

IFIT3 and IFIT5 Play Potential Roles in Innate Immune Response of Porcine Pulmonary Microvascular Endothelial Cells to Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Isolation and Culture of Porcine Pulmonary MVECs
2.3. Immunofluorescence Staining
2.4. Transmission Electron Microscopy (TEM)
2.5. Quantitative Real-Time PCR (RT-PCR)
2.6. Western Blotting
2.7. Cell Transfection
2.8. Statistical Analysis
3. Results
3.1. Characteristics of Porcine Pulmonary MVECs
3.2. Confirmation of HP-PRRSV Infection in Porcine Pulmonary MVECs
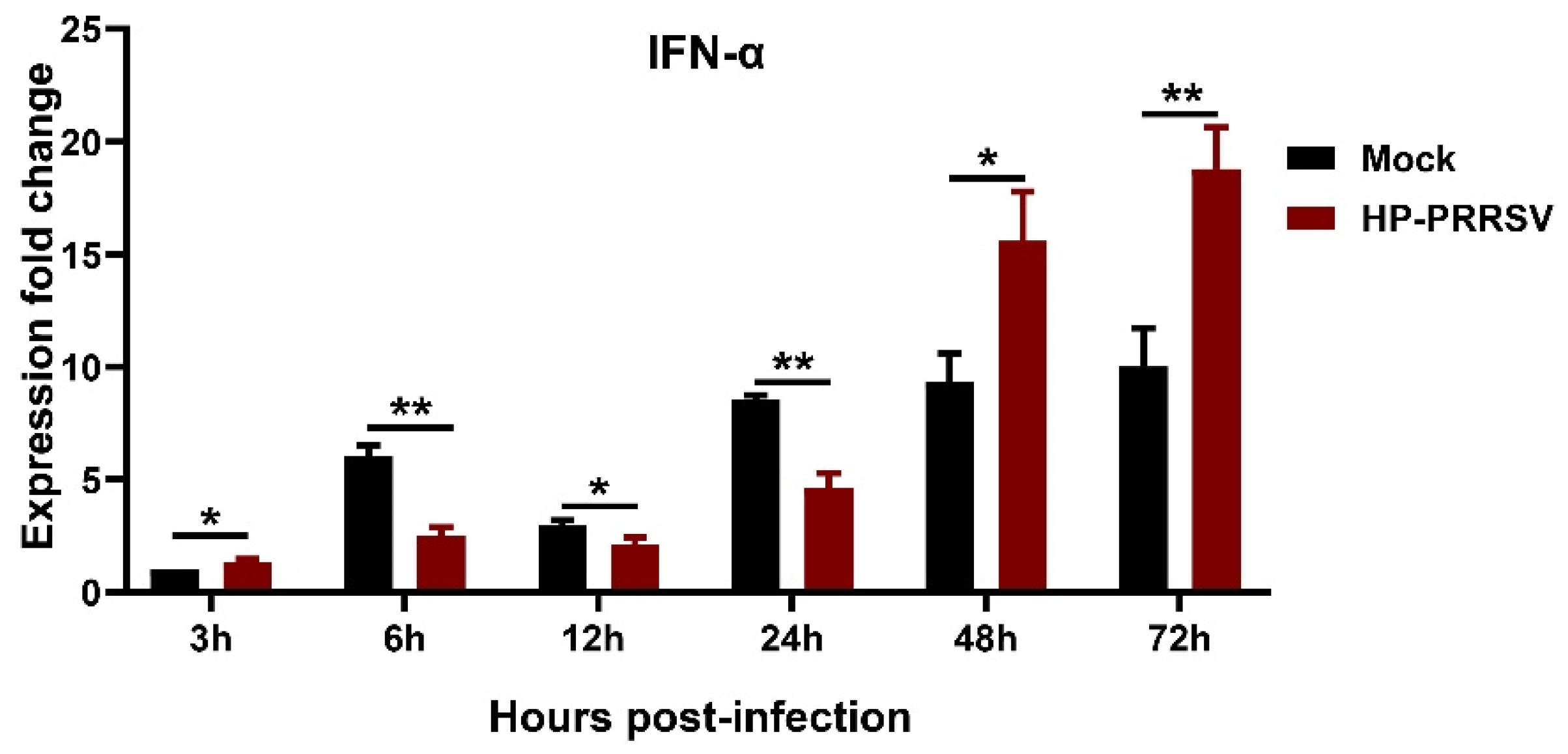
3.3. IFN-α Expression in HP-PRRSV-Infected Porcine Pulmonary MVECs
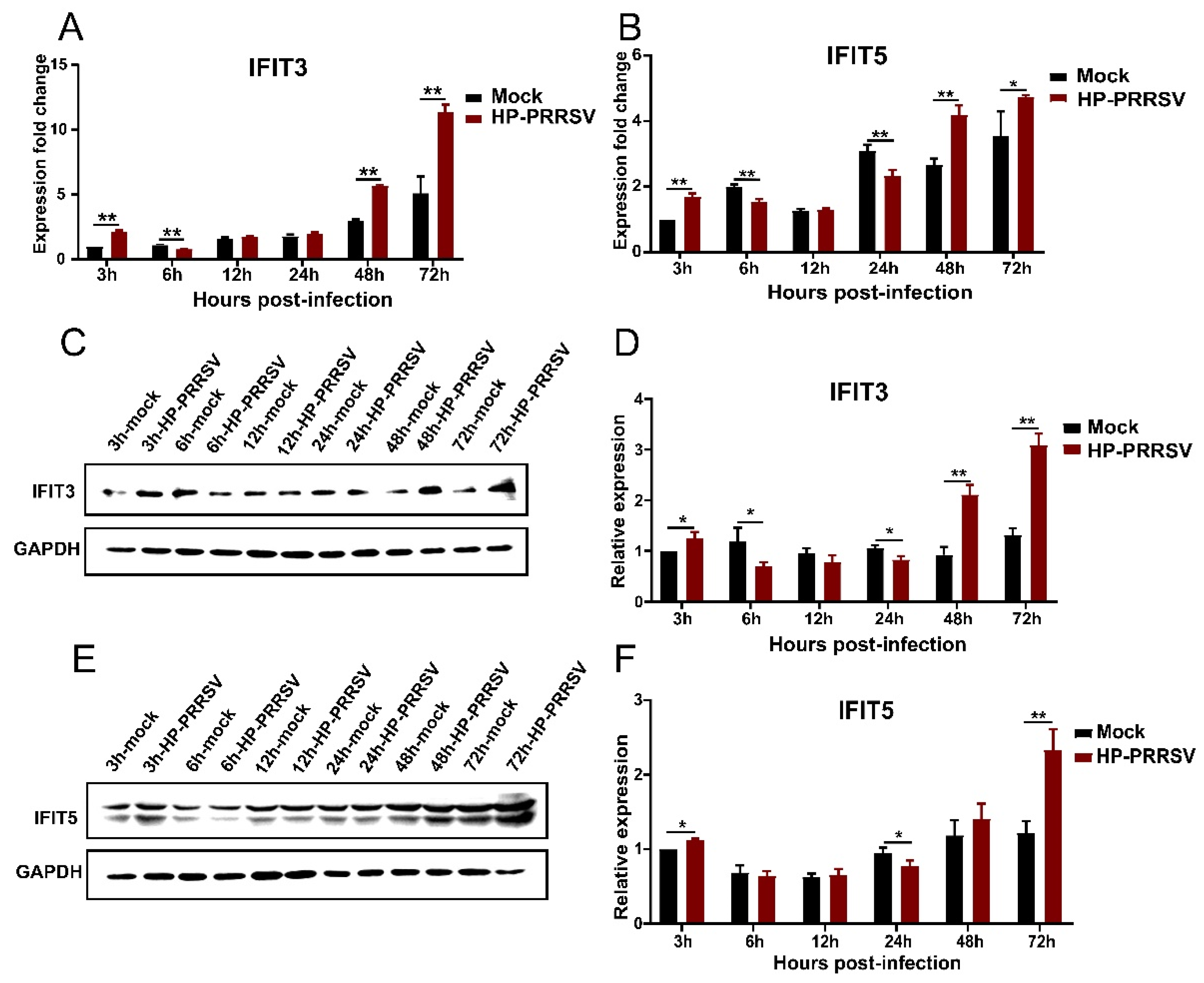
3.4. Expression of IFIT3 and IFIT5 in HP-PRRSV-Infected Porcine Pulmonary MVECs
3.5. Effects of Silencing IFIT3 and IFIT5 Expression on the HP-PRRSV Multiplication
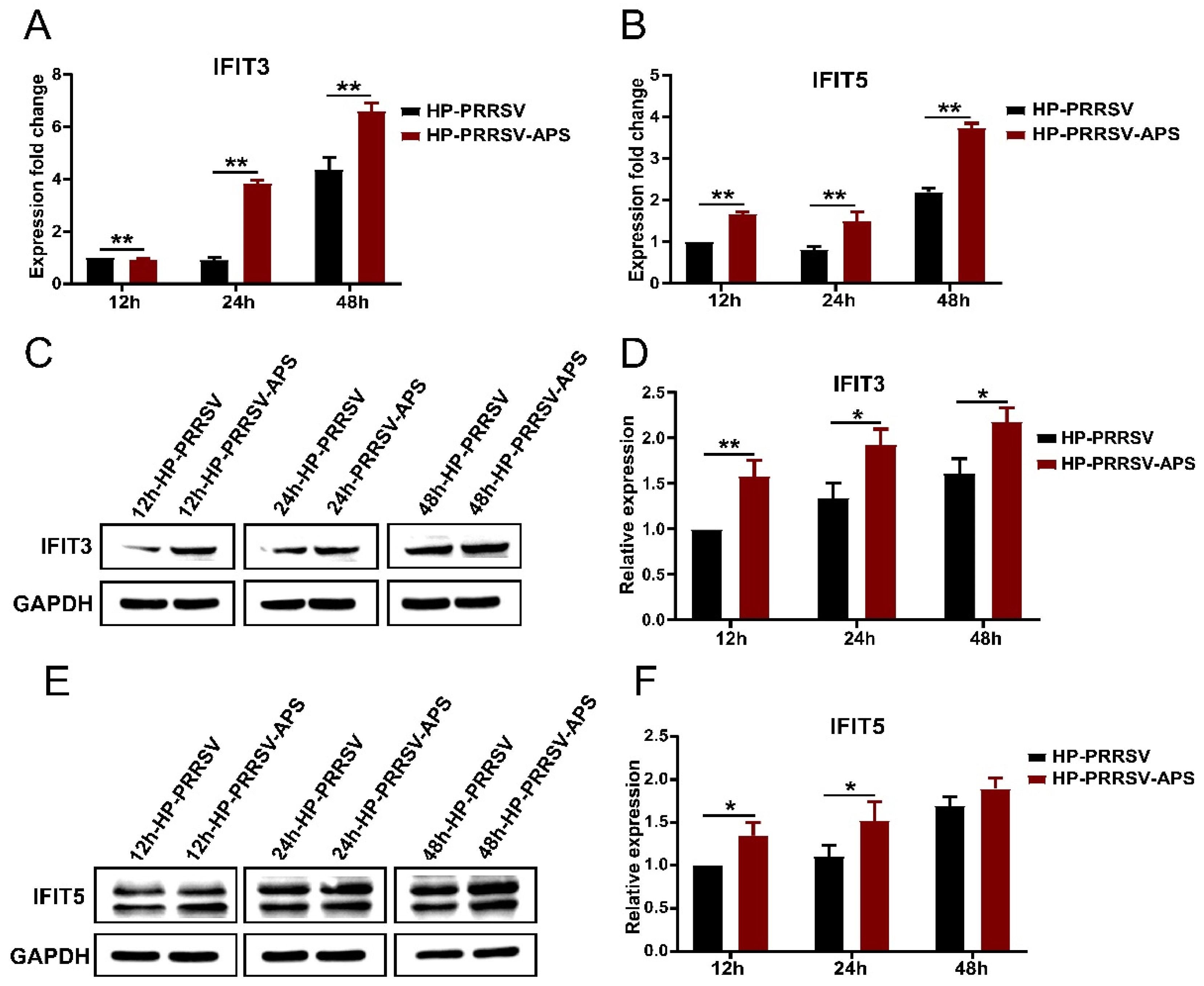
3.6. Effects of APS on the Expression of IFIT3 and IFIT5
3.7. APS Inhibit HP-PRRSV Multiplication in Porcine Pulmonary MVECs
3.8. Gene Silence of IFIT3 and IFIT5 Lowers the Anti-HP-PRRSV Action of APS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Han, J.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. Pathogenesis and control of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2017, 209, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yang, Z.; Ma, S.; Hu, G.; Dong, H.; Zhang, T. Susceptibility of porcine pulmonary microvascular endothelial cells to porcine reproductive and respiratory syndrome virus. J. Vet. Med. Sci. 2020, 82, 1404–1409. [Google Scholar] [CrossRef]
- Shao, Y.; Saredy, J.; Yang, W.Y.; Sun, Y.; Lu, Y.; Saaoud, F.; Drummer, C., IV; Johnson, C.; Xu, K.; Jiang, X.; et al. Vascular endothelial cells and innate immunity. Arterioscler. Thromb. Vasc. Biol. 2020, 40, e138–e152. [Google Scholar] [CrossRef] [PubMed]
- Vladimer, G.I.; Górna, M.W.; Superti-Furga, G. IFITs: Emerging roles as key anti-viral proteins. Front. Immunol. 2014, 5, 94. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Zhang, L.; Cao, L.; Huang, K.; Zhao, Y.; Zhang, Y.; Chen, X.; Lin, X.; Chen, M.; Jin, M. SARS-CoV-2 promote autophagy to suppress type I interferon response. Signal Transduct. Target. Ther. 2021, 6, 180. [Google Scholar] [CrossRef]
- Zhu, H.; Cong, J.P.; Shenk, T. Use of differential display analysis to assess the effect of human cytomegalovirus infection on the accumulation of cellular RNAs: Induction of interferon-responsive RNAs. Proc. Natl. Acad. Sci. USA 1997, 94, 13985–13990. [Google Scholar] [CrossRef]
- Zhao, H.; Boije, H.; Granberg, F.; Pettersson, U.; Svensson, C. Activation of the interferon-induced STAT pathway during an adenovirus type 12 infection. Virology 2009, 392, 186–195. [Google Scholar] [CrossRef]
- Zhou, X.; Michal, J.J.; Zhang, L.; Ding, B.; Lunney, J.K.; Liu, B.; Jiang, Z. Interferon induced IFIT family genes in host antiviral defense. Int. J. Biol. Sci. 2013, 9, 200–208. [Google Scholar] [CrossRef]
- Feng, B.; Zhang, Q.; Zhang, T.; Wang, J.; Dong, H.; Mu, X.; Hu, G. IFIT1 Expression Patterns Induced by H9N2 Virus and Inactivated Viral Particle in Human Umbilical Vein Endothelial Cells and Bronchus Epithelial Cells. Mol. Cells 2018, 41, 271–281. [Google Scholar]
- Zhang, L.; Liu, J.; Bai, J.; Du, Y.; Wang, X.; Liu, X.; Jiang, P. Poly (I:C) inhibits porcine reproductive and respiratory syndrome virus replication in MARC-145 cells via activation of IFIT3. Antivir. Res. 2013, 99, 197–206. [Google Scholar] [CrossRef]
- Ma, G. RNA-Seq Analysis and Determination of Restriction Factors against Highly Pathogenic Porcine Reproductive and Respiratory Syndrome. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2016. [Google Scholar]
- Kawakita, T.; Nakai, S.; Kumazawa, Y.; Miura, O.; Yumioka, E.; Nomoto, K. Induction of interferon after administration of a traditional Chinese medicine, xiao-chai-hu-tang (shosaiko-to). Int. J. Immunopharmacol. 1990, 12, 515–521. [Google Scholar] [CrossRef]
- Yin, Y.; Lu, L.; Wang, D.; Shi, Y.; Wang, M.; Huang, Y.; Chen, D.; Deng, C.; Chen, J.; Lv, P.; et al. Astragalus Polysaccharide Inhibits Autophagy and Apoptosis from Peroxide-Induced Injury in C2C12 Myoblasts. Cell Biochem. Biophys. 2015, 73, 433–439. [Google Scholar] [CrossRef]
- Zheng, Y.; Ren, W.; Zhang, L.; Zhang, Y.; Liu, D.; Liu, Y. A review of the pharmacological action of Astragalus polysaccharide. Front. Pharmacol. 2020, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Meng, G.; Teng, F.; Sun, Q.; Zhang, Y. Retracted: Effects of astragalus polysaccharide on apoptosis of myocardial microvascular endothelial cells in rats undergoing hypoxia/reoxygenation by mediation of the PI3K/Akt/eNOS signaling pathway. J. Cell. Biochem. 2018, 119, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Kang, H.; Tuo, T.; Wang, L.; Xia, Y.; Zhang, Y.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; et al. Astragalus polysaccharide alleviated the inhibition of CSFV C-strain replication caused by PRRSV via the TLRs/NF-κB/TNF-α pathways. Virus Res. 2022, 319, 198854. [Google Scholar] [CrossRef]
- Zhuge, Z.Y.; Zhu, Y.H.; Liu, P.Q.; Yan, X.D.; Yue, Y.; Weng, X.G.; Zhang, R.; Wang, J.F. Effects of Astragalus polysaccharide on immune responses of porcine PBMC stimulated with PRRSV or CSFV. PLoS ONE 2012, 7, e29320. [Google Scholar]
- Wu, Y.; Wang, Z.; Hu, G.; Zhang, T. Isolation and culture of rat Intestinal mucosal microvascular endothelial cells using immunomagnetic beads and their marker expression. J. Immunol. Methods 2022, 507, 113296. [Google Scholar] [CrossRef]
- Nellinger, S.; Schmidt, I.; Heine, S.; Volz, A.-C.; Kluger, P.J. Adipose stem cell-derived extracellular matrix represents a promising biomaterial by inducing spontaneous formation of prevascular-like structures by mvECs. Biotechnol. Bioeng. 2020, 117, 3160–3172. [Google Scholar] [CrossRef]
- Du, T.; Nan, Y.; Xiao, S.; Zhao, Q.; Zhou, E.-M. Antiviral strategies against PRRSV infection. Trends Microbiol. 2017, 25, 968–979. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.; Pan, Y.; Gao, J.; Xu, Y.; Li, X.; Tian, Z.; Chen, H.; Wang, Y. Downregulation of miR-218 by porcine reproductive and respiratory syndrome virus facilitates viral replication via inhibition of type I interferon responses. J. Biol. Chem. 2021, 296, 100683. [Google Scholar] [CrossRef]
- Sun, Y.; Han, M.; Kim, C.; Calvert, J.G.; Yoo, D. Interplay between interferon-mediated innate immunity and porcine reproductive and respiratory syndrome virus. Viruses 2012, 4, 424–446. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Farzan, M. The broad-spectrum antiviral functions of IFIT and IFITM proteins. Nat. Rev. Immunol. 2013, 13, 46–57. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Li, J.N.; Su, C.M.; Yoo, D. Molecular and cellular mechanisms for PRRSV pathogenesis and host response to infection. Virus Res. 2020, 286, 197980. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Chen, W.; Wei, B.; Shan, Y.-F.; Wang, C. IFN-induced TPR protein IFIT3 potentiates antiviral signaling by bridging MAVS and TBK1. J. Immunol. 2011, 187, 2559–2568. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, X.; Liu, H.; Wang, W.; Liu, X.; Li, X.; Wu, X. Astragalus polysaccharides inhibit avian infectious bronchitis virus infection by regulating viral replication. Microb. Pathog. 2018, 114, 124–128. [Google Scholar] [CrossRef]
- Sun, W.; Wu, W.; Jiang, N.; Ge, X.; Zhang, Y.; Han, J.; Guo, X.; Zhou, L.; Yang, H. Highly Pathogenic PRRSV-Infected Alveolar Macrophages Impair the Function of Pulmonary Microvascular Endothelial Cells. Viruses 2022, 14, 452. [Google Scholar] [CrossRef]
- Dang, S.S.; Jia, X.L.; Song, P.; Cheng, Y.-A.; Zhang, X.; Sun, M.-Z.; Liu, E.-Q. Inhibitory effect of emodin and Astragalus polysaccharide on the replication of HBV. World J. Gastroenterol. 2009, 15, 5669. [Google Scholar] [CrossRef]
- Li, W.; Hu, X.; Wang, S.; Jiao, Z.; Sun, T.; Liu, T.; Song, K. Characterization and anti-tumor bioactivity of astragalus polysaccharides by immunomodulation. Int. J. Biol. Macromol. 2020, 145, 985–997. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, W.; Zhang, J.; Song, X.; Sun, W.; Fan, Y. The immunoregulatory activities of astragalus polysaccharide liposome on macrophages and dendritic cells. Int. J. Biol. Macromol. 2017, 105, 852–861. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Song, X.; Cui, D.; Zhang, T. IFIT3 and IFIT5 Play Potential Roles in Innate Immune Response of Porcine Pulmonary Microvascular Endothelial Cells to Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus. Viruses 2022, 14, 1919. https://0-doi-org.brum.beds.ac.uk/10.3390/v14091919
Wu Y, Song X, Cui D, Zhang T. IFIT3 and IFIT5 Play Potential Roles in Innate Immune Response of Porcine Pulmonary Microvascular Endothelial Cells to Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus. Viruses. 2022; 14(9):1919. https://0-doi-org.brum.beds.ac.uk/10.3390/v14091919
Chicago/Turabian StyleWu, Yanmei, Xiaoxiao Song, Defeng Cui, and Tao Zhang. 2022. "IFIT3 and IFIT5 Play Potential Roles in Innate Immune Response of Porcine Pulmonary Microvascular Endothelial Cells to Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus" Viruses 14, no. 9: 1919. https://0-doi-org.brum.beds.ac.uk/10.3390/v14091919