
Properties of Adenovirus Vectors with Increased Affinity to DSG2 and the Potential Benefits of Oncolytic Approaches and Gene Therapy

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vector Production and Titration
2.2. Cell Cultures
2.2.1. T84 Cells in Transwell Culture
2.2.2. Spheroid Culture
2.3. Analyzing Transduction Efficiencies via GFP Positive Cells
2.4. Immunostaining for Receptor Detection
2.5. Trans-Epithelial Electrical Resistance (TEER) Measurement of T84 Transwell Culture
2.6. Histological Staining of T84 Transwell Culture
2.7. Virus Cellular Entry Measured by Internalization Assay
2.8. Quantitative PCR Analysis for Adenovirus Genome Quantification
2.9. Oncolytic Assay
2.10. Virus Spreading Ability Detected by Plaque Assay
2.11. Silver Gel for the Confirmation of Virus Protein Expression
2.12. Statistics
3. Results
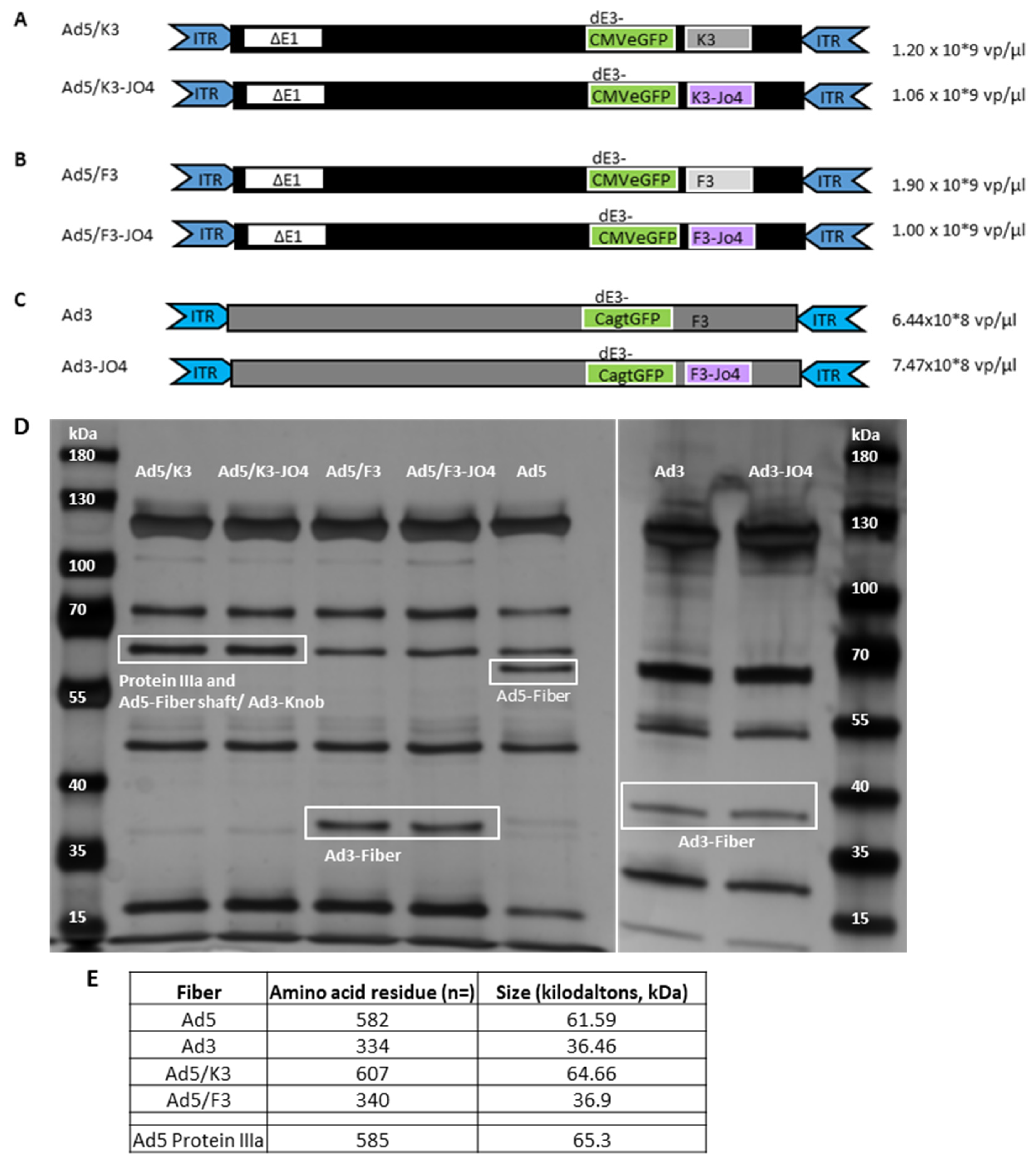
3.1. Construction of JO4 Containing Vectors
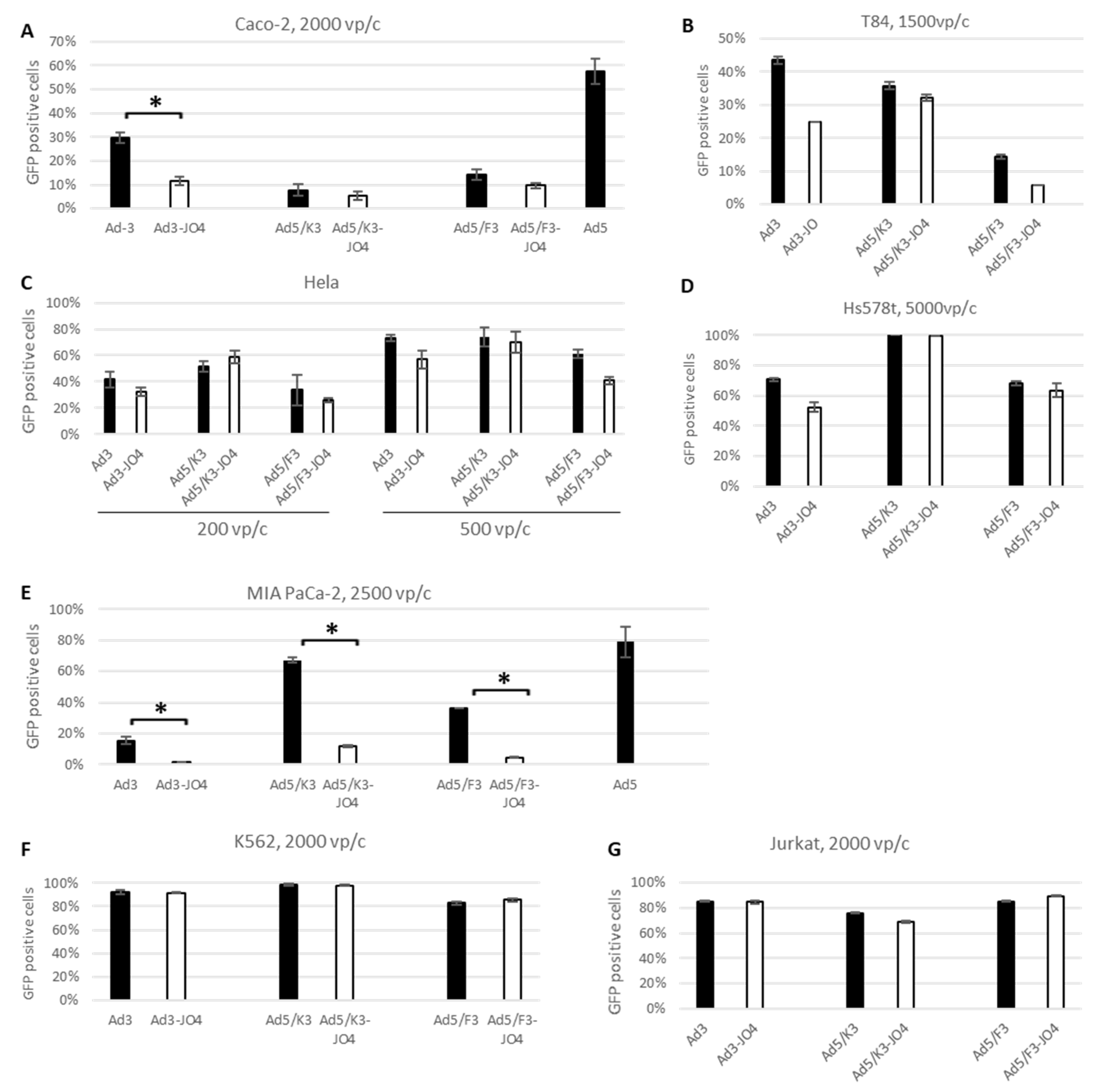
3.2. Transduction Efficiency Measured by Transgene Expression in Human Cancer Cell Lines
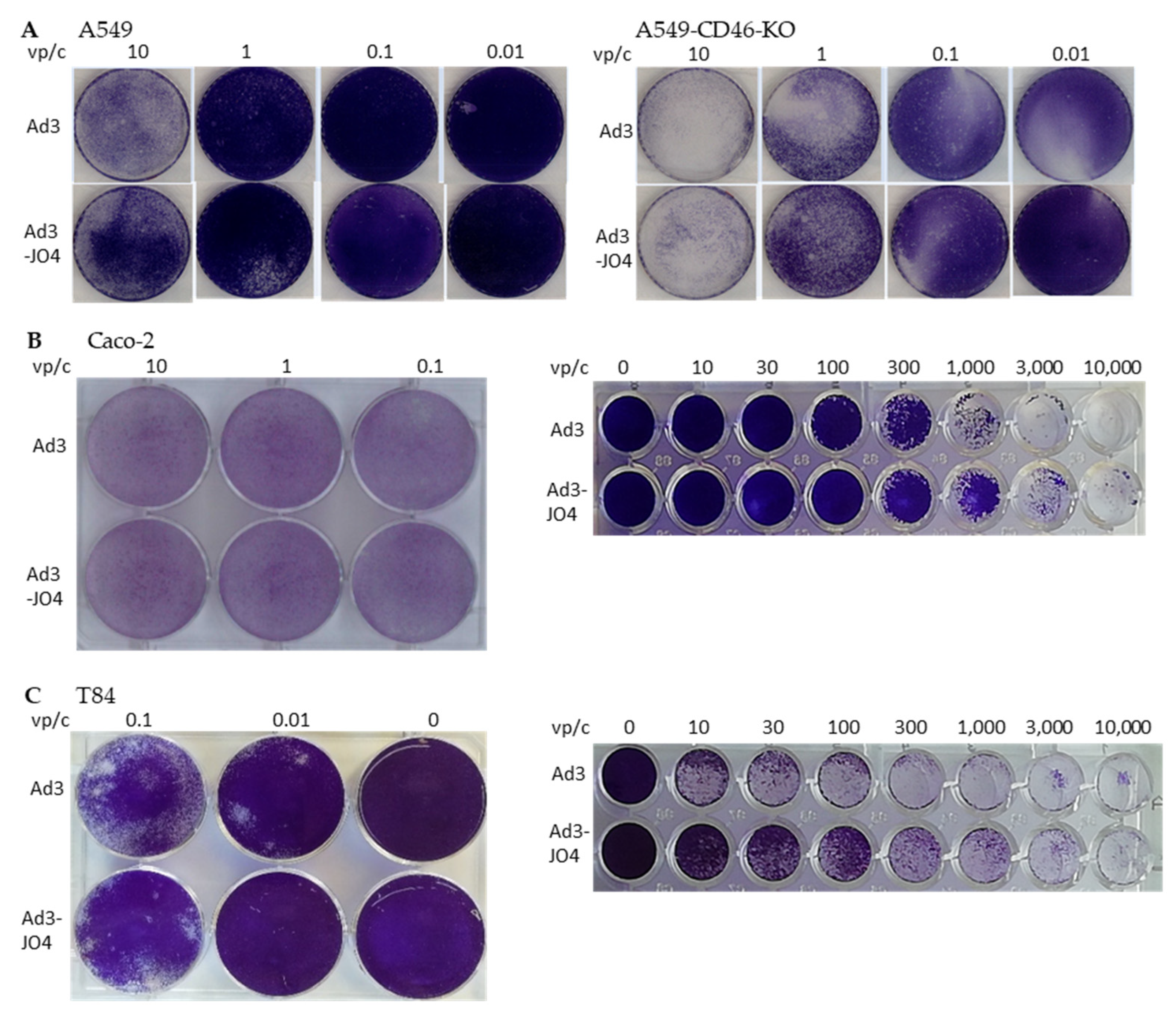
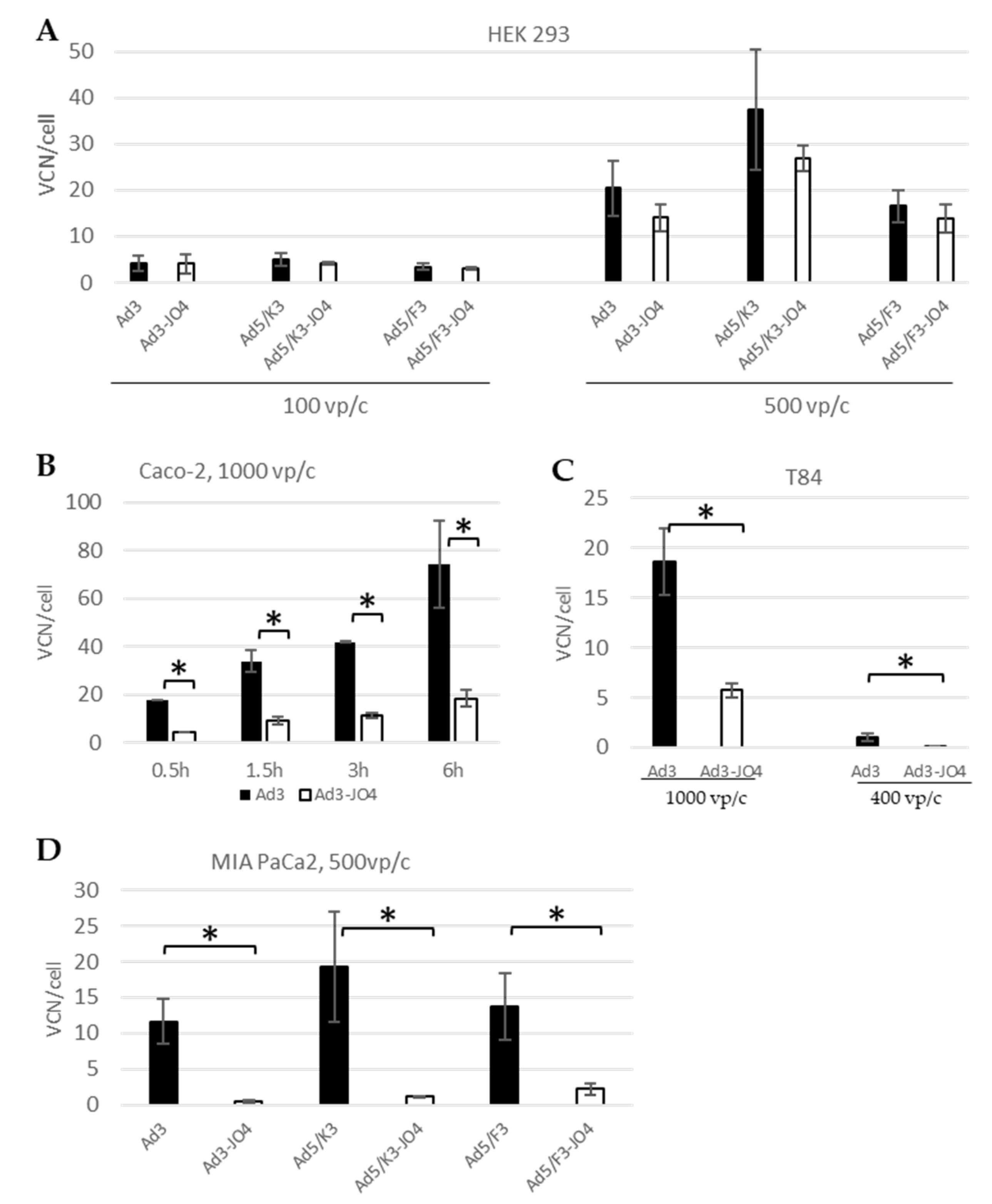
3.3. Virus Spreading in a Two-Dimensional (2D) Cell Culture Model
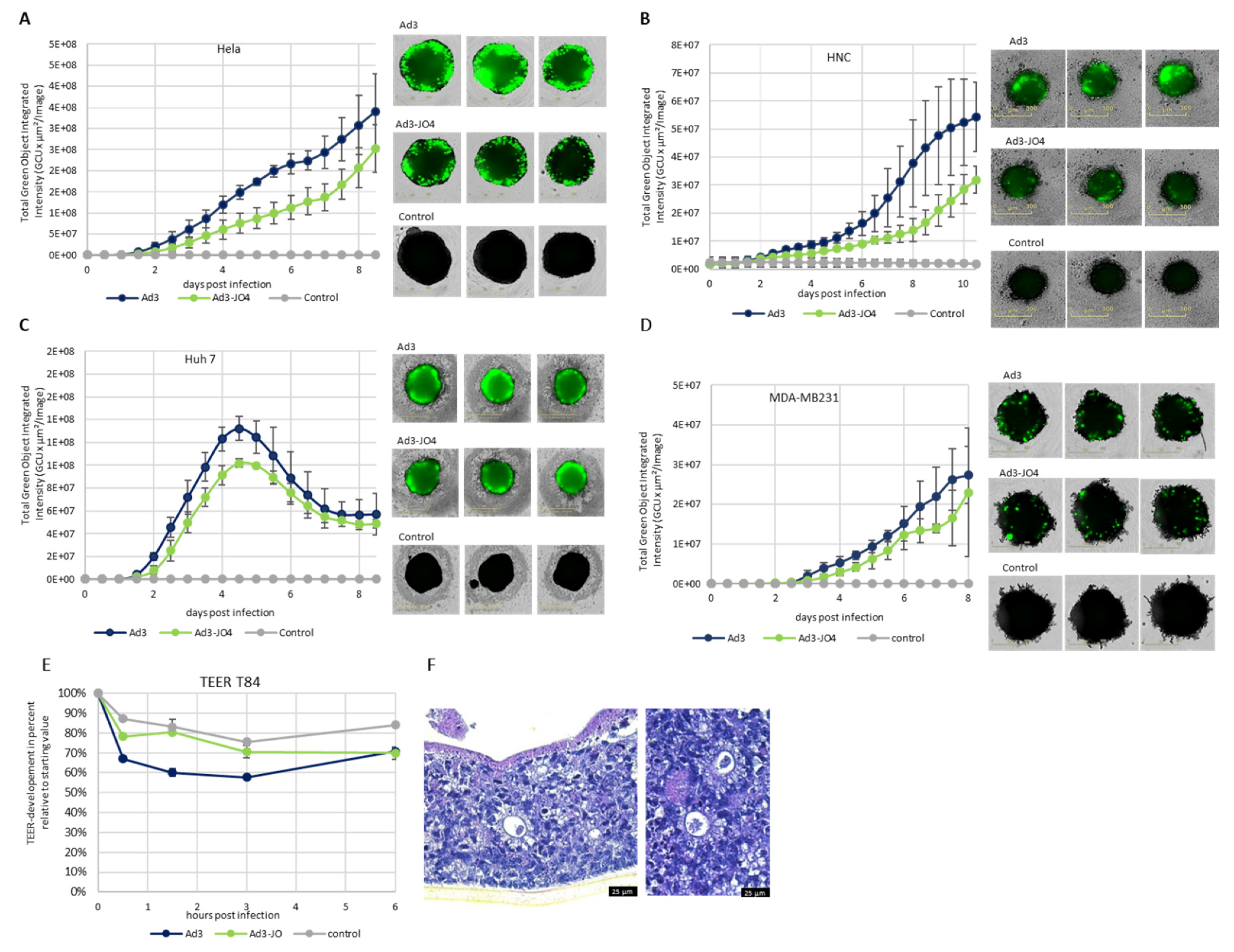
3.4. Performance of JO4 Containing Vectors in a Three-Dimensional (3D) Cell Culture Model
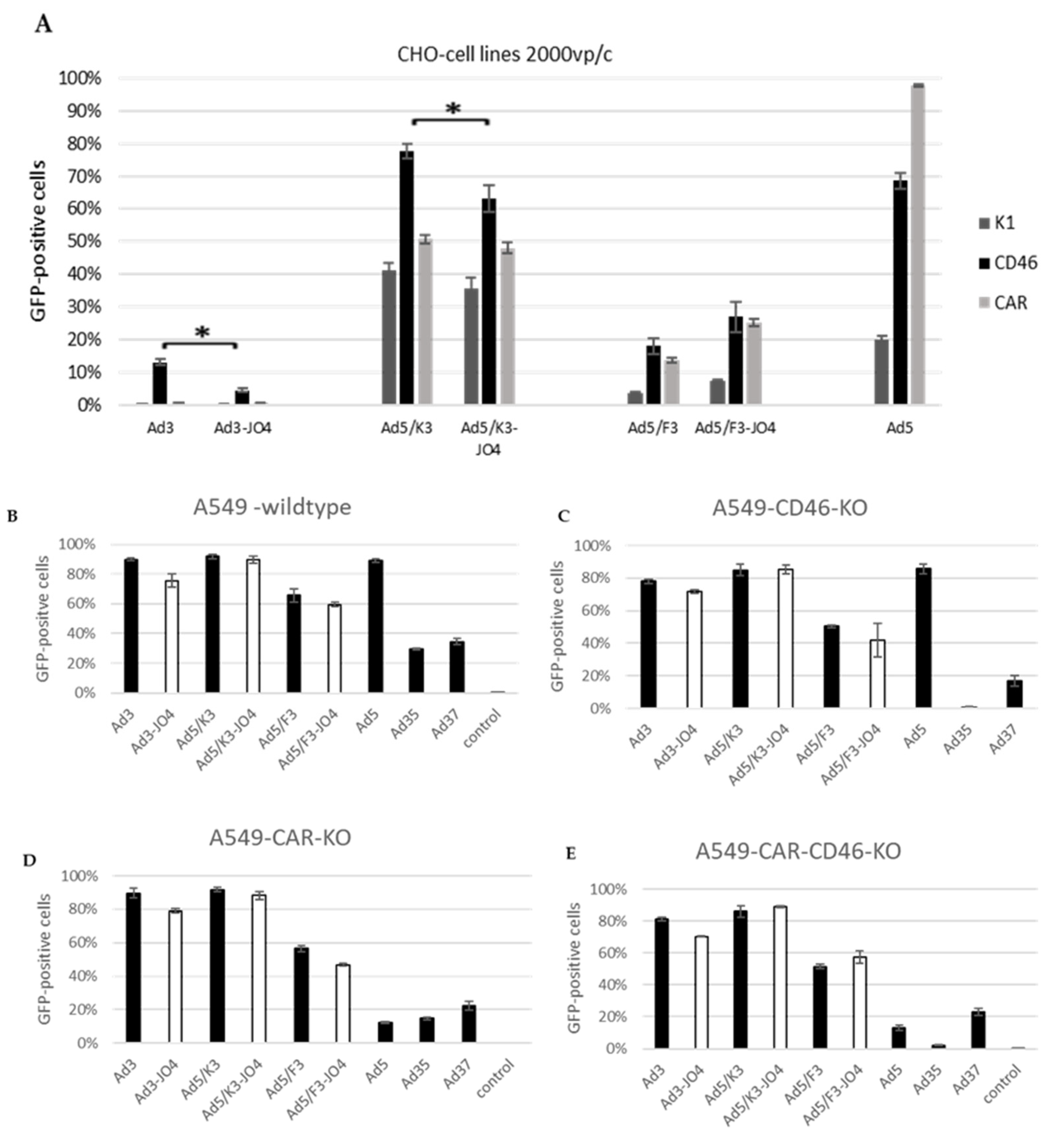
3.5. Effect of JO4 on CD46 and CAR Receptor Expressing CHO Cells and Receptor Knockout A549 Cells
3.6. Cellular Entry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berman, J.J. Tumor taxonomy for the developmental lineage classification of neoplasms. BMC Cancer 2004, 4, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leech, A.O.; Cruz, R.G.; Hill, A.D.; Hopkins, A.M. Paradigms lost—An emerging role for over-expression of tight junction adhesion proteins in cancer pathogenesis. Ann. Transl. Med. 2015, 3, 184. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Earnshaw, W.C.; Lippincott-Schwartz, J.; Johnson, G. Intercellular Junctions. In Cell Biology; Pollard, T.D., Earnshaw, W.C., Lippincott-Schwartz, J., Johnson, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 543–553. [Google Scholar]
- Green, K.J.; Gaudry, C.A. Are desmosomes more than tethers for intermediate filaments? Nat. Rev. Mol. Cell Biol. 2000, 1, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Green, C.L.S.K.J.; Simpson, C.L. Desmosomes: New Perspectives on a Classic. J. Investig. Dermatol. 2007, 127, 2499–2515. [Google Scholar] [CrossRef] [Green Version]
- Chitaev, N.A.; Troyanovsky, S.M. Direct Ca2+-dependent Heterophilic Interaction between Desmosomal Cadherins, Desmoglein and Desmocollin, Contributes to Cell-Cell Adhesion. J. Cell Biol. 1997, 138, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Yashiro, M.; Nishioka, N.; Hirakawa, K. Decreased expression of the adhesion molecule desmoglein-2 is associated with diffuse-type gastric carcinoma. Eur. J. Cancer 2006, 42, 2397–2403. [Google Scholar] [CrossRef]
- Ramani, V.C.; Hennings, L.; Haun, R.S. Desmoglein 2 is a substrate of kallikrein 7 in pancreatic cancer. BMC Cancer 2008, 8, 373. [Google Scholar] [CrossRef] [Green Version]
- Barber, A.G.; Castillo-Martin, M.; Bonal, D.; Rybicki, B.A.; Christiano, A.M.; Cordon-Cardo, C. Characterization of Desmoglein Expression in the Normal Prostatic Gland. Desmoglein 2 Is an Independent Prognostic Factor for Aggressive Prostate Cancer. PLoS ONE 2014, 9, e98786. [Google Scholar] [CrossRef]
- Brennan, D.; Mahoney, M.G. Increased expression of Dsg2 in malignant skin carcinomas: A tissue-microarray based study. Cell Adhes. Migr. 2009, 3, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Cai, F.; Zhu, Q.; Miao, Y.; Shen, S.; Su, X.; Shi, Y. Desmoglein-2 is overexpressed in non-small cell lung cancer tissues and its knockdown suppresses NSCLC growth by regulation of p27 and CDK2. J. Cancer Res. Clin. Oncol. 2016, 143, 59–69. [Google Scholar] [CrossRef]
- Han, C.-P.; Yu, Y.-H.; Wang, A.-G.; Tian, Y.; Zhang, H.-T.; Zheng, Z.-M.; Liu, Y.-S. Desmoglein-2 overexpression predicts poor prognosis in hepatocellular carcinoma patients. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5481–5489. [Google Scholar]
- Tan, L.Y.; Mintoff, C.; Johan, M.Z.; Ebert, B.W.; Fedele, C.; Zhang, Y.F.; Szeto, P.; Sheppard, K.E.; McArthur, G.A.; Foster-Smith, E.; et al. Desmoglein 2 promotes vasculogenic mimicry in melanoma and is associated with poor clinical outcome. Oncotarget 2016, 7, 46492–46508. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Beidler, P.; Wang, H.; Li, C.; Quassab, A.; Coles, C.; Drescher, C.; Carter, D.; Lieber, A. Desmoglein-2 as a prognostic and biomarker in ovarian cancer. Cancer Biol. Ther. 2020, 21, 1154–1162. [Google Scholar] [CrossRef]
- Ebert, L.M.; Vandyke, K.; Johan, M.Z.; DeNichilo, M.; Tan, L.Y.; Min, K.K.M.; Weimann, B.M.; Ebert, B.W.; Pitson, S.M.; Zannettino, A.C.W.; et al. Desmoglein-2 expression is an independent predictor of poor prognosis patients with multiple myeloma. Mol. Oncol. 2021, 16, 1221–1240. [Google Scholar] [CrossRef]
- Mateo, M.; Generous, A.; Sinn, P.; Cattaneo, R. Connections matter—How viruses use cell-cell adhesion components. J. Cell Sci. 2015, 128, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, Z.-Y.; Liu, Y.; Persson, J.; Beyer, I.; Möller, T.; Koyuncu, D.; Drescher, M.R.; Strauss, R.; Zhang, X.-B.; et al. Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat. Med. 2010, 17, 96–104. [Google Scholar] [CrossRef]
- Feng, Y.; Yi, C.; Liu, X.; Qu, L.; Su, W.; Shu, T.; Zheng, X.; Ye, X.; Luo, J.; Hao, M.; et al. Human Desmoglein-2 and Human CD46 Mediate Human Adenovirus Type 55 Infection, but Human Desmoglein-2 Plays the Major Roles. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Wang, H.; Ducournau, C.; Saydaminova, K.; Richter, M.; Yumul, R.; Ho, M.; Carter, D.; Zubieta, C.; Fender, P.; Lieber, A. Intracellular Signaling and Desmoglein 2 Shedding Triggered by Human Adenoviruses Ad3, Ad14, and Ad14P1. J. Virol. 2015, 89, 10841–10859. [Google Scholar] [CrossRef] [Green Version]
- Coyne, C.B.; Bergelson, J.M. CAR: A virus receptor within the tight junction. Adv. Drug Deliv. Rev. 2005, 57, 869–882. [Google Scholar] [CrossRef]
- Fender, P.; Boussaid, A.; Mezin, P.; Chroboczek, J. Synthesis, cellular localization, and quantification of penton-dodecahedron in serotype 3 adenovirus-infected cells. Virology 2005, 340, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yumul, R.; Cao, H.; Ran, L.; Fan, X.; Richter, M.; Epstein, F.; Gralow, J.; Zubieta, C.; Fender, P.; et al. Structural and Functional Studies on the Interaction of Adenovirus Fiber Knobs and Desmoglein 2. J. Virol. 2013, 87, 11346–11362. [Google Scholar] [CrossRef] [Green Version]
- Pitner, R.; Kim, J.; Davis-Bergthold, J.; Turner, C.; Stermann, E.; Wang, H.; Adams, J.; Carter, L.; Ahlgren, J.A.; Fender, P.; et al. Structure-based Design of JOC-x, a Conjugatable Tumor Tight Junction Opener to Enhance Cancer Therapy. Sci. Rep. 2019, 9, 6169. [Google Scholar] [CrossRef]
- Stermann, E.; Mottet, M.; Ducournau, C.; Iseni, F.; Vragniau, C.; Wang, H.; Zubieta, C.; Lieber, A.; Fender, P. Mapping of Adenovirus of serotype 3 fibre interaction to desmoglein 2 revealed a novel ‘non-classical’ mechanism of viral receptor engagement. Sci. Rep. 2018, 8, 8381. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Yumul, R.; Wang, H.; Saydaminova, K.; Ho, M.; May, D.; Baldessari, A.; Gough, M.; Drescher, C.; Urban, N.; et al. Preclinical safety and efficacy studies with an affinity-enhanced epithelial junction opener and PEGylated liposomal doxorubicin. Mol. Ther.-Methods Clin. Dev. 2015, 2, 15005. [Google Scholar] [CrossRef]
- Beyer, I.; Cao, H.; Persson, J.; Song, H.; Richter, M.; Feng, Q.; Yumul, R.; van Rensburg, R.; Li, Z.; Berenson, R.; et al. Coadministration of Epithelial Junction Opener JO-1 Improves the Efficacy and Safety of Chemotherapeutic Drugs. Clin. Cancer Res. 2012, 18, 3340–3351. [Google Scholar] [CrossRef] [Green Version]
- Beyer, I.; Van Rensburg, R.; Lieber, A. Overcoming physical barriers in cancer therapy. Tissue Barriers 2013, 1, e23647. [Google Scholar] [CrossRef] [Green Version]
- Lieber, A.; Wang, H.; Germond, A.; Li, C.; Gil, S.; Kim, J.; Kiem, H.-P. In vivo HSC transduction in rhesus macaques with an HDAd5/3+ vector targeting desmoglein 2 and transiently over-expressing cxcr4. Blood Adv. 2022, 6, 4360–4372. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Li, Z.; Tuve, S.; Stone, D.; Kalyushniy, O.; Shayakhmetov, D.; Verlinde, C.L.M.; Stehle, T.; McVey, J.; et al. In Vitro and in Vivo Properties of Adenovirus Vectors with Increased Affinity to CD46. J. Virol. 2008, 82, 10567–10579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, S.; Uusi-Kerttula, H.; Jones, R.; Hanna, L.; Chester, J.D.; Parker, A.L. Evaluation of CD46 re-targeted adenoviral vectors for clinical ovarian cancer intraperitoneal therapy. Cancer Gene Ther. 2016, 23, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Saydaminova, K.; Yumul, R.; Krishnan, R.; Liu, J.; Nagy, E.-E.; Singh, M.; Izsvak, Z.; Cattaneo, R.; Uckert, W.; et al. In vivo transduction of primitive mobilized hematopoietic stem cells after intravenous injection of integrating adenovirus vectors. Blood 2016, 128, 2206–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Psatha, N.; Wang, H.; Singh, M.; Samal, H.B.; Zhang, W.; Ehrhardt, A.; Izsvák, Z.; Papayannopoulou, T.; Lieber, A. Integrating HDAd5/35++ Vectors as a New Platform for HSC Gene Therapy of Hemoglobinopathies. Mol. Ther.-Methods Clin. Dev. 2018, 9, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Trinh, H.V.; Lesage, G.; Chennamparampil, V.; Vollenweider, B.; Burckhardt, C.J.; Schauer, S.; Havenga, M.; Greber, U.F.; Hemmi, S. Avidity Binding of Human Adenovirus Serotypes 3 and 7 to the Membrane Cofactor CD46 Triggers Infection. J. Virol. 2012, 86, 1623–1637. [Google Scholar] [CrossRef] [Green Version]
- Gaggar, A.; Shayakhmetov, D.M.; Lieber, A. CD46 is a cellular receptor for group B adenoviruses. Nat. Med. 2003, 9, 1408–1412. [Google Scholar] [CrossRef]
- Zhang, W.; Fu, J.; Ehrhardt, A. Novel Vector Construction Based on Alternative Adenovirus Types via Homologous Recombination. Hum. Gene Ther. Methods 2018, 29, 124–134. [Google Scholar] [CrossRef]
- Zhang, W.; Fu, J.; Liu, J.; Wang, H.; Schiwon, M.; Janz, S.; Schaffarczyk, L.; von der Goltz, L.; Ehrke-Schulz, E.; Doerner, J.; et al. An Engineered Virus Library as a Resource for the Spectrum-wide Exploration of Virus and Vector Diversity. Cell Rep. 2017, 19, 1698–1709. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Bian, X.; Xia, L.; Ding, X.; Müller, R.; Zhang, Y.; Fu, J.; Stewart, A.F. Improved seamless mutagenesis by recombineering using ccdB for counterselection. Nucleic Acids Res. 2013, 42, e37. [Google Scholar] [CrossRef]
- Jager, L.; A Hausl, M.; Rauschhuber, C.; Wolf, N.M.; A Kay, M.; Ehrhardt, A. A rapid protocol for construction and production of high-capacity adenoviral vectors. Nat. Protoc. 2009, 4, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Baer, A.; Kehn-Hall, K. Viral Concentration Determination Through Plaque Assays: Using Traditional and Novel Overlay Systems. J. Vis. Exp. 2014, 93, e52065. [Google Scholar] [CrossRef]
- Blum, H.; Beier, H.; Gross, H.J. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. ELECTROPHORESIS 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Krutzke, L.; Prill, J.; Engler, T.; Schmidt, C.; Xu, Z.; Byrnes, A.; Simmet, T.; Kreppel, F. Substitution of blood coagulation factor X-binding to Ad5 by position-specific PEGylation: Preventing vector clearance and preserving infectivity. J. Control. Release 2016, 235, 379–392. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, Z.; Yumul, R.; Lara, S.; Hemminki, A.; Fender, P.; Lieber, A. Multimerization of Adenovirus Serotype 3 Fiber Knob Domains Is Required for Efficient Binding of Virus to Desmoglein 2 and Subsequent Opening of Epithelial Junctions. J. Virol. 2011, 85, 6390–6402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoukas, R.L.; Volkwein, W.; Gao, J.; Schiwon, M.; Bahlmann, N.; Dittmar, T.; Hagedorn, C.; Ehrke-Schulz, E.; Zhang, W.; Baiker, A.; et al. A Human In Vitro Model to Study Adenoviral Receptors and Virus Cell Interactions. Cells 2022, 11, 841. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.-Z.; Wang, H.; Zhang, Y.; Cao, H.; Li, Z.; Fender, P.; Lieber, A. Penton-Dodecahedral Particles Trigger Opening of Intercellular Junctions and Facilitate Viral Spread during Adenovirus Serotype 3 Infection of Epithelial Cells. PLoS Pathog. 2013, 9, e1003718. [Google Scholar] [CrossRef] [PubMed]
- Beyer, I.; van Rensburg, R.; Strauss, R.; Li, Z.; Wang, H.; Persson, J.; Yumul, R.; Feng, Q.; Song, H.; Bartek, J.; et al. Epithelial Junction Opener JO-1 Improves Monoclonal Antibody Therapy of Cancer. Cancer Res. 2011, 71, 7080–7090. [Google Scholar] [CrossRef] [Green Version]
- Yumul, R.; Richter, M.; Lu, Z.-Z.; Saydaminova, K.; Wang, H.; Wang, C.-H.K.; Carter, D.; Lieber, A. Epithelial Junction Opener Improves Oncolytic Adenovirus Therapy in Mouse Tumor Models. Hum. Gene Ther. 2016, 27, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Mese, K.; Bunz, O.; Schellhorn, S.; Volkwein, W.; Jung, D.; Gao, J.; Zhang, W.; Baiker, A.; Ehrhardt, A. Identification of novel human adenovirus candidates using the coxsackievirus and adenovirus receptor for cell entry. Virol. J. 2020, 17, 52. [Google Scholar] [CrossRef] [Green Version]
- Nemerow, G.R.; Stewart, P.L. Role of αv Integrins in Adenovirus Cell Entry and Gene Delivery. Microbiol. Mol. Biol. Rev. 1999, 63, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Madisch, I.; Hofmayer, S.; Moritz, C.; Grintzalis, A.; Hainmueller, J.; Pring-Akerblom, P.; Heim, A. Phylogenetic Analysis and Structural Predictions of Human Adenovirus Penton Proteins as a Basis for Tissue-Specific Adenovirus Vector Design. J. Virol. 2007, 81, 8270–8281. [Google Scholar] [CrossRef] [Green Version]
- Amstutz, B.; Gastaldelli, M.; Kälin, S.; Imelli, N.; Boucke, K.; Wandeler, E.; Mercer, J.; Hemmi, S.; Greber, U.F. Subversion of CtBP1-controlled macropinocytosis by human adenovirus serotype 3. EMBO J. 2008, 27, 956–969. [Google Scholar] [CrossRef] [Green Version]
- Nestić, D.; Božinović, K.; Pehar, I.; Wallace, R.; Parker, A.L.; Majhen, D. The Revolving Door of Adenovirus Cell Entry: Not All Pathways Are Equal. Pharmaceutics 2021, 13, 1585. [Google Scholar] [CrossRef]
- Ebert, L.; Tan, L.Y.; Johan, M.Z.; Min, K.K.M.; Cockshell, M.P.; Parham, K.A.; Betterman, K.; Szeto, P.; Boyle, S.; Silva, L.; et al. A non-canonical role for desmoglein-2 in endothelial cells: Implications for neoangiogenesis. Angiogenesis 2016, 19, 463–486. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, K.; Hu, C.; Nakamura, T.; Marks, J.D.; Russell, S.J.; Peng, K.-W. Affinity Thresholds for Membrane Fusion Triggering by Viral Glycoproteins. J. Virol. 2007, 81, 13149–13157. [Google Scholar] [CrossRef] [Green Version]
- Zafar, M.S.; Basnet, S.; Launonen, M.I.-M.; Quixabeira, M.D.C.A.; Santos, J.; Hemminki, O.; Malmstedt, M.M.; Cervera-Carrascon, V.; Aronen, P.; Kalliokoski, R.; et al. Oncolytic Adenovirus Type 3 Coding for CD40L Facilitates Dendritic Cell Therapy of Prostate Cancer in Humanized Mice and Patient Samples. Hum. Gene Ther. 2021, 32, 192–202. [Google Scholar] [CrossRef]
- Zafar, S.; Sorsa, S.; Siurala, M.; Hemminki, O.; Havunen, R.; Cervera-Carrascon, V.; Santos, J.M.; Wang, H.; Lieber, A.; De Gruijl, T.; et al. CD40L coding oncolytic adenovirus allows long-term survival of humanized mice receiving dendritic cell therapy. OncoImmunology 2018, 7, e1490856. [Google Scholar] [CrossRef] [Green Version]
- Kuryk, L.; Møller, A.W. Chimeric oncolytic Ad5/3 virus replicates and lyses ovarian cancer cells through desmoglein-2 cell entry receptor. J. Med. Virol. 2020, 92, 1309–1315. [Google Scholar] [CrossRef] [Green Version]
- Bramante, S.; Koski, A.; Liikanen, I.; Vassilev, L.; Oksanen, M.; Siurala, M.; Heiskanen, R.; Hakonen, T.; Joensuu, T.; Kanerva, A.; et al. Oncolytic virotherapy for treatment of breast cancer, including triple-negative breast cancer. OncoImmunology 2015, 5, e1078057. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Jurkat | K562 | Caco-2 | T84 | Hs578T | MIA PaCa-2 | Hela | A549 | HEK293 | HCT116 | HNC | MDA-MB231 | Huh7 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAR (%) | 96.4 | 1.1 | 93.5 | 92.6 | 14.1 | 8.9 | 99.7 | 98.9 | 97.2 | 99.5 | 10.3 | 84.8 | 69.5 |
| CD46 (%) | 99.4 | 100 | 98.5 | 94.4 | 99.9 | 99.8 | 99.9 | 99.9 | 93.5 | 99.3 | 99.7 | 99.8 | 99.7 |
| DSG2 (%) | 41.0 | 84.5 | 75.6 | 74.5 | 97.4 | 0.2 | 99.6 | 99.8 | 71.8 | 90.2 | 99.3 | 83.2 | 94.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahlmann, N.A.; Tsoukas, R.L.; Erkens, S.; Wang, H.; Jönsson, F.; Aydin, M.; Naumova, E.A.; Lieber, A.; Ehrhardt, A.; Zhang, W. Properties of Adenovirus Vectors with Increased Affinity to DSG2 and the Potential Benefits of Oncolytic Approaches and Gene Therapy. Viruses 2022, 14, 1835. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081835
Bahlmann NA, Tsoukas RL, Erkens S, Wang H, Jönsson F, Aydin M, Naumova EA, Lieber A, Ehrhardt A, Zhang W. Properties of Adenovirus Vectors with Increased Affinity to DSG2 and the Potential Benefits of Oncolytic Approaches and Gene Therapy. Viruses. 2022; 14(8):1835. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081835
Chicago/Turabian StyleBahlmann, Nora A., Raphael L. Tsoukas, Sebastian Erkens, Hongjie Wang, Franziska Jönsson, Malik Aydin, Ella A. Naumova, André Lieber, Anja Ehrhardt, and Wenli Zhang. 2022. "Properties of Adenovirus Vectors with Increased Affinity to DSG2 and the Potential Benefits of Oncolytic Approaches and Gene Therapy" Viruses 14, no. 8: 1835. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081835