

Epizootic Haemorrhagic Disease Virus Serotype 8 in Tunisia, 2021

 , , , , , , , , ,
, , , , , , , , ,  and add
Show full author list
and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Ethical Statement
2.2. Specimen Collection
2.3. Serological Tests for EHDV
2.4. EHDV Detection by Real Time RT-PCR and Genotyping
2.5. Virus Isolation and Virus Neutralization Test (VNT)
2.6. Shotgun (SG) Metagenomic Analysis by Oxford Nanopore MinION
2.7. Illumina Genome Sequencing
2.8. Phylogenetic Analysis
3. Results
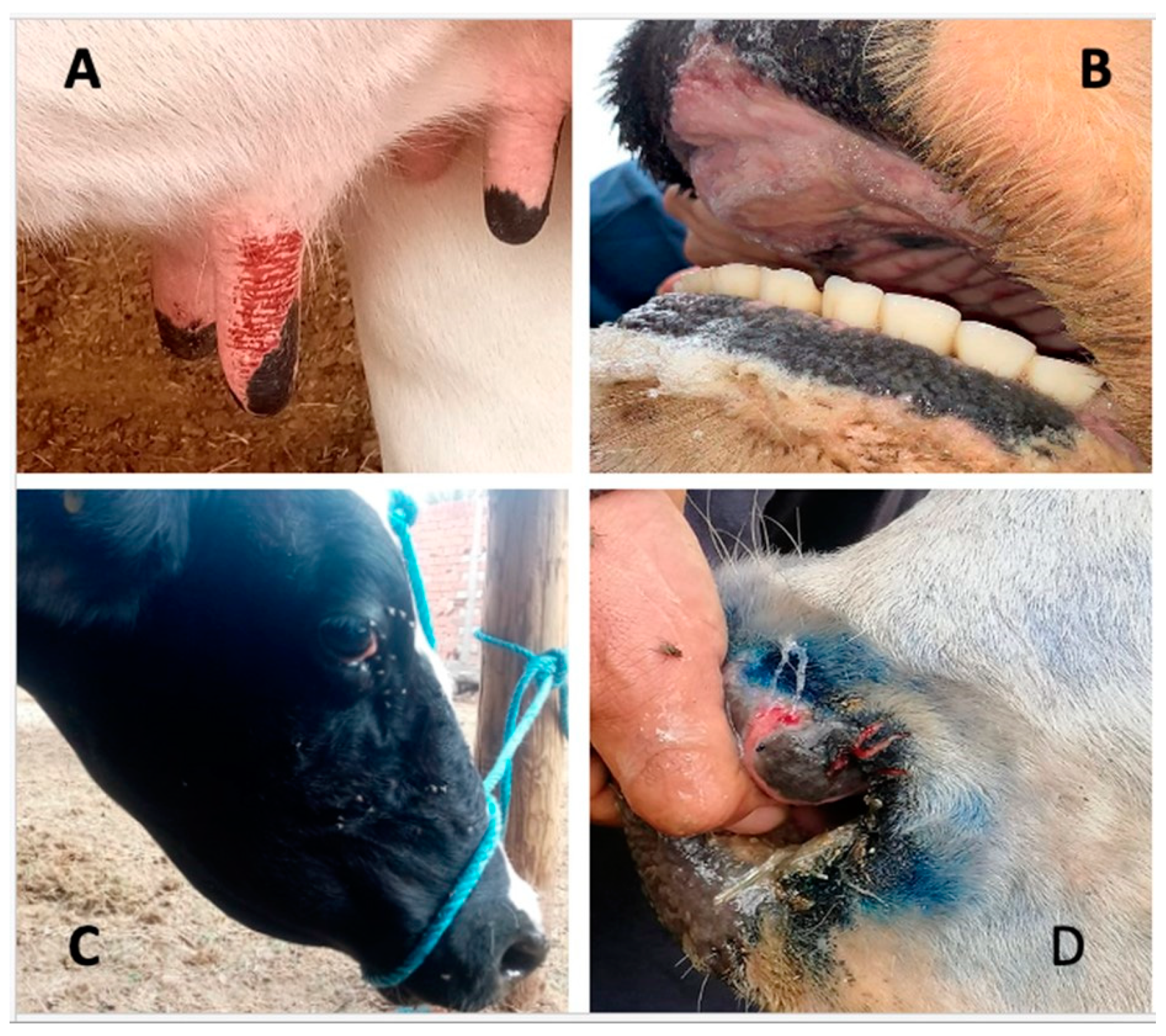
3.1. Serological and Virological Tests Results
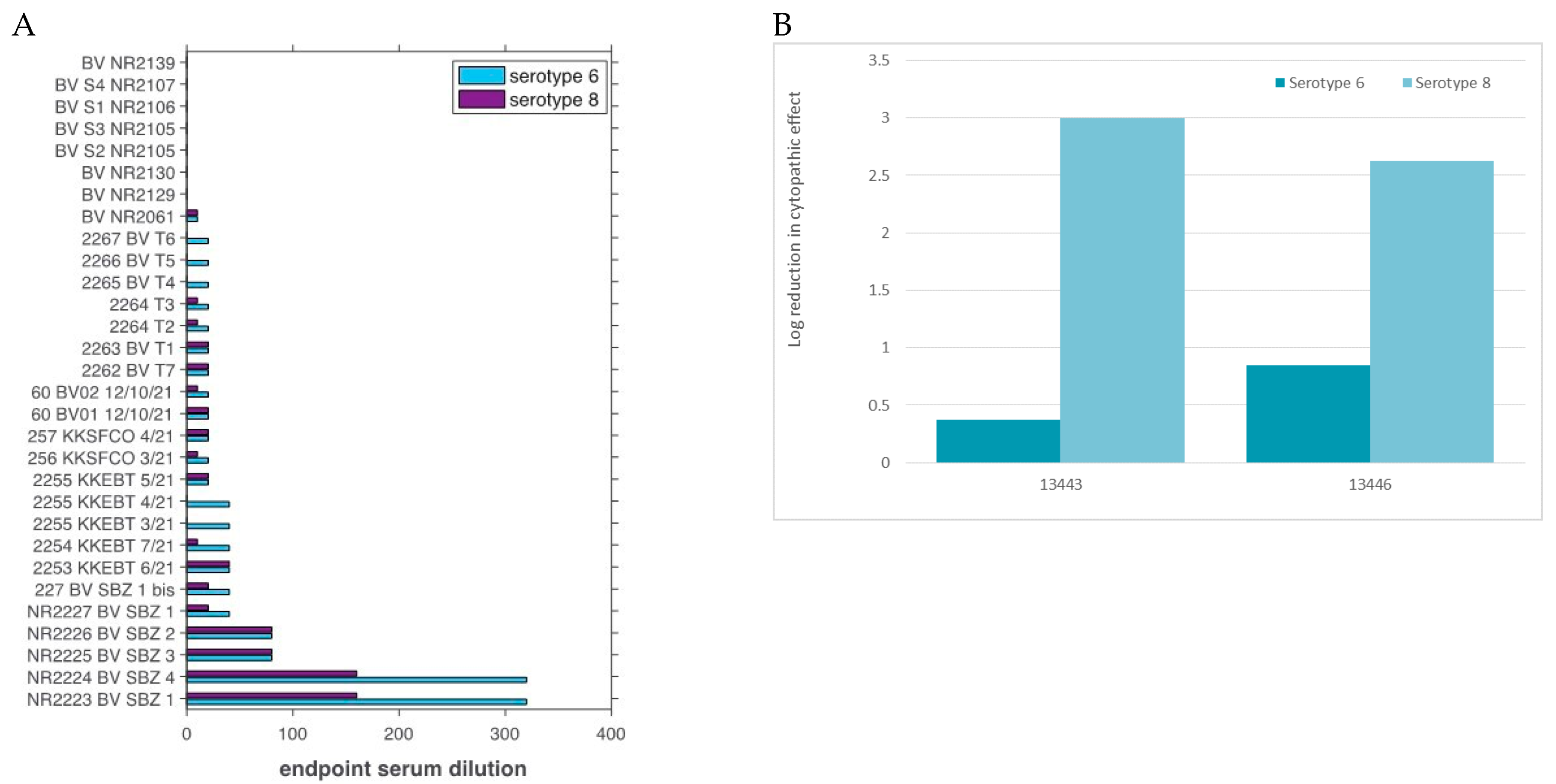
3.2. Virus Isolation and VNT
3.3. EHDV Genome Identified by Shotgun Metagenomic Sequencing
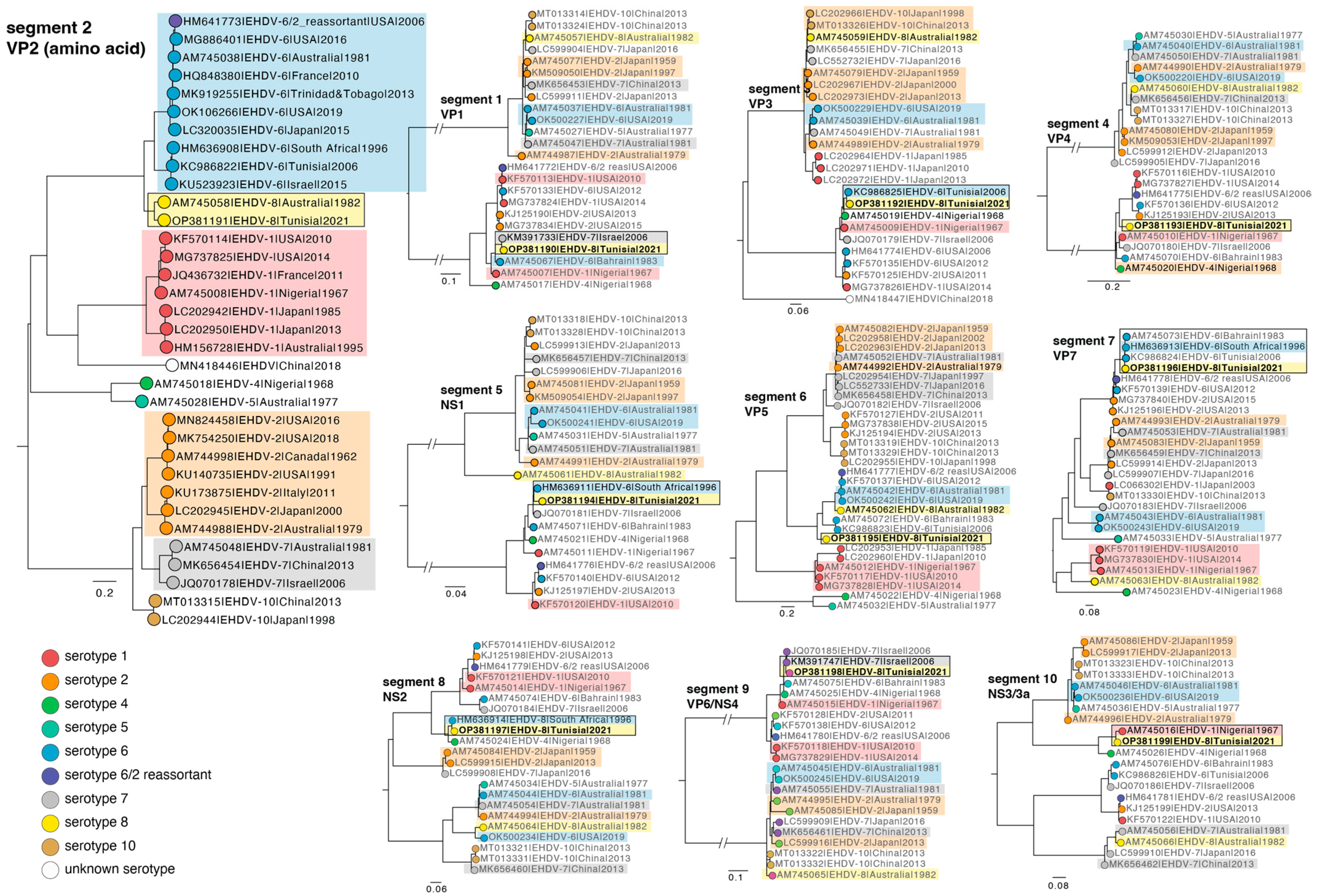
3.4. The Whole Genome Sequence of EHDV-8/17 TUN 2021 Strain
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- MacLachlan, N.J. Bluetongue: Pathogenesis and duration of viraemia. Vet. Ital. 2004, 40, 462–467. [Google Scholar] [PubMed]
- Maclachlan, N.J.; Mayo, C.E.; Daniels, P.W.; Savini, G.; Zientara, S.; Gibbs, E.P. Bluetongue. Rev. Sci. Tech. 2015, 34, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, E.P.; Lawman, M.J. Infection of British deer and farm animals with epizootic haemorrhagic disease of deer virus. J. Comp. Pathol. 1977, 87, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Afonso, A.; Mellor, P.; Aradaib, I.; Yadin, H.; Sanaa, M.; Wilson, W.; Monaco, F.; Domingo, M. Epizootic heamorragic disease. Res. Vet. Sci. 2011, 91, 1–17. [Google Scholar] [CrossRef]
- Breard, E.; Sailleau, C.; Hamblin, C.; Graham, S.D.; Gourreau, J.M.; Zientara, S. Outbreak of epizootic haemorrhagic disease on the island of La Réunion. Vet. Rec. 2004, 155, 422–423. [Google Scholar] [CrossRef] [PubMed]
- Belhouchet, M.; Mohd Jaafar, F.; Firth, A.E.; Grimes, J.M.; Mertens, P.P.; Attoui, H. Detection of a fourth orbivirus non-structural protein. PLoS ONE 2011, 6, e25697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, S.J.; Maan, S.; Maan, N.; Kgosana, L.; Bachanek-Bankowska, K.; Batten, C.; Darpel, K.E.; Sutton, G.; Attoui, H.; Mertens, P.P. Genetic and phylogenetic analysis of the outer-coat proteins VP2 and VP5 of epizootic haemorrhagic disease virus (EHDV): Comparison of genetic and serological data to characterise the EHDV serogroup. Virus Res. 2009, 145, 200–210. [Google Scholar] [CrossRef]
- Uchinuno, Y.; Ito, T.; Goto, Y.; Miura, Y.; Ishibashi, K.; Itou, T.; Sakai, T. Differences in Ibaraki virus RNA segment 3 sequences from three epidemics. J. Vet. Med. Sci. 2003, 65, 1257–1263. [Google Scholar] [CrossRef] [Green Version]
- Shirafuji, H.; Kato, T.; Yamakawa, M.; Tanaka, T.; Minemori, Y.; Yanase, T. Characterization of genome segments 2, 3 and 6 of epizootic hemorrhagic disease virus strains isolated in Japan in 1985-2013: Identification of their serotypes and geographical genetic types. Infect. Genet. Evol. 2017, 53, 38–46. [Google Scholar] [CrossRef]
- Yang, H.; Li, Z.; Wang, J.; Li, Z.; Yang, Z.; Liao, D.; Zhu, J.; Li, H. Novel Serotype of Epizootic Hemorrhagic Disease Virus, China. Emerg. Infect. Dis. 2020, 26, 3081–3083. [Google Scholar] [CrossRef]
- McVey, D.S.; MacLachlan, N.J. Vaccines for Prevention of Bluetongue and Epizootic Hemorrhagic Disease in Livestock: A North American Perspective. Vector Borne Zoonotic Dis. 2015, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Verdezoto, J.; Breard, E.; Viarouge, C.; Quenault, H.; Lucas, P.; Sailleau, C.; Zientara, S.; Augot, D.; Zapata, S. Novel serotype of bluetongue virus in South America and first report of epizootic haemorrhagic disease virus in Ecuador. Transbound. Emerg. Dis. 2018, 65, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Dommergues, L.; Viarouge, C.; Métras, R.; Youssouffi, C.; Sailleau, C.; Zientara, S.; Cardinale, E.; Cêtre-Sossah, C. Evidence of bluetongue and Epizootic Haemorrhagic disease circulation on the island of Mayotte. Acta Trop. 2019, 191, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Viarouge, C.; Lancelot, R.; Rives, G.; Bréard, E.; Miller, M.; Baudrimont, X.; Doceul, V.; Vitour, D.; Zientara, S.; Sailleau, C. Identification of bluetongue virus and epizootic hemorrhagic disease virus serotypes in French Guiana in 2011 and 2012. Vet. Microbiol. 2014, 174, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Mahmoud, M.A.E.; Viarouge, C.; Sailleau, C.; Zientara, S.; Breard, E. Presence of bluetongue and epizootic hemorrhagic disease viruses in Egypt in 2016 and 2017. Infect. Genet. Evol. 2019, 73, 221–226. [Google Scholar] [CrossRef]
- Golender, N.; Bumbarov, V.Y. Detection of Epizootic Hemorrhagic Disease Virus Serotype 1, Israel. Emerg. Infect. Dis. 2019, 25, 825–827. [Google Scholar] [CrossRef]
- Golender, N.; Khinich, Y.; Gorohov, A.; Abramovitz, I.; Bumbarov, V. Epizootic hemorrhagic disease virus serotype 6 outbreak in Israeli cattle in 2015. J. Vet. Diagn. Invest. 2017, 29, 885–888. [Google Scholar] [CrossRef]
- Komarov, A.; Goldsmit, L. A disease, similar to bluetongue in cattle and sheep in Israel. Ref. Vet. 1951, 8, 96–100. [Google Scholar]
- Ben Dhaou, S.; Sailleau, C.; Babay, B.; Viarouge, C.; Sghaier, S.; Zientara, S.; Hammami, S.; Bréard, E. Molecular characterisation of epizootic haemorrhagic disease virus associated with a Tunisian outbreak among cattle in 2006. Acta Vet. Hung. 2016, 64, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.; Danzetta, M.L.; di Sabatino, D.; Spedicato, M.; Alkhatal, Z.; Dayhum, A.; Tolari, F.; Forzan, M.; Mazzei, M.; Savini, G. First seroprevalence investigation of epizootic haemorrhagic disease virus in Libya. Open Vet. J. 2021, 11, 301–308. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, F.; Chang, J.; Zhang, Y.; Zhu, J.; Li, H.; Yu, L. Identification and complete-genome phylogenetic analysis of an epizootic hemorrhagic disease virus serotype 7 strain isolated in China. Arch. Virol. 2019, 164, 3121–3126. [Google Scholar] [CrossRef] [PubMed]
- Maan, N.S.; Maan, S.; Potgieter, A.C.; Wright, I.M.; Belaganahalli, M.; Mertens, P.P.C. Development of Real-Time RT-PCR Assays for Detection and Typing of Epizootic Haemorrhagic Disease Virus. Transbound. Emerg. Dis. 2017, 64, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Viarouge, C.; Breard, E.; Zientara, S.; Vitour, D.; Sailleau, C. Duplex Real-Time RT-PCR Assays for the Detection and Typing of Epizootic Haemorrhagic Disease Virus. PLoS ONE 2015, 10, e0132540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, C.; Vögtlin, A.; Hüssy, D.; Jandt, T.; Gobet, H.; Hilbe, M.; Burgener, C.; Schweizer, L.; Häfliger-Speiser, S.; Beer, M.; et al. Putative Novel Atypical BTV Serotype ‘36’ Identified in Small Ruminants in Switzerland. Viruses 2021, 13, 721. [Google Scholar] [CrossRef]
- Sailleau, C.; Breard, E.; Viarouge, C.; Belbis, G.; Lilin, T.; Vitour, D.; Zientara, S. Experimental infection of calves with seven serotypes of Epizootic Hemorrhagic Disease virus: Production and characterization of reference sera. Vet. Ital. 2019, 55, 339–346. [Google Scholar] [CrossRef]
- Marcacci, M.; De Luca, E.; Zaccaria, G.; Di Tommaso, M.; Mangone, I.; Aste, G.; Savini, G.; Boari, A.; Lorusso, A. Genome characterization of feline morbillivirus from Italy. J. Virol. Methods 2016, 234, 160–163. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Uribasterra, M.G.; Orange, J.P.; Dinh, E.T.N.; Peters, C.; Peters, R.M.; Goodfriend, O.; Wisely, S.M.; Blackburn, J.K. Epizootic Hemorrhagic Disease Virus and Bluetongue Virus Seroprevalence in Wild White-Tailed Deer (Odocoileus virginianus) in Florida, USA. J. Wildl. Dis. 2020, 56, 928–932. [Google Scholar] [CrossRef]
- Lorusso, A.; Sghaier, S.; Carvelli, A.; Di Gennaro, A.; Leone, A.; Marini, V.; Pelini, S.; Marcacci, M.; Rocchigiani, A.M.; Puggioni, G.; et al. Bluetongue virus serotypes 1 and 4 in Sardinia during autumn 2012: New incursions or re-infection with old strains? Infect. Genet. Evol. 2013, 19, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.; Sghaier, S.; Ancora, M.; Marcacci, M.; Di Gennaro, A.; Portanti, O.; Mangone, I.; Teodori, L.; Leone, A.; Camma, C.; et al. Molecular epidemiology of bluetongue virus serotype 1 circulating in Italy and its connection with northern Africa. Infect. Genet. Evol. 2014, 28, 144–149. [Google Scholar] [CrossRef]
- Sghaier, S.; Lorusso, A.; Portanti, O.; Marcacci, M.; Orsini, M.; Barbria, M.E.; Mahmoud, A.S.; Hammami, S.; Petrini, A.; Savini, G. A novel Bluetongue virus serotype 3 strain in Tunisia, November 2016. Transbound. Emerg. Dis. 2017, 64, 709–715. [Google Scholar] [CrossRef]
- Cappai, S.; Rolesu, S.; Loi, F.; Liciardi, M.; Leone, A.; Marcacci, M.; Teodori, L.; Mangone, I.; Sghaier, S.; Portanti, O.; et al. Western Bluetongue virus serotype 3 in Sardinia, diagnosis and characterization. Transbound. Emerg. Dis. 2019, 66, 1426–1431. [Google Scholar] [CrossRef] [Green Version]
- Serroni, A.; Ulisse, S.; Iorio, M.; Laguardia, C.; Testa, L.; Armillotta, G.; Caporale, M.; Salini, R.; Lelli, D.; Wernery, U.; et al. Development of a Competitive Enzyme-Linked Immunosorbent Assay Based on Purified Recombinant Viral Protein 7 for Serological Diagnosis of Epizootic Haemorrhagic Disease in Camels. J. Trop. Med. 2022, 2022, 5210771. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Accession Nr | Segment | Blastn Output Strain (Accession Number) | % Identity |
|---|---|---|---|
| OP381190 | 1 | Israel 2006 ISR2006/04 serotype 7 (KM391733) | 98.58 |
| OP381191 | 2 | Australia 1982 CPR_3961A serotype 8 (AM745058) | 76.53 |
| OP381192 | 3 | Tunisia 2006 2577 serotype 6 (KC986825) | 97.41 |
| OP381193 | 4 | Nigeria 1968 IbAr33583 serotype 4 (AM745020) | 93.29 |
| OP381194 | 5 | South Africa 1996 M44/96 serotype 6 (HM636911) | 97.55 |
| OP381195 | 6 | Australia 1982 CPR_3961A serotype 8 (AM745062) | 80.52 |
| OP381196 | 7 | South Africa 1996 M44/96 serotype 6 (HM636913) | 98.01 |
| OP381197 | 8 | South Africa 1996 M44/96 serotype 6 (HM636914) | 98.31 |
| OP381198 | 9 | Israel 2006 ISR2006/04 serotype 7 (KM391738) | 97.63 |
| OP381199 | 10 | Nigeria 1967 IbAr22619 serotype 1 (AM745016) | 95.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sghaier, S.; Sailleau, C.; Marcacci, M.; Thabet, S.; Curini, V.; Ben Hassine, T.; Teodori, L.; Portanti, O.; Hammami, S.; Jurisic, L.; et al. Epizootic Haemorrhagic Disease Virus Serotype 8 in Tunisia, 2021. Viruses 2023, 15, 16. https://0-doi-org.brum.beds.ac.uk/10.3390/v15010016
Sghaier S, Sailleau C, Marcacci M, Thabet S, Curini V, Ben Hassine T, Teodori L, Portanti O, Hammami S, Jurisic L, et al. Epizootic Haemorrhagic Disease Virus Serotype 8 in Tunisia, 2021. Viruses. 2023; 15(1):16. https://0-doi-org.brum.beds.ac.uk/10.3390/v15010016
Chicago/Turabian StyleSghaier, Soufien, Corinne Sailleau, Maurilia Marcacci, Sarah Thabet, Valentina Curini, Thameur Ben Hassine, Liana Teodori, Ottavio Portanti, Salah Hammami, Lucija Jurisic, and et al. 2023. "Epizootic Haemorrhagic Disease Virus Serotype 8 in Tunisia, 2021" Viruses 15, no. 1: 16. https://0-doi-org.brum.beds.ac.uk/10.3390/v15010016