
Small Heat Shock Protein (sHsp22.98) from Trialeurodes vaporariorum Plays Important Role in Apple Scar Skin Viroid Transmission
 , , and
, , and
Abstract
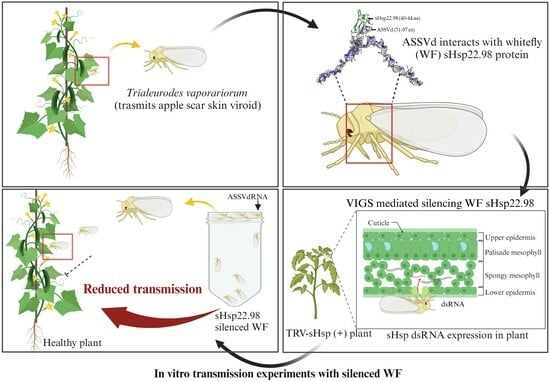
:
1. Introduction
2. Materials and Methods
2.1. Maintenance of Whitefly Colony and Pathogen
2.2. Characterization of sHsp Gene
2.3. Heterologous Expression and Purification of Small Heat Shock Protein
2.4. Western Blotting
2.5. In Vitro Binding Assay
2.6. Bioinformatic Analysis of ASSVd Association with Small Heat Shock Protein
2.7. Transient Silencing of Small Heat Shock Protein in T. vaporariorum
2.8. Transmission of ASSVd by T. vaporariorum
3. Results
3.1. Molecular Characterization of T. vaporariorum Small Heat Shock Protein (sHsp22.98)
3.2. Recombinant Expression of sHsp22.98
3.3. In Vitro Binding of Recombinant sHsp22.98 to ASSVd RNA
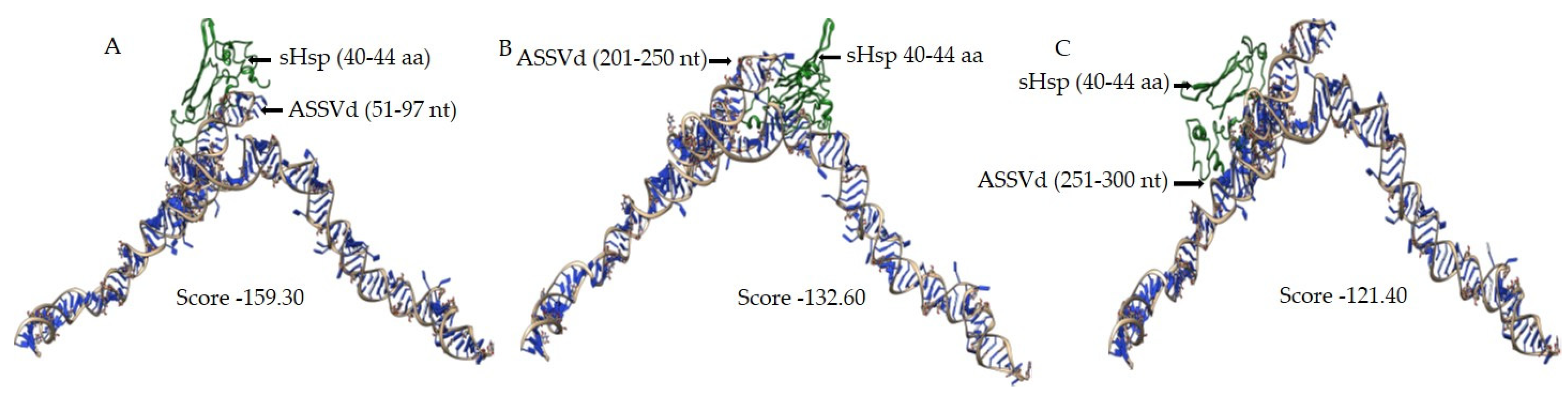
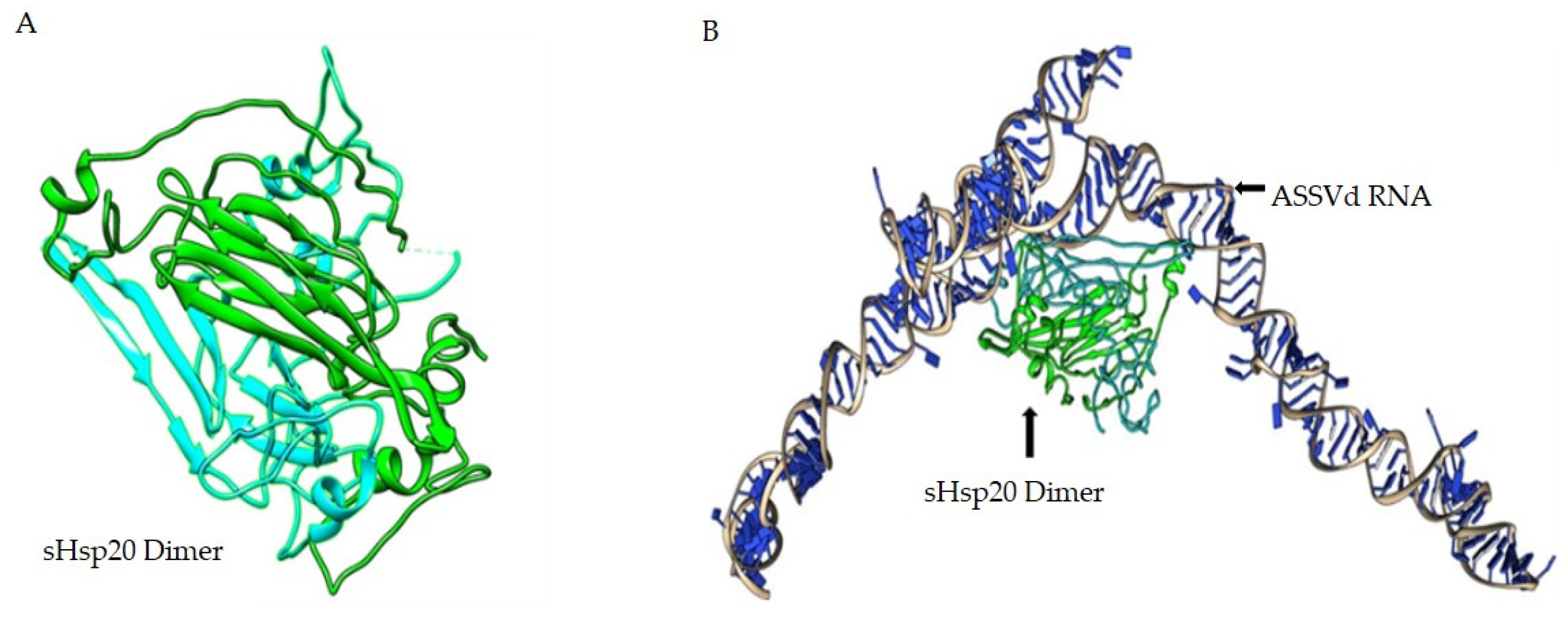
3.4. In Silico Analysis of sHsp22.98 Dynamic Association with ASSVd
3.5. Plant-Mediated Silencing of sHsp22.98 in T. vaporariorum
3.6. Transmission of ASSVd through sHsp22.98-Silenced Whiteflies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Umer, M.; Liu, J.; You, H.; Xu, C.; Dong, K.; Luo, N.; Kong, L.; Li, X.; Hong, N.; Wang, G.; et al. Genomic, morphological and biological traits of the viruses infecting major fruit trees. Viruses 2019, 11, 515. [Google Scholar] [CrossRef] [PubMed]
- Bettoni, J.C.; Fazio, G.; Carvalho Costa, L.; Hurtado-Gonzales, O.P.; Rwahnih, M.A.; Nedrow, A.; Volk, G.M. Thermotherapy followed by shoot tip cryotherapy eradicates latent viruses and apple hammerhead viroid from in vitro apple rootstocks. Plants 2022, 11, 582. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, R.M.; Ram, R.; Badyal, J.; Hallan, V.; Zaidi, A.A.; Varma, A. Determination of major viral and sub viral pathogens incidence in apple orchards in Himachal Pradesh. Indian J. Virol. 2012, 23, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Randles, J.W.; Bar-Joseph, M.; Diener, T.O. A proposed scheme for viroid classification and nomenclature. Arch. Virol. 1998, 143, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, J.; Koganezawa, H. Nucleotide sequence and secondary structure of apple scar skin viroid. Nucleic Acids Res. 1987, 15, 7045–7052. [Google Scholar] [CrossRef]
- Sipahioglu, H.M.; Usta, M.; Ocak, M. Development of a rapid enzymatic cDNA amplification test for the detection of apple scar skin viroid (ASSVd) in apple trees from eastern Anatolia, Turkey. Arch. Phytopathol. Plant Prot. 2009, 42, 352–360. [Google Scholar] [CrossRef]
- Puchta, H.; Luckinger, R.; Yang, X.; Hadidi, A.; Sänger, H.L. Nucleotide sequence and secondary structure of apple scar skin viroid (ASSVd) from China. Plant Mol. Biol. 1990, 14, 1065–1067. [Google Scholar] [CrossRef]
- Chen, W. Pear rusty skin, a viroid disease. In Proceedings of the 7th International Congress of Virology, Edmonton, AB, Canada; 1987; p. 300. [Google Scholar]
- Osaki, H.; Kudo, A.; Ohtsu, Y. Japanese pear fruit dimple disease caused by apple scar skin viroid (ASSVd). Jpn. J. Phytopathol 1996, 62, 379–385. [Google Scholar] [CrossRef]
- Hadidi, A.; Flores, R.; Randles, J.W.; Palukaitis, P. Viroids and Satellites, 1st ed.; Academic Press: Oxford, UK, 2017; p. 59. [Google Scholar]
- Li, G.; Li, J.; Zhang, H.; Li, J.; Jia, L.; Zhou, S.; Wang, Y.; Sun, J.; Tan, M.; Shao, J. ASSVd infection inhibits the vegetative growth of apple trees by affecting leaf metabolism. Front. Plant Sci. 2023, 14, 1137630. [Google Scholar] [CrossRef]
- Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Manihot esculenta (Cassava). In Encyclopedia of Plant Viruses and Viroids; Springer: New Delhi, India, 2019. [Google Scholar] [CrossRef]
- Kaponi, M.S.; Luigi, M.; Barba, M.; Kyriakopoulou, P.E. Molecular characterization of Hellenic variants of Apple scar skin viroid and Pear blister canker viroid in pome fruit trees. In Proceedings of the 21st International Conference on Virus and Other Graft Transmissible Diseases of Fruit Crops, Neustadt, Germany, 5 July 2009. [Google Scholar]
- Walia, Y.; Dhir, S.; Bhadoria, S.; Hallan, V.; Zaidi, A.A. Molecular characterization of apple scar skin viroid from Himalayan wild cherry. For. Pathol. 2012, 42, 84–87. [Google Scholar] [CrossRef]
- Walia, Y.; Dhir, S.; Ram, R.; Zaidi, A.A.; Hallan, V. Identification of the herbaceous host range of Apple scar skin viroid and analysis of its progeny variants. Plant Pathol. 2014, 63, 684–690. [Google Scholar] [CrossRef]
- Tian, M.; Wei, S.; Bian, R.; Luo, J.; Khan, H.A.; Tai, H.; Kondo, H.; Hadidi, A.; Andika, I.B.; Sun, L. Natural cross-kingdom spread of apple scar skin viroid from apple trees to fungi. Cells 2022, 11, 3686. [Google Scholar] [CrossRef] [PubMed]
- Walia, Y.; Dhir, S.; Zaidi, A.A.; Hallan, V. Apple scar skin viroid naked RNA is actively transmitted by the whitefly Trialeurodes vaporariorum. RNA Biol. 2015, 12, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Mound, L.A.; Halsey, S.H. Whitefly of the World: A Systematic Catalogue of the Aleyrodidae (Homoptera) with Host Plant and Natural Enemy Data; John Wiley and Sons: Hoboken, NJ, USA, 1978. [Google Scholar]
- Wisler, G.C.; Duffus, J.E.; Liu, H.Y.; Li, R.H. Ecology and epidemiology of whitefly-transmitted closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef]
- Tzanetakis, I.E.; Wintermantel, W.M.; Martin, R.R. First Report of beet pseudo yellows virus in strawberry in the United States: A second crinivirus able to cause pallidosis disease. Plant Dis. 2003, 87, 1398. [Google Scholar] [CrossRef]
- Tzanetakis, I.E.; Halgren, A.B.; Keller, K.E.; Hokanson, S.C.; Maas, J.L.; McCarthy, P.L.; Martin, R.R. Identification and detection of a virus associated with strawberry pallidosis disease. Plant Dis. 2004, 88, 383–390. [Google Scholar] [CrossRef]
- Salazar, L.F.; Muller, G.; Querci, M.; Zapata, J.L.; Owens, R.A. Potato yellow vein virus: Its host range, distribution in South America and identification as a crinivirus transmitted by Trialeurodes vaporariorum. Ann. Appl. Biol. 2000, 137, 7–19. [Google Scholar] [CrossRef]
- Jones, D.R. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol. 2003, 109, 195–219. [Google Scholar] [CrossRef]
- Wintermantel, W.M.; Hladky, L.L.; Cortez, A.A. Genome sequence, host range, and whitefly transmission of the torradovirus tomato necrotic dwarf virus. Acta Hortic. 2018, 1207, 295–302. [Google Scholar] [CrossRef]
- Maniataki, E.; Tabler, M.; Tsagris, M. Viroid RNA systemic spread may depend on the interaction of a 71-nucleotide bulged hairpin with the host protein VirP1. RNA 2003, 9, 346–354. [Google Scholar] [CrossRef]
- Gómez, G.; Pallás, V. Identification of an in vitro ribonucleoprotein complex between a viroid rna and a phloem protein from cucumber plants. Mol. Plant-Microbe Interact. 2001, 14, 910–913. [Google Scholar] [CrossRef] [PubMed]
- Eiras, M.; Nohales, M.A.; Kitajima, E.W.; Flores, R.; Daròs, J.A. Ribosomal protein L5 and transcription factor IIIA from Arabidopsis thaliana bind in vitro specifically Potato spindle tuber viroid RNA. Arch. Virol. 2011, 156, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Smith, H.N.; Ren, D.; Mudiyanselage, S.D.D.; Dawe, A.L.; Wang, L.; Wang, Y. Potato spindle tuber viroid modulates its replication through a direct interaction with a splicing regulator. J. Virol. 2018, 92, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Navarro, B.; Flores, R.; Di Serio, F. Advances in Viroid-Host Interactions. Annu. Rev. Virol. 2021, 8, 305–325. [Google Scholar] [CrossRef]
- Ma, J.; Mudiyanselage, S.D.D.; Hao, J.; Wang, Y. Cellular roadmaps of viroid infection. Trends Microbiol. 2023, 31, S0966-842X(23)00167-1. [Google Scholar] [CrossRef]
- Wang, Y.; Qu, J.; Ji, S.; Wallace, A.J.; Wu, J.; Li, Y.; Gopalan, V.; Ding, B. A land plant-specific transcription factor directly enhances transcription of a pathogenic noncoding RNA template by DNA-dependent RNA polymerase II. Plant Cell 2016, 28, 1094–1107. [Google Scholar] [CrossRef]
- Nohales, M.Á.; Flores, R.; Daròs, J.A. Viroid RNA redirects host DNA ligase 1 to act as an RNA ligase. Proc. Natl. Acad. Sci. USA 2012, 109, 13805–13810. [Google Scholar] [CrossRef]
- Castellano, M.; Pallas, V.; Gomez, G. A pathogenic long noncoding RNA redesigns the epigenetic landscape of the infected cells by subverting host histone deacetylase 6 activity. New Phytol. 2016, 211, 1311–1322. [Google Scholar] [CrossRef]
- Di Serio, F.; Owens, R.A.; Navarro, B.; Serra, P.; Martínez de Alba, Á.E.; Delgado, S.; Carbonell, A.; Gago-Zachert, S. Role of RNA silencing in plant-viroid interactions and in viroid pathogenesis. Virus Res. 2022, 323, 198964. [Google Scholar] [CrossRef]
- Navarro, J.A.; Vera, A.; Flores, R. A chloroplastic RNA polymerase resistant to tagetitoxin is involved in replication of avocado sunblotch viroid. Virology 2000, 268, 218–225. [Google Scholar] [CrossRef]
- Syller, J.; Marczewski, W.; Pawłowicz, J. Transmission by aphids of potato spindle tuber viroid encapsidated by potato leafroll luteovirus particles. Eur. J. Plant Pathol. 1997, 103, 285–289. [Google Scholar] [CrossRef]
- Walia, Y. Apple Scar Skin Viroid: Infectivity and Identification of Interacting Protein. Ph.D. Thesis, Guru Nanak Dev University, Amritsar, India, 6 January 2014. [Google Scholar]
- Scharf, K.D.; Heider, H.; Höhfeld, I.; Lyck, R.; Schmidt, E.; Nover, L. The tomato Hsf system: HsfA2 needs interaction with HsfA1 for efficient nuclear import and may be localized in cytoplasmic heat stress granules. Mol. Cell. Biol. 1998, 18, 2240–2251. [Google Scholar] [CrossRef] [PubMed]
- Kedersha, N.L.; Gupta, M.; Li, W.; Miller, I.; Anderson, P. RNA-Binding proteins Tia-1 and Tiar link the phosphorylation of Eif-2α to the assembly of mammalian stress granules. J. Cell Biol. 1999, 147, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Korber, P.; Zander, T.; Herschlag, D.; Bardwell, J.C. A new heat shock protein that binds nucleic acids. J. Biol. Chem. 1999, 274, 249–256. [Google Scholar] [CrossRef]
- Korber, P.; Stahl, J.M.; Nierhaus, K.H.; Bardwell, J.C. Hsp15: A ribosome-associated heat shock protein. EMBO J. 2000, 19, 741–748. [Google Scholar] [CrossRef]
- Staker, B.L.; Korber, P.; Bardwell, J.C.; Saper, M.A. Structure of Hsp15 reveals a novel RNA-binding motif. EMBO J. 2000, 19, 749–757. [Google Scholar] [CrossRef]
- Kanakala, S.; Kontsedalov, S.; Lebedev, G.; Ghanim, M. Plant-mediated silencing of the whitefly Bemisia tabaci cyclophilin B and heat shock protein 70 impairs insect development and virus transmission. Front. Physiol. 2019, 10, 557. [Google Scholar] [CrossRef]
- Andreason, S.A.; Arif, M.; Brown, J.K.; Ochoa-Corona, F.; Fletcher, J.; Wayadande, A. Single-target and multiplex discrimination of whiteflies (Hemiptera: Aleyrodidae) Bemisia tabaci and Trialeurodes vaporariorum with modified priming oligonucleotide thermodynamics. J. Econ. Entomol. 2017, 110, 1821–1830. [Google Scholar] [CrossRef]
- Ream, J.A.; Lewis, L.K.; Lewis, K.A. Rapid agarose gel electrophoretic mobility shift assay for quantitating protein: RNA interactions. Anal. Biochem. 2016, 511, 36–41. [Google Scholar] [CrossRef]
- Marcos, J.F.; Vilar, M.; Pérez-Payá, E.; Pallás, V. In vivo detection, RNA-binding properties and characterization of the RNA-binding domain of the p7 putative movement protein from carnation mottle carmovirus (CarMV). Virology 1999, 255, 354–365. [Google Scholar] [CrossRef]
- Diener, T.O. Viroids: The smallest known agents of infectious disease. Annu. Rev. Microbiol. 1974, 28, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Diener, T.O. Viroids and the nature of viroid diseases. In 100 Years of Virology; Calisher, C.H., Horzinek, M.C., Eds.; Springer: Vienna, Austria, 1999; pp. 203–220. [Google Scholar] [CrossRef]
- Ding, B. Viroids: Self-replicating, mobile, and fast-evolving noncoding regulatory RNAs. Wiley Interdiscip. Rev. RNA 2010, 1, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.J.; Domdey, H.; Lossow, C.; Jank, P.; Raba, M.; Alberty, H.; Sänger, H.L. Nucleotide sequence and secondary structure of potato spindle tuber viroid. Nature 1978, 273, 203–208. [Google Scholar] [CrossRef]
- Gómez, G.; Pallás, V. A long-distance translocatable phloem protein from cucumber forms a ribonucleoprotein complex in vivo with Hop stunt viroid RNA. J. Virol. 2004, 78, 10104–10110. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A.; Sun, L.; Randles, J.W. Modes of viroid transmission. Cells 2022, 11, 719. [Google Scholar] [CrossRef] [PubMed]
- Keese, P.; Symons, R.H. Domains in viroids: Evidence of intermolecular RNA rearrangements and their contribution to viroid evolution. Proc. Natl. Acad. Sci. USA 1985, 82, 4582–4586. [Google Scholar] [CrossRef] [PubMed]
- Henics, T.; Nagy, E.; Oh, H.J.; Csermely, P.; von Gabain, A.; Subjeck, J.R. Mammalian Hsp70 and Hsp110 proteins bind to RNA motifs involved in mRNA stability. J. Biol. Chem. 1999, 274, 17318–17324. [Google Scholar] [CrossRef]
- Laroia, G.; Cuesta, R.; Brewer, G.; Schneider, R.J. Control of mRNA decay by heat shock-ubiquitin-proteasome pathway. Science 1999, 284, 499–502. [Google Scholar] [CrossRef]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Meriin, A.B.; Sherman, M.Y.; Morimoto, R.I.; Massie, B. The chaperone function of hsp70 is required for protection against stress-induced apoptosis. Mol. Cell. Biol. 2000, 20, 7146–7159. [Google Scholar] [CrossRef]
- Balakrishnan, K.; De Maio, A. Heat shock protein 70 binds its own messenger ribonucleic acid as part of a gene expression self-limiting mechanism. Cell Stress Chaperones 2006, 11, 44–50. [Google Scholar] [CrossRef]
- Albihlal, W.S.; Gerber, A.P. Unconventional RNA-binding proteins: An uncharted zone in RNA biology. FEBS Lett. 2018, 592, 2917–2931. [Google Scholar] [CrossRef] [PubMed]
- Laksanalamai, P.; Robb, F.T. Small heat shock proteins from extremophiles: A review. Extremophiles 2004, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, E.; Asselbergh, B.; Rivera-Mejías, P.; Bervoets, S.; Vendredy, L.; De Winter, V.; Spaas, K.; de Rycke, R.; van Isterdael, G.; Impens, F.; et al. Small heat shock proteins operate as molecular chaperones in the mitochondrial intermembrane space. Nat. Cell Biol. 2023, 25, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Dissanayaka Mudiyanselage, S.D.; Park, W.J.; Wang, M.; Takeda, R.; Liu, B.; Wang, Y. A nuclear import pathway exploited by pathogenic noncoding RNAs. Plant Cell 2022, 34, 3543–3556. [Google Scholar] [CrossRef]
- Seo, H.; Kim, K.; Park, W.J. Effect of VIRP1 protein on nuclear import of citrus exocortis viroid (CEVd). Biomolecules 2021, 11, 95. [Google Scholar] [CrossRef]
- Chaturvedi, S.; Kalantidis, K.; Rao, A.L.N. A bromodomain-containing host protein mediates the nuclear importation of a satellite RNA of cucumber mosaic virus. J. Virol. 2014, 88, 1890–1896. [Google Scholar] [CrossRef]
- Montasser, M.S. First report of potential biological control of potato spindle tuber viroid disease by virus-satellite combination. Plant Dis. 1991, 75, 319E. [Google Scholar] [CrossRef]
- Yang, X.; Kang, L.; Tien, P. Resistance of tomato infected with cucumber mosaic virus satellite RNA to potato spindle tuber viroid. Ann. Appl. Biol. 1996, 129, 543–551. [Google Scholar] [CrossRef]
- Lindenau, S.; Winter, S.; Margaria, P. The amino-proximal region of the coat protein of cucumber vein yellowing virus (family Potyviridae) affects the infection process and whitefly transmission. Plants 2021, 10, 2771. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence | Target Region | References |
|---|---|---|---|
| VH23F | ATGGCATCTTTGAGAAGTCTCT | For sHSP22.98 amplification | In this study |
| VH23R | TTATGATTCCATTTTGTCACCTTTG | ||
| M13-F | GTAAAACGACGGCCAGT | For sequencing | In this study |
| M13-R | CAGGAAACAGCTATGAC | ||
| VH25 F | GGACACTCGCGCTTACTTTAC | For mtCOI amplification | [44] |
| VH 25 R | CAACATAACGTCGAGGCATCC | ||
| VH40 F | GTCGACGACGACAGGTGAGTT | For ASSVd amplification | [17] |
| VH40 R | GTCGACGAAGGCCGGTGAGAA | ||
| VH42 F | GGATCCATGGCATCTTTGAGAAGTCTCT | For sHsp22.98-pET28a construct | In this study |
| VH42 R | CTCGAGTTATGATTTCATTTTGTCACCTTTGC | ||
| VH65F | GAGCTCCAGCACTTTGGAATCGGTTTG | For tvHsp22.98-TRV construct | In this study |
| VH65R | CTCGAGGTTTGGCTTCGACGACTAGAT | ||
| To-PDS-F | CGGTCTAGAGGCACTCAACTTTATAAA CC | For To-PDS-TRV construct | In this study |
| To-PDS-R | CGGGGATCCCTTCAGTTTTCTGTCAAACC | ||
| CMV-F | ACCCTGAAACCGCCTGAAAT | For detection of CMV | In this study |
| CMV-R | TCCGAACTGTAACCCACACG | ||
| ToLCPalV-F | ATGGTGAAGCGTCCAGCAG | For detection of ToLCPalV | In this study |
| ToLCPalV-R | TTAATTTGTTACCGAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhary, S.; Selvaraj, V.; Awasthi, P.; Bhuria, S.; Purohit, R.; Kumar, S.; Hallan, V. Small Heat Shock Protein (sHsp22.98) from Trialeurodes vaporariorum Plays Important Role in Apple Scar Skin Viroid Transmission. Viruses 2023, 15, 2069. https://0-doi-org.brum.beds.ac.uk/10.3390/v15102069
Chaudhary S, Selvaraj V, Awasthi P, Bhuria S, Purohit R, Kumar S, Hallan V. Small Heat Shock Protein (sHsp22.98) from Trialeurodes vaporariorum Plays Important Role in Apple Scar Skin Viroid Transmission. Viruses. 2023; 15(10):2069. https://0-doi-org.brum.beds.ac.uk/10.3390/v15102069
Chicago/Turabian StyleChaudhary, Savita, Vijayanandraj Selvaraj, Preshika Awasthi, Swati Bhuria, Rituraj Purohit, Surender Kumar, and Vipin Hallan. 2023. "Small Heat Shock Protein (sHsp22.98) from Trialeurodes vaporariorum Plays Important Role in Apple Scar Skin Viroid Transmission" Viruses 15, no. 10: 2069. https://0-doi-org.brum.beds.ac.uk/10.3390/v15102069