and backyard poultry in Egypt. Poult. Sci. 2010, 89, 1609–1613. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-W.; Senne, D.A.; Suarez, D.L. Effect of vaccine use in the evolution of Mexican lineage H5N2 avian influenza virus. J. Virol. 2004, 78, 8372–8381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H. Challenge for One Health: Co-Circulation of Zoonotic H5N1 and H9N2 Avian Influenza Viruses in Egypt. Viruses 2018, 10, 121. [Google Scholar] [CrossRef] [Green Version]
- Hassan, K.E.; El-Kady, M.F.; EL-Sawah, A.A.A.; Luttermann, C.; Parvin, R.; Shany, S.; Beer, M.; Harder, T. Respiratory disease due to mixed viral infections in poultry flocks in Egypt between 2017 and 2018: Upsurge of highly pathogenic avian influenza virus subtype H5N8 since 2018. Transbound. Emerg. Dis. 2021, 68, 21–36. [Google Scholar] [CrossRef]
- Amer, F.; Li, R.; Rabie, N.; El-Husseiny, M.H.; Yehia, N.; Hagag, N.M.; Samy, M.; Selim, A.; Hassan, M.K.; Hassan, W.M.M.; et al. Temporal Dynamics of Influenza A(H5N1) Subtype before and after the Emergence of H5N8. Viruses 2021, 13, 1565. [Google Scholar] [CrossRef] [PubMed]
- Salaheldin, A.H.; El-Hamid, A.; Elbestawy, A.R.; Veits, J.; Hafez, H.M.; Mettenleiter, T.C.; Abdelwhab, E.M. Multiple Introductions of Influenza A(H5N8) Virus into Poultry, Egypt, 2017. Emerg. Infect. Dis. 2018, 24, 943–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yehia, N.; Naguib, M.M.; Li, R.; Hagag, N.; El-Husseiny, M.; Mosaad, Z.; Nour, A.; Rabea, N.; Hasan, W.M.; Hassan, M.K.; et al. Multiple introductions of reassorted highly pathogenic avian influenza viruses (H5N8) clade 2.3.4.4b causing outbreaks in wild birds and poultry in Egypt. Infect. Genet. Evol. 2018, 58, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.E.; Saad, N.; Abozeid, H.H.; Shany, S.; El-Kady, M.F.; Arafa, A.; El-Sawah, A.A.A.; Pfaff, F.; Hafez, H.M.; Beer, M.; et al. Genotyping and reassortment analysis of highly pathogenic avian influenza viruses H5N8 and H5N2 from Egypt reveals successive annual replacement of genotypes. Infect. Genet. Evol. 2020, 84, 104375. [Google Scholar] [CrossRef] [PubMed]
- Kammon, A.; Heidari, A.; Dayhum, A.; Eldaghayes, I.; Sharif, M.; Monne, I.; Cattoli, G.; Asheg, A.; Farhat, M.; Kraim, E. Characterization of Avian Influenza and Newcastle Disease Viruses from Poultry in Libya. Avian Dis. 2015, 59, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Salaheldin, A.H.; Veits, J.; Abd El-Hamid, H.S.; Harder, T.C.; Devrishov, D.; Mettenleiter, T.C.; Hafez, H.M.; Abdelwhab, E.M. Isolation and genetic characterization of a novel 2.2.1.2a H5N1 virus from a vaccinated meat-turkeys flock in Egypt. Virol. J. 2017, 14, 48. [Google Scholar] [CrossRef] [Green Version]
- WHO. Cumulative Number of Confirmed Human Cases for Avian Influenza A(H5N1) Reported to WHO, 2003–2021, 15 April 2021. Available online: https://www.who.int/publications/m/item/cumulative-number-of-confirmed-human-cases-for-avian-influenza-a(h5n1)-reported-to-who-2003-2021-15-april-2021 (accessed on 16 March 2022).
- Gomaa, M.R.; Kayed, A.S.; Elabd, M.A.; Zeid, D.A.; Zaki, S.A.; El Rifay, A.S.; Sherif, L.S.; McKenzie, P.P.; Webster, R.G.; Webby, R.J.; et al. Avian influenza A(H5N1) and A(H9N2) seroprevalence and risk factors for infection among Egyptians: A prospective, controlled seroepidemiological study. J. Infect. Dis. 2015, 211, 1399–1407. [Google Scholar] [CrossRef] [Green Version]
- Abdelwhab, E.M.; Hafez, H.M. An overview of the epidemic of highly pathogenic H5N1 avian influenza virus in Egypt: Epidemiology and control challenges. Epidemiol. Infect. 2011, 139, 647–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelwhab, E.M.; Hassan, M.K.; Abdel-Moneim, A.S.; Naguib, M.M.; Mostafa, A.; Hussein, I.T.M.; Arafa, A.; Erfan, A.M.; Kilany, W.H.; Agour, M.G.; et al. Introduction and enzootic of A/H5N1 in Egypt: Virus evolution, pathogenicity and vaccine efficacy ten years on. Infect. Genet. Evol 2016, 40, 80–90. [Google Scholar] [CrossRef]
- Cattoli, G.; Milani, A.; Temperton, N.; Zecchin, B.; Buratin, A.; Molesti, E.; Aly, M.M.; Arafa, A.; Capua, I. Antigenic drift in H5N1 avian influenza virus in poultry is driven by mutations in major antigenic sites of the hemagglutinin molecule analogous to those for human influenza virus. J. Virol. 2011, 85, 8718–8724. [Google Scholar] [CrossRef] [Green Version]
- Cattoli, G.; Fusaro, A.; Monne, I.; Coven, F.; Joannis, T.; El-Hamid, H.S.; Hussein, A.A.; Cornelius, C.; Amarin, N.M.; Mancin, M.; et al. Evidence for differing evolutionary dynamics of A/H5N1 viruses among countries applying or not applying avian influenza vaccination in poultry. Vaccine 2011, 29, 9368–9375. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.; Hoffmann, D.; Henritzi, D.; Beer, M.; Harder, T.C. Riems influenza a typing array (RITA): An RT-qPCR-based low density array for subtyping avian and mammalian influenza a viruses. Sci. Rep. 2016, 6, 27211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D.R. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef]
- Tarek, M.; Naguib, M.M.; Arafa, A.S.; Tantawy, L.A.; Selim, K.M.; Talaat, S.; Sultan, H.A. Epidemiology, Genetic Characterization, and Pathogenesis of Avian Influenza H5N8 Viruses Circulating in Northern and Southern Parts of Egypt, 2017–2019. Animals 2021, 11, 2208. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Milne, I.; Lindner, D.; Bayer, M.; Husmeier, D.; McGuire, G.; Marshall, D.F.; Wright, F. TOPALi v2: A rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 2009, 25, 126–127. [Google Scholar] [CrossRef] [Green Version]
- Duvvuri, V.R.; Duvvuri, B.; Cuff, W.R.; Wu, G.E.; Wu, J. Role of positive selection pressure on the evolution of H5N1 hemagglutinin. Genom. Proteom. Bioinform. 2009, 7, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Cui, P.; Zeng, X.; Li, X.; Li, Y.; Shi, J.; Zhao, C.; Qu, Z.; Wang, Y.; Guo, J.; Gu, W.; et al. Genetic and biological characteristics of the globally circulating H5N8 avian influenza viruses and the protective efficacy offered by the poultry vaccine currently used in China. Sci. China Life Sci. 2022, 65, 795–808. [Google Scholar] [CrossRef]
- Kandeil, A.; Sabir, J.S.M.; Abdelaal, A.; Mattar, E.H.; El-Taweel, A.N.; Sabir, M.J.; Khalil, A.A.; Webby, R.; Kayali, G.; Ali, M.A. Efficacy of commercial vaccines against newly emerging avian influenza H5N8 virus in Egypt. Sci. Rep. 2018, 8, 9697. [Google Scholar] [CrossRef]
- Swayne, D.E.; Spackman, E.; Pantin-Jackwood, M. Success factors for avian influenza vaccine use in poultry and potential impact at the wild bird-agricultural interface. Ecohealth 2014, 11, 94–108. [Google Scholar] [CrossRef]
- Yehia, N.; Hassan, W.M.M.; Sedeek, A.; Elhusseiny, M.H. Genetic variability of avian influenza virus subtype H5N8 in Egypt in 2017 and 2018. Arch. Virol. 2020, 165, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Peyre, M.; Samaha, H.; Makonnen, Y.J.; Saad, A.; Abd-Elnabi, A.; Galal, S.; Ettel, T.; Dauphin, G.; Lubroth, J.; Roger, F.; et al. Avian influenza vaccination in Egypt: Limitations of the current strategy. J. Mol. Genet. Med. 2009, 3, 198–204. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
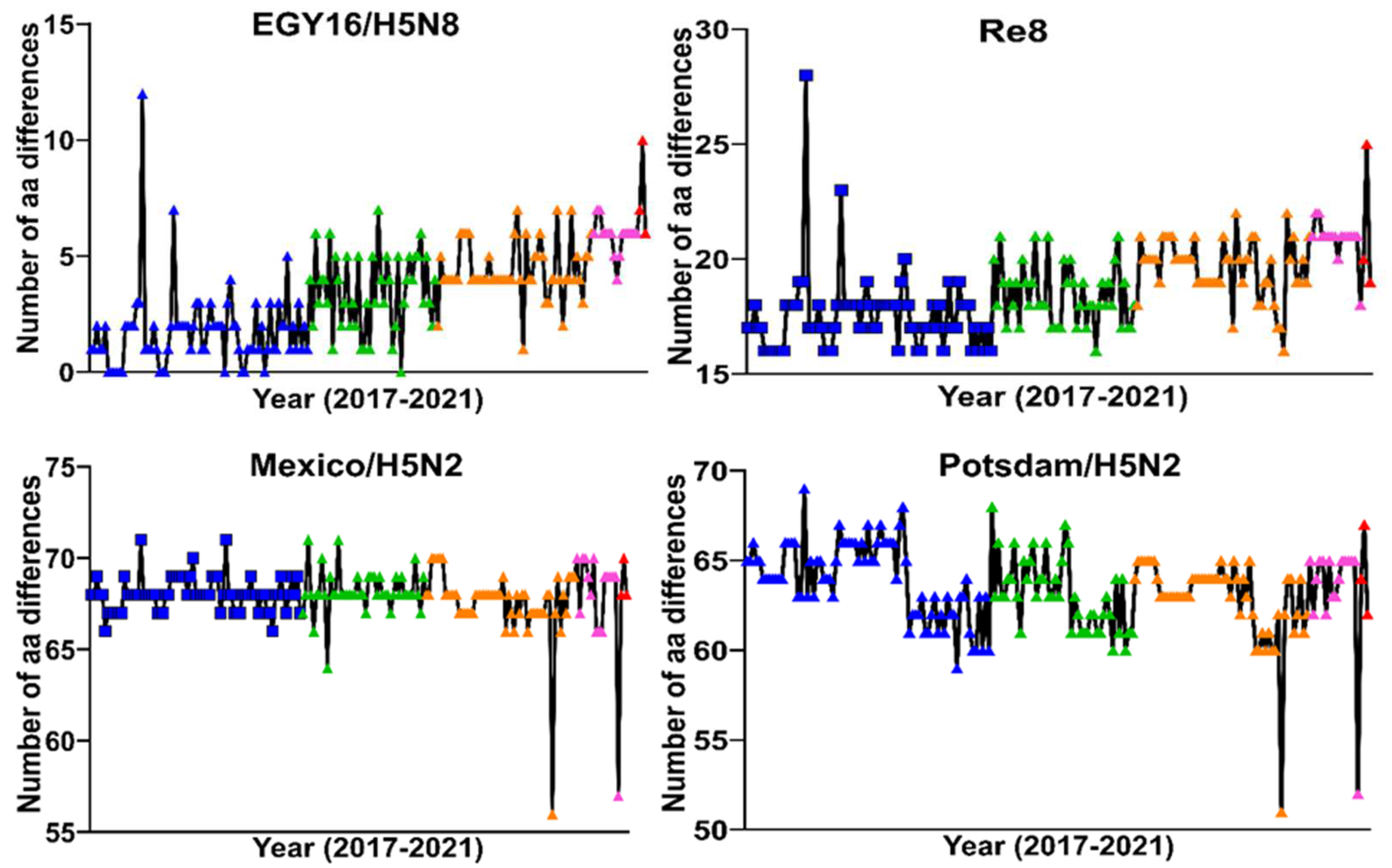{kind=link}
| Virus | Date | Locality | Breed | Age | Number of Animals | Frequency of Vaccination | Accession Numbers | |
|---|---|---|---|---|---|---|---|---|
| HA | NA | |||||||
| A/duck/Egypt/Behera-AH1/2019 | October 2019 | Elbehera | Broiler | 34d | 18,000 | Once | EPI1999284 | EPI1999283 |
| A/turkey/Egypt/Alex-AH/2019 | October 2019 | Alexandria | Turkey | 55d | 6000 | Once | EPI1999276 | EPI1999275 |
| A/chicken/Egypt/Assuit-AH/2019 | November 2019 | Assuit | Broiler | 28d | 10,000 | Not | EPI1999303 | EPI1999302 |
| A/chicken/Egypt/Kafrelsheik-AH/2019 | November 2019 | Kafrelsheik | Broiler | 29d | 20,000 | Once | EPI1999290 | EPI1999289 |
| A/chicken/Egypt/Elmonufia-backyard-AH/2019 | December 2019 | Elmonufia | Local Breed | 44d | 500 | Not | EPI1999296 | EPI1999295 |
| A/turkey/Egypt/Cairo/AH/2019 | December 2019 | Cairo | Turkey | 60d | 7000 | Once | EPI1999274 | EPI1999273 |
| A/turkey/Egypt/Alex-AH1/2019 | December 2019 | Alexandria | Turkey | 90d | 5000 | Once | EPI1999279 | EPI1999277 |
| A/duck/Egypt/Elbehera-AH2/2019 | December 2019 | Elbehera | Duck | 40d | 10,000 | Once | EPI1999281 | EPI1999280 |
| A/chicken/Egypt/Cairo-HC11B-AH/2020 | January 2020 | Cairo | Broiler | 33d | 21,000 | Once | EPI1999299 | n.d. |
| A/duck/Egypt/Behera-HB2-AH/2020 | January 2020 | Elbehera | Duck | 45d | 12,000 | Once | EPI1999282 | n.d. |
| A/chicken/Egypt/Giza-AH/2020 | January 2020 | Giza | Layer | 24 wks | 20,000 | 3 times | EPI1999294 | EPI1999293 |
| A/chicken/Egypt/Alex-AH/2020 | January 2020 | Alexandria | Broiler | 31d | 17,000 | Once | EPI1999308 | EPI1999306 |
| A/chicken/Egypt/Elmonufia-AH/2020 | February 2020 | Elmonufia | Broiler | 26d | 18,000 | Once | EPI1999298 | EPI1999297 |
| A/chicken/Egypt/Giza-HG4L-AH/2020 | February 2020 | Giza | Broiler | 28d | 16,000 | Once | EPI1999292 | n.d. |
| A/chicken/Egypt/Sohag-AH/2020 | February 2020 | Sohag | Broiler | 32d | 15,000 | Not | EPI1999286 | EPI1999285 |
| A/chicken/Egypt/Qalyubia-layer-AH/2020 | March 2020 | Qalyubia | Layer | 32 wks | 35,000 | 3 times | EPI1999288 | EPI1999287 |
| A/chicken/Egypt/Alex-AH2/2020 | March 2020 | Alexandria | Broiler | 19d | 22,000 | Once | EPI1999310 | EPI1999309 |
| A/chicken/Egypt/Alex-AH1/2020 | May 2020 | Alexandria | Broiler | 29d | 15,000 | Once | EPI1999312 | EPI1999311 |
| A/chicken/Egypt/Behera-AH/2021 | February 2021 | Elbehera | Broiler | 33 | 12,000 | Once | EPI1999301 | EPI1999300 |
| A/chicken/Egypt/Alex-Breeder-AH/2021 | March 2021 | Alexandria | Broiler Breeder | 40 wks | 16,000 | 4 times | EPI1999305 | EPI1999304 |
| No. | Vaccine Seed Virus | Subtype | Abbreviation | Clade/Lineage | Accession Numbers (aa) | Company | AA Identity to Egyptian H5N8 (Min–Max) |
|---|---|---|---|---|---|---|---|
| 1 | A/chicken/Mexico/232/1994 | H5N2 | Mexico/H5N2 | North American | AAR88841 | Ceva, Mexico | 75.3–90.1 |
| 2 | A/duck/Potsdam/1402-6/1986 | H5N2 | Potsdam/H5N2 | Eurasian | ABI84497 | Intervet, The Netherlands | 87.2–92.0 |
| 3 | A/chicken/Egypt/18-H/2009 | H5N1 | EGY09/H5N1 | 2.2.1.1 | ADG28676 | Harbin Veterinary Research Institute, China | 88.7–94.2 |
| 4 | A/duck/Egypt/M2583D /2010 | H5N1 | EGY10/H5N1 | 2.2.1.1 | AEP37317 | ME-VAC, Egypt | 90.8–96.4 |
| 5 | A/chicken/Vietnam/C58/2004 | H5N1 | Vietnam/H5N1 | 1 | AAW80718.1 | Zoetis, USA | 90.9–96.7 |
| 6 | A/duck/China/E319-2/2003 | H5N1 | CN03/H5N1 | 2.3.2 | AAR99628 | Boehringer Ingelheim, Germany | 92.5–98.4 |
| 7 | A/duck/Anhui/1/2006 | H5N1 | Re5 | 2.3.4 | ADG59091 | QYH, China | 92.9–98.4 |
| 8 | A/chicken/Guizhou/4/2013 | H5N1 | Re8 | 2.3.4.4 | EPI675769 | Merial, USA & QYH, China | 94.9–97.1 |
| 9 | A/green-winged teal/Egypt/877/2016 | H5N8 | EGY16/H5N8 | 2.3.4.4b | ART29489 | ME-VAC, Egypt | 97.8–100 |
| Phylogroup | Virus/aa Position (H5 Numbering) * | 72 | 94 | 140 | 169 | 188 | 236 | 268 | 522 | 532 |
|---|---|---|---|---|---|---|---|---|---|---|
| Vaccine strain | A/green-winged teal/Egypt/877/2016 | R | S | T | R | T | N | E | V | V |
| Egypt-II | A/Duck/Egypt/Elbehera_AH2/2019 | N | . | . | . | . | . | G | . | . |
| A/chicken/Egypt/kafrelsheik_AH/2019 | N | . | . | . | I | . | G | . | . | |
| A/chicken/Egypt/Alex_AH1/2020 | N | . | . | . | I | . | G | A | . | |
| A/chicken/Egypt/Alex_AH/2020 | N | . | . | . | I | . | G | A | M | |
| A/chicken/Egypt/Alex_AH2/2020 | N | . | . | . | I | . | G | . | . | |
| A/chicken/Egypt/Elmonufia_AH/2020 | S | R | . | Q | . | . | G | . | M | |
| A/turkey/Egypt/Cairo/AH/2019 | S | R | . | Q | . | . | G | . | . | |
| A/turkey/Egypt/Alex_AH/2019 | S | R | . | Q | . | . | G | . | . | |
| A/duck/Egypt/Behera_AH1/2019 | S | R | . | Q | . | . | G | . | . | |
| Egypt-III | A/chicken/Egypt/behera-AH/2021 | . | . | A | Q | . | . | G | A | . |
| A/chiken/Egypt/Alex-Breeder-AH/2021 | . | . | A | Q | . | . | G | A | M | |
| A/chicken/Egypt/Giza-AH/2020 | . | . | A | . | . | D | G | A | M | |
| A/chicken/Egypt/Elmonufia-backyard-AH/2019 | . | . | A | . | . | D | G | A | M | |
| A/chicken/Egypt/qalyubia-layer-AH/2020 | . | . | A | . | N | D | G | A | M | |
| A/chicken/Egypt/sohag-AH/2020 | . | . | A | . | . | D | G | A | M | |
| A/chicken/Egypt/Assuit-AH/2019 | . | . | A | . | . | D | G | A | M | |
| A/Duck/Egypt/Behera-HB2-AH/2020 | . | . | A | . | . | D | G | A | M | |
| A/Chicken/Egypt/Giza-HG4L-AH/2020 | . | . | . | . | . | D | G | A | M | |
| A/Chicken/Egypt/Cairo-HC11B-AH/20 | . | . | A | . | . | D | G | A | M | |
| A/turkey/Egypt/Alex-AH1/2019 | . | . | A | . | . | . | G | A | M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salaheldin, A.H.; Elbestawy, A.R.; Abdelkader, A.M.; Sultan, H.A.; Ibrahim, A.A.; Abd El-Hamid, H.S.; Abdelwhab, E.M. Isolation of Genetically Diverse H5N8 Avian Influenza Viruses in Poultry in Egypt, 2019–2021. Viruses 2022, 14, 1431. https://0-doi-org.brum.beds.ac.uk/10.3390/v14071431
Salaheldin AH, Elbestawy AR, Abdelkader AM, Sultan HA, Ibrahim AA, Abd El-Hamid HS, Abdelwhab EM. Isolation of Genetically Diverse H5N8 Avian Influenza Viruses in Poultry in Egypt, 2019–2021. Viruses. 2022; 14(7):1431. https://0-doi-org.brum.beds.ac.uk/10.3390/v14071431
Chicago/Turabian StyleSalaheldin, Ahmed H., Ahmed R. Elbestawy, Abdelkader M. Abdelkader, Hesham A. Sultan, Awad A. Ibrahim, Hatem S. Abd El-Hamid, and Elsayed M. Abdelwhab. 2022. "Isolation of Genetically Diverse H5N8 Avian Influenza Viruses in Poultry in Egypt, 2019–2021" Viruses 14, no. 7: 1431. https://0-doi-org.brum.beds.ac.uk/10.3390/v14071431