
Neutralizing mAbs against SFTS Virus Gn Protein Show Strong Therapeutic Effects in an SFTS Animal Model

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Control Antibodies
2.2. SFTS Virus-Infected Cell Lysate
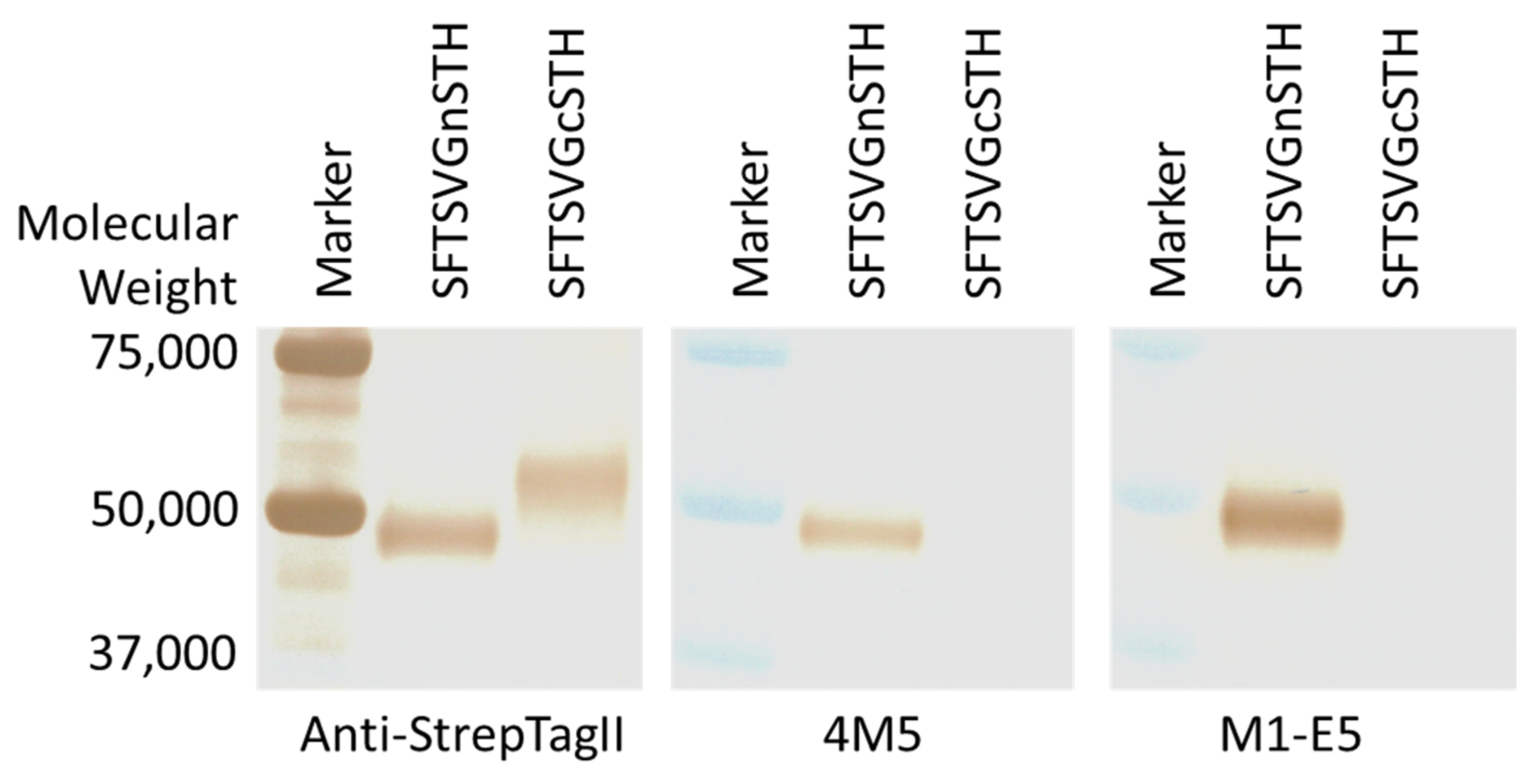
2.3. Recombinant Proteins
2.4. Human mAbs
2.5. Mouse mAbs
2.6. Neutralization Tests
2.7. Therapeutic Effects
2.8. Competition ELISA
2.9. ELISA with Fc-Tagged Gn
2.10. Ethical Statements
2.11. Statistics
3. Results
3.1. mAbs Specific to SFTS Virus Gn and Gc
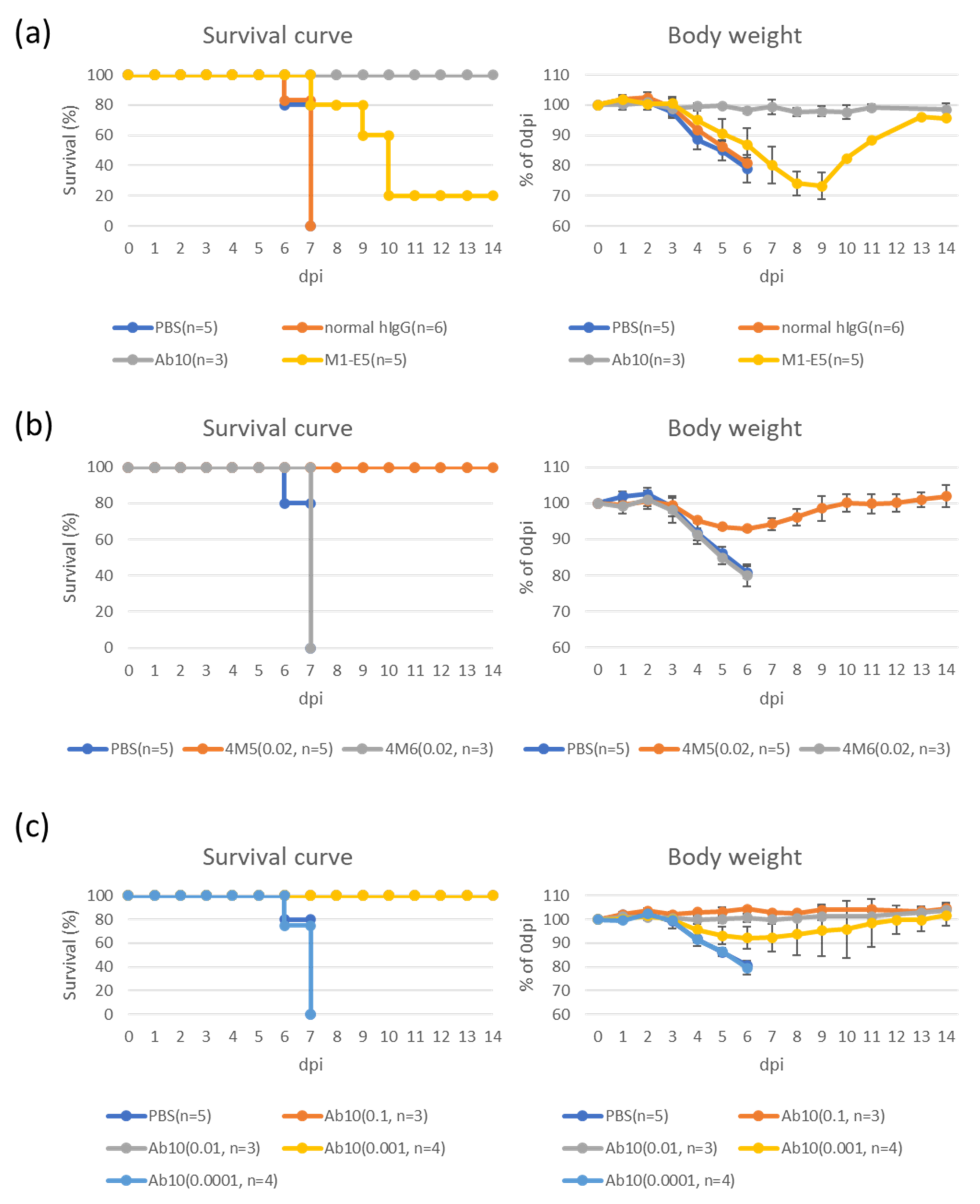
3.2. Therapeutic Effects of Selected mAbs in an SFTS Animal Model
3.3. Epitopes of Anti-Gn mAbs
3.4. Epitopes of Anti-Gc mAbs
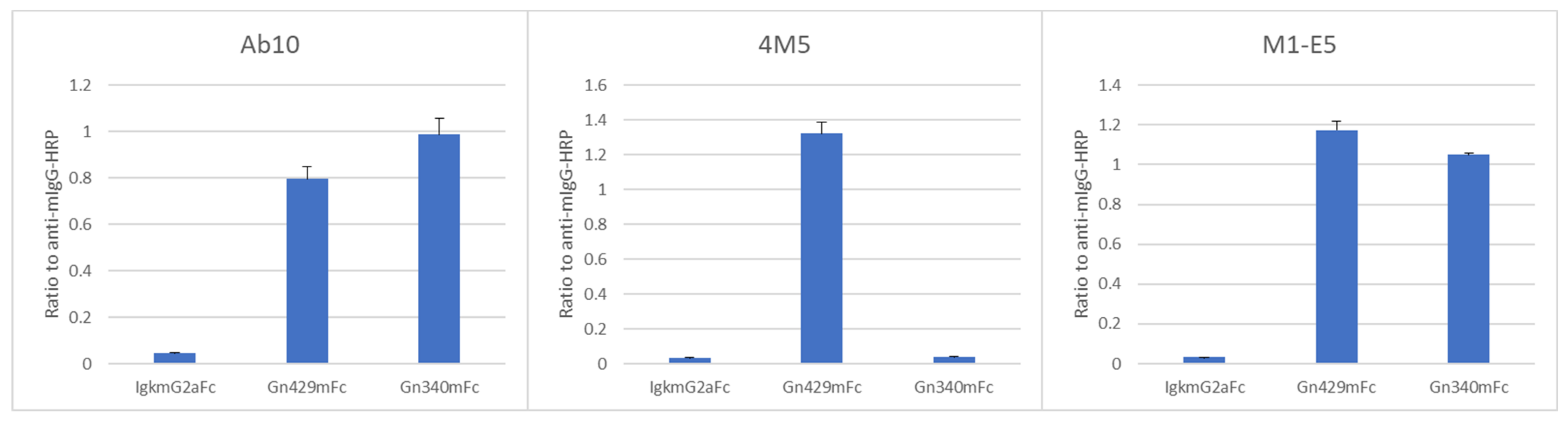
3.5. A Different Property of Neutralizing Anti-Gn/Gc mAbs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Yu, X.J.; Liang, M.F.; Zhang, S.Y.; Liu, Y.; Li, J.D.; Sun, Y.L.; Zhang, L.; Zhang, Q.F.; Popov, V.L.; Li, C.; et al. Fever with thrombocytopenia associated with a novel bunyavirus in China. N. Engl. J. Med. 2011, 364, 1523–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Liu, L.; Huang, X.; Ma, H.; Zhang, Y.; Du, Y.; Wang, P.; Tang, X.; Wang, H.; Kang, K.; et al. Metagenomic analysis of fever, thrombocytopenia and leukopenia syndrome (FTLS) in Henan Province, China: Discovery of a new bunyavirus. PLoS Pathog. 2011, 7, e1002369. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jiang, X.M.; Cui, N.; Yuan, C.; Zhang, S.F.; Lu, Q.B.; Yang, Z.D.; Xin, Q.L.; Song, Y.B.; Zhang, X.A.; et al. Clinical effect and antiviral mechanism of T-705 in treating severe fever with thrombocytopenia syndrome. Signal Transduct. Target. Ther. 2021, 6, 145. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Kato, H.; Yamagishi, T.; Shimada, T.; Matsui, T.; Yoshikawa, T.; Kurosu, T.; Shimojima, M.; Morikawa, S.; Hasegawa, H.; et al. SFTS Epidemiological Research Group Japan. Severe Fever with Thrombocytopenia Syndrome, Japan, 2013–2017. Emerg. Infect. Dis. 2020, 26, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Lu, Q.B.; Yao, W.S.; Zhao, J.; Zhang, X.A.; Cui, N.; Yuan, C.; Yang, T.; Peng, X.F.; Lv, S.M.; et al. Clinical efficacy and safety evaluation of favipiravir in treating patients with severe fever with thrombocytopenia syndrome. EBioMedicine 2021, 72, 103591. [Google Scholar] [CrossRef]
- Suemori, K.; Saijo, M.; Yamanaka, A.; Himeji, D.; Kawamura, M.; Haku, T.; Hidaka, M.; Kamikokuryo, C.; Kakihana, Y.; Azuma, T.; et al. A multicenter non-randomized, uncontrolled single arm trial for evaluation of the efficacy and the safety of the treatment with favipiravir for patients with severe fever with thrombocytopenia syndrome. PLoS Negl. Trop. Dis. 2021, 15, e0009103. [Google Scholar] [CrossRef]
- Zhao, W.; Zhen, J.-C. Analysis of the utilization of oral antihypertensive drugs in more than 40 hospitals from Beijing area during 2008–2011. China Pharm. 2013, 24, 506–509. [Google Scholar]
- Li, X.-P.; Yu, Z.-Q.; Liu, D. Analysis of the application of antihypertensive drugs in 34 hospitals of Wuhan city during 2011 to 2013. China Pharm. 2015, 20, 2748–2751. [Google Scholar]
- Li, H.; Zhang, L.K.; Li, S.F.; Zhang, S.F.; Wan, W.W.; Zhang, Y.L.; Xin, Q.L.; Dai, K.; Hu, Y.Y.; Wang, Z.B.; et al. Calcium channel blockers reduce severe fever with thrombocytopenia syndrome virus (SFTSV) related fatality. Cell Res 2019, 29, 739–753. [Google Scholar] [CrossRef]
- Urata, S.; Yasuda, J.; Iwasaki, M. Loperamide Inhibits Replication of Severe Fever with Thrombocytopenia Syndrome Virus. Viruses 2021, 13, 869. [Google Scholar] [CrossRef]
- Smee, D.F.; Jung, K.H.; Westover, J.; Gowen, B.B. 2’-Fluoro-2’-deoxycytidine is a broad-spectrum inhibitor of bunyaviruses in vitro and in phleboviral disease mouse models. Antivir. Res. 2018, 160, 48–54. [Google Scholar] [CrossRef]
- Xu, S.; Jiang, N.; Nawaz, W.; Liu, B.; Zhang, F.; Liu, Y.; Wu, X.; Wu, Z. Infection of humanized mice with a novel phlebovirus presented pathogenic features of severe fever with thrombocytopenia syndrome. PLoS Pathog. 2021, 17, e1009587. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, Y.; Gao, F.; Jiao, Y.; Oladejo, B.O.; Chai, Y.; Bi, Y.; Lu, S.; Dong, M.; Zhang, C.; et al. Structures of phlebovirus glycoprotein Gn and identification of a neutralizing antibody epitope. Proc. Natl. Acad. Sci. USA 2017, 114, E7564–E7573. [Google Scholar] [CrossRef] [Green Version]
- Halldorsson, S.; Behrens, A.J.; Harlos, K.; Huiskonen, J.T.; Elliott, R.M.; Crispin, M.; Brennan, B.; Bowden, T.A. Structure of a phleboviral envelope glycoprotein reveals a consolidated model of membrane fusion. Proc. Natl. Acad. Sci. USA 2016, 113, 7154–7159. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, S.; Fukuma, A.; Tani, H.; Fukushi, S.; Saijo, M.; Shimojima, M. A neutralization assay with a severe fever with thrombocytopenia syndrome virus strain that makes plaques in inoculated cells. J. Virol. Methods 2017, 244, 4–10. [Google Scholar] [CrossRef]
- Tani, H.; Kawachi, K.; Kimura, M.; Taniguchi, S.; Shimojima, M.; Fukushi, S.; Igarashi, M.; Morikawa, S.; Saijo, M. Identification of the amino acid residue important for fusion of severe fever with thrombocytopenia syndrome virus glycoprotein. Virology 2019, 535, 102–110. [Google Scholar] [CrossRef]
- Lewis, G.K.; Pazgier, M.; Evans, D.T.; Ferrari, G.; Bournazos, S.; Parsons, M.S.; Bernard, N.F.; Finzi, A. Beyond Viral Neutralization. AIDS Res. Hum. Retrovir. 2017, 33, 760–764. [Google Scholar] [CrossRef]
- Bradfute, S.B. The discovery and development of novel treatment strategies for filoviruses. Expert Opin. Drug Discov. 2022, 17, 139–149. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, L.; Zhang, W.; Chi, Y.; Zeng, X.; Li, X.; Qi, X.; Jin, Q.; Zhang, X.; Huang, M.; et al. Human antibody neutralizes severe Fever with thrombocytopenia syndrome virus, an emerging hemorrhagic Fever virus. Clin. Vaccine Immunol. 2013, 20, 1426–1432. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Kim, J.; Ko, M.; Chun, J.Y.; Kim, H.; Kim, S.; Min, J.Y.; Park, W.B.; Oh, M.D.; Chung, J. An anti-Gn glycoprotein antibody from a convalescent patient potently inhibits the infection of severe fever with thrombocytopenia syndrome virus. PLoS Pathog. 2019, 15, e1007375. [Google Scholar] [CrossRef] [Green Version]
- Shimada, S.; Posadas-Herrera, G.; Aoki, K.; Morita, K.; Hayasaka, D. Therapeutic effect of post-exposure treatment with antiserum on severe fever with thrombocytopenia syndrome (SFTS) in a mouse model of SFTS virus infection. Virology 2015, 482, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Fukuma, A.; Fukushi, S.; Yoshikawa, T.; Tani, H.; Taniguchi, S.; Kurosu, T.; Egawa, K.; Suda, Y.; Singh, H.; Nomachi, T.; et al. Severe Fever with Thrombocytopenia Syndrome Virus Antigen Detection Using Monoclonal Antibodies to the Nucleocapsid Protein. PLoS Negl. Trop. Dis. 2016, 10, e0004595. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar]
- Matsuura, Y.; Possee, R.D.; Overton, H.A.; Bishop, D.H. Baculovirus expression vectors: The requirements for high level expression of proteins, including glycoproteins. J. Gen. Virol. 1987, 68, 1233–1250. [Google Scholar] [CrossRef]
- Shimojima, M.; Morikawa, S.; Maeda, K.; Tohya, Y.; Miyazawa, T.; Mikami, T. Generation of monoclonal antibodies against a feline CD antigen (CD4) expressed by a recombinant baculovirus. J. Vet. Med. Sci. 1997, 59, 467–469. [Google Scholar] [CrossRef] [Green Version]
- Takimoto, K.; Taharaguchi, M.; Morikawa, S.; Ike, F.; Yamada, Y. Detection of the antibody to lymphocytic choriomeningitis virus in sera of laboratory rodents infected with viruses of laboratory and newly isolated strains by ELISA using purified recombinant nucleoprotein. Exp. Anim. 2008, 57, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Onodera, T.; Kita, S.; Adachi, Y.; Moriyama, S.; Sato, A.; Nomura, T.; Sakakibara, S.; Inoue, T.; Tadokoro, T.; Anraku, Y.; et al. A SARS-CoV-2 antibody broadly neutralizes SARS-related coronaviruses and variants by coordinated recognition of a virus-vulnerable site. Immunity 2021, 54, 2385–2398. [Google Scholar] [CrossRef]
- Shimojima, M.; Sugimoto, S.; Taniguchi, S.; Yoshikawa, T.; Kurosu, T.; Saijo, M. Efficient functional screening of a cellular cDNA library to identify severe fever with thrombocytopenia syndrome virus entry factors. Sci. Rep. 2020, 10, 5996. [Google Scholar] [CrossRef]
- Kimura, M.; Egawa, K.; Ozawa, T.; Kishi, H.; Shimojima, M.; Taniguchi, S.; Fukushi, S.; Fujii, H.; Yamada, H.; Tan, L.; et al. Characterization of pseudotyped vesicular stomatitis virus bearing the heartland virus envelope glycoprotein. Virology 2021, 556, 124–132. [Google Scholar] [CrossRef]
- Tani, H.; Shimojima, M.; Fukushi, S.; Yoshikawa, T.; Fukuma, A.; Taniguchi, S.; Morikawa, S.; Saijo, M. Characterization of Glycoprotein-Mediated Entry of Severe Fever with Thrombocytopenia Syndrome Virus. J. Virol. 2016, 90, 5292–5301. [Google Scholar] [CrossRef] [Green Version]
- Phanthanawiboon, S.; Limkittikul, K.; Sakai, Y.; Takakura, N.; Saijo, M.; Kurosu, T. Acute Systemic Infection with Dengue Virus Leads to Vascular Leakage and Death through Tumor Necrosis Factor-α and Tie2/Angiopoietin Signaling in Mice Lacking Type I and II Interferon Receptors. PLoS ONE 2016, 11, e0148564. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clones | Isotypes | ELISA EC50, µg/mL | NT IC50, µg/mL | Therapeutic Effects with Doses, mg/day | References |
|---|---|---|---|---|---|
| No. 2 | Mouse IgG1 | 0.14 | 16< | ND 1 | Present study |
| No. 4 | Mouse IgG1 | 0.090 | 16< | ND | Present study |
| No. 5 | Mouse IgG1 | 0.11 | 16< | ND | Present study |
| No. 11 | Mouse IgG1 | 0.10 | 16< | ND | Present study |
| 4M2 | Mouse IgG2b | 0.12 | 16< | ND | Immune Technology Corp. |
| 4M3 | Mouse IgG2a | 0.046 | 16< | ND | Immune Technology Corp. |
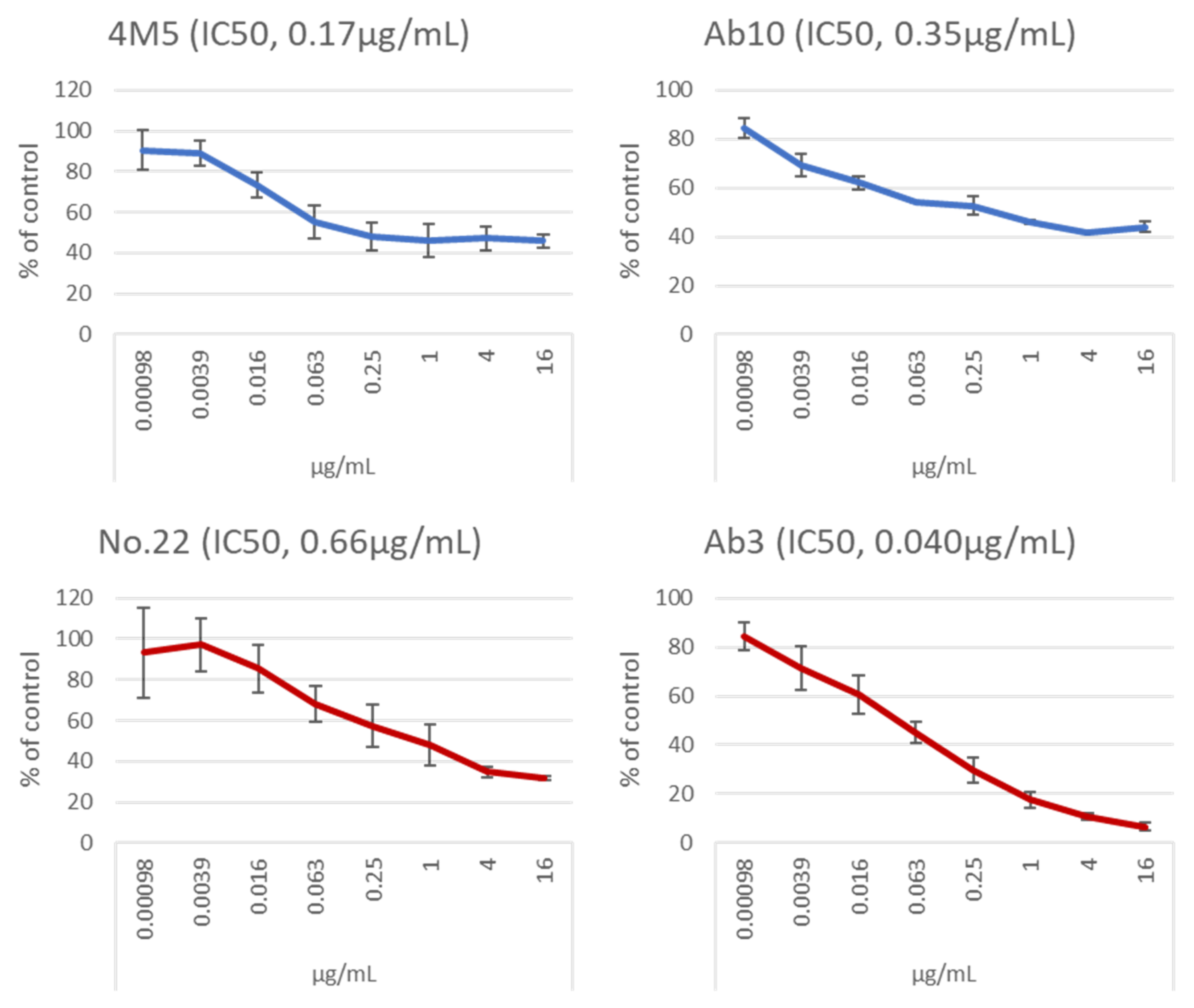
| 4M5 | Mouse IgG1 | 0.16 | 0.17 | Y 2 at 0.02 | Immune Technology Corp. |
| 4M6 | Mouse IgG1 | 0.067 | 16< | N 3 at 0.02 | Immune Technology Corp. |
| 4M7 | Mouse IgG1 | 0.062 | 16< | ND | Immune Technology Corp. |
| 4M8 | Mouse IgG2b | 0.016 | 16< | ND | Immune Technology Corp. |
| M1-B8 | Human IgG1 | 0.056 | 16< | N at 1 | Present study |
| M1-D1 | Human IgG1 | 0.061 | 16< | N at 1 | Present study |
| M1-E1 | Human IgG1 | 0.051 | 16< | N at 1 | Present study |
| M1-E5 | Human IgG1 | 0.046 | 16< | Y at 1 | Present study |
| Ab10 | Human IgG1 | 0.027 | 0.35 | Y at 1, 0.1, 0.01, 0.001N at 0.0001 | [20] |
| MAb4-5 | Human IgG1 | 0.48 | 16< | N at 1 | [19] |
| Clones | Isotypes | ELISA EC50, µg/mL | NT IC50, µg/mL | Therapeutic Effects with Doses, mg/day | References |
|---|---|---|---|---|---|
| No. 16 | Mouse IgG1 | 0.18 | 16< | ND 1 | Present study |
| No. 22 | Mouse IgG2b | 0.077 | 0.66 | N 2 at 1 | Present study |
| No. 23 | Mouse IgG1 | 0.96 | 1.1 | ND | Present study |
| No. 31 | Mouse IgG1 | 0.11 | 16< | ND | Present study |
| No. 33 | Mouse IgG1 | 0.61 | 16< | ND | Present study |
| No. 40 | Mouse IgG1 | 0.49 | 16< | ND | Present study |
| 5D12 | Mouse IgG1 | 0.14 | 16< | N at 1 | Present study |
| 6D12 | Mouse IgG2a | 0.50 | 16< | N at 1 | Present study |
| 8E9 | Mouse IgG1 | 2.2 | 16< | N at 1 | Present study |
| 5M1 | Mouse IgG1 | 0.22 | 16< | ND | Immune Technology Corp. |
| 5M3 | Mouse IgG1 | 0.045 | 16< | ND | Immune Technology Corp. |
| 5M5 | Mouse IgG1 | 0.094 | 3.9 | ND | Immune Technology Corp. |
| 5M9 | Mouse IgG1 | 0.037 | 2.1 | ND | Immune Technology Corp. |
| C3A11 | Mouse IgG2a | 0.048 | 16< | N at 1 | [29] |
| C6C1 | Mouse IgG1 | 0.053 | 1.8 | N at 1 | [29] |
| Ab3 | Human IgG1 | 0.054 | 0.040 | N at 1 | Patent (#10947299) |
| Competitors 2 | Binding of Biotinylated Ab10 | Binding of Biotinylated 4M5 | Binding of Biotinylated M1-E5 |
|---|---|---|---|
| Ab10 | Reduced | Not reduced | Not reduced |
| 4M5, 4M6, 4M8 | Not reduced | Reduced | Not reduced |
| M1-B8, M1-D1, M1-E1, M1-E5 | Not reduced | Not reduced | Reduced 1 |
| Competitors 2 | Binding of Biotinylated No. 22 | Binding of Biotinylated C6C1 | Binding of Biotinylated Ab3 |
|---|---|---|---|
| No. 22, No. 23, 5M9, C3A11 | Reduced | Not reduced | Not reduced |
| C6C1 | Not reduced | Reduced | Not reduced |
| Ab3 | Not reduced | Not reduced | Reduced 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimojima, M.; Sugimoto, S.; Umekita, K.; Onodera, T.; Sano, K.; Tani, H.; Takamatsu, Y.; Yoshikawa, T.; Kurosu, T.; Suzuki, T.; et al. Neutralizing mAbs against SFTS Virus Gn Protein Show Strong Therapeutic Effects in an SFTS Animal Model. Viruses 2022, 14, 1665. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081665
Shimojima M, Sugimoto S, Umekita K, Onodera T, Sano K, Tani H, Takamatsu Y, Yoshikawa T, Kurosu T, Suzuki T, et al. Neutralizing mAbs against SFTS Virus Gn Protein Show Strong Therapeutic Effects in an SFTS Animal Model. Viruses. 2022; 14(8):1665. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081665
Chicago/Turabian StyleShimojima, Masayuki, Satoko Sugimoto, Kunihiko Umekita, Taishi Onodera, Kaori Sano, Hideki Tani, Yuki Takamatsu, Tomoki Yoshikawa, Takeshi Kurosu, Tadaki Suzuki, and et al. 2022. "Neutralizing mAbs against SFTS Virus Gn Protein Show Strong Therapeutic Effects in an SFTS Animal Model" Viruses 14, no. 8: 1665. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081665