

The Functional Implications of Broad Spectrum Bioactive Compounds Targeting RNA-Dependent RNA Polymerase (RdRp) in the Context of the COVID-19 Pandemic

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources and Search Strategy
- Concept #1: ‘RNA directed RNA polymerase’/exp OR (‘RNA-dependent RNA polymerase’ OR ‘RNA dependent RNA replicase’ OR ‘RNA directed RNA replicase’ OR ‘RNA replicase’):ab,ti,kw
- Concept #2: ‘coronavirus disease 2019’/exp OR (‘2019-nCoV disease’ OR ‘COVID 19’ OR ‘SARS-CoV-2 disease’ OR ‘SARS-CoV-2 infection’ OR ‘SARS-CoV-2 pneumonia’ OR ‘SARS-CoV2 disease’ OR ‘SARS-CoV2 infection’ OR ‘SARSCoV2 disease’):ab,ti,kw
- Concept #3: (‘inhibit’ OR ‘inhibition’ OR ‘inhibitor’):ab,ti,kw
- Concept #4: (‘mechanism’ OR ‘variant’ OR ‘structure’):ab,ti,kw
2.2. Eligibility Criteria and Study Selection
2.3. Data Charting and Synthesis
3. Results
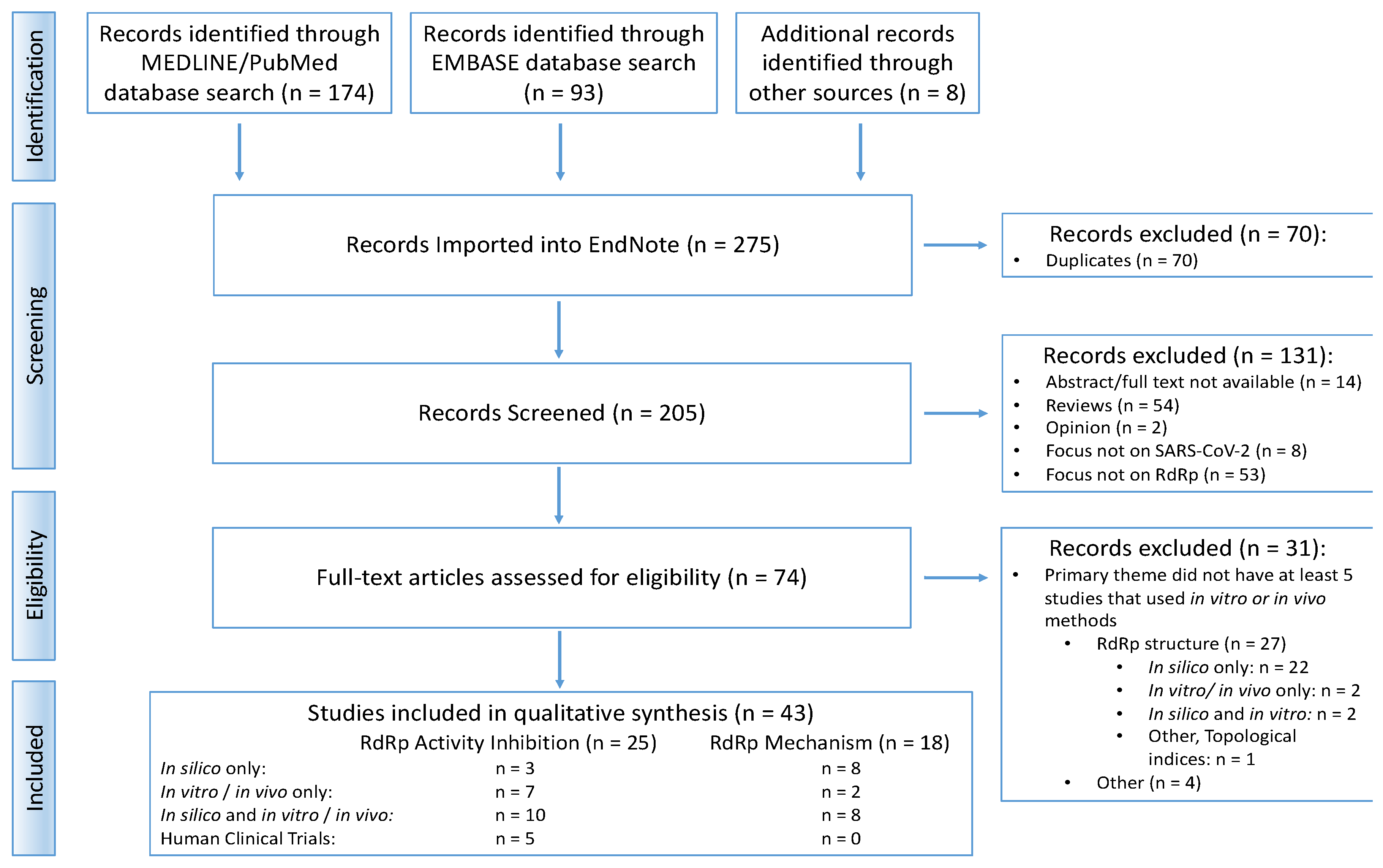
3.1. Overview of the Search Process
3.2. Summary of Findings
3.2.1. Adenosine-like Fungal Species
Cordycepin
- Through molecular dynamics simulations and pharmacokinetics predictions, Bibi et al. (2022) found that cordycepin has the potential to be a strong and stable RdRp inhibitor through hydrophobic interactions and hydrogen bonding with residues in the RdRp active site [26].
- Rabie (2022a) found that cordycepin has the ability to infiltrate SARS-CoV-2 genetic strands and stop the viral replication process by promoting ambiguous coding or early termination. The metabolic and structural similarities between cordycepin and nucleoside adenosine permit this incognito integration, as many enzymes cannot distinguish cordycepin triphosphate from ATP. Since the cordycepin molecule lacks a hydroxyl group, it disrupts SARS-CoV-2 RdRp activity, effectively terminating RNA synthesis and generating inactive, noninfectious material instead. Through in vitro bioactivity assays, Rabie found that cordycepin demonstrates stronger antiviral properties against SARS-CoV-2 than remdesivir. Long-lasting activities observed in vitro also validate the in silico findings that cordycepin exhibits high metabolic stability. Rabie suggested that cordycepin should be considered as a potential COVID-19 therapeutic due to its ability to target three main areas that contribute to SARS-CoV-2 infection: the spike protein, Mpro, and RdRp. Moreover, because cordycepin is nonspecific, in contrast to therapeutics that specifically target the spike protein, it can be effective against SARS-CoV-2 variants; this study showed inhibitory behaviors against variants of concern, including VOC-202012/01 [27].
- Li et al. (2022) noted that cordycepin may inhibit SARS-CoV-2 replication, because it replaces the 3′-hydroxyl group with a hydrogen atom, and once it is incorporated into the new RNA strand (after mimicking a natural nucleotide), it prompts replication termination. Among the series of adenosine analogs tested, including the HIV inhibitor didanosine and the leukemia drug clofarabine, didanosine and cordycepin may be considered the best candidates to thwart nucleotide addition at the SARS-CoV-2 RdRp active site [28].
3.2.2. Antibodies
Engineered Super-Antibody to Hepatitis C RdRp
- 4.
- Glab-ampai et al. (2022) evaluated the efficacy of an engineered super-antibody to hepatitis C RdRp against SARS-CoV-2 strains (Alpha, Beta, Delta, and Omicron), among other RNA viruses. Molecular docking and dynamic simulations revealed that this super-antibody can disrupt RdRp enzymatic activity through allosteric changes to the spatial conformation, thereby inhibiting viral replication. In vitro, cells infected with different SARS-CoV-2 variants were treated with the hepatitis C super-antibody, and viral RNA quantification showed a dose–response relationship; viral load reduction depends on the concentration of super-antibodies [29].
Poliovirus Vaccine That Induces Antibodies against Poliovirus RdRp
- 5.
- In a retrospective in vitro study, Comunale et al. (2021) examined whether serum samples from individuals recently vaccinated with the poliovirus vaccine could inhibit SARS-CoV-2 replication. Comparing sera from individuals who had not recently been vaccinated with those who had, enzyme-linked immunosorbent assays (ELISA) showed that antibody titers (including anti-RdRp) significantly increase with poliovirus vaccination. Using fluorescence-based RdRp assays, Comunale et al. also tested the ability of polio-immune sera to inhibit viral SARS-CoV-2 RNA synthesis and found that 76.5% of the tested samples could disrupt SARS-CoV-2 RdRp activity. Moreover, cytopathic effect (CPE)-based antiviral assays in Vero E6 cells demonstrated that polio-immune sera can inhibit SARS-CoV-2-induced CPE [30].
3.2.3. Antifungal Antibiotics
Biosynthetic Gene Cluster of NPP B1
- 6.
- Park et al. (2022) explored the antiviral capabilities of a biosynthetic gene cluster of NPP B1, a compound similar to amphotericin B, which is an antifungal medication that is often used to treat cryptococcal meningitis in HIV-positive patients [31]. Amphotericin B has previously demonstrated antiviral properties against enveloped viruses, such as HIV, Japanese encephalitis, and rubella [32,33]; therefore, Park et al. tested NPP B1′s ability to inhibit viral SARS-CoV-2 RNA synthesis using a fluorescence-based RdRp assay in vitro. NPP B1 can exhibit more than 50% inhibition of SARS-CoV-2 RNA synthesis compared to a control (no antibiotics), but other antibiotics (kanamycin, aminoglycoside, beta-lactam, or ampicillin) may not exhibit any inhibitory behaviors [34].
3.2.4. Antimalarials
Chloroquine and Derivatives (i.e., Hydroxychloroquine)
- 7.
- Through molecular docking and homology modeling, Nimgampalle et al. (2021) assessed the binding efficiencies of chloroquine and its derivatives to SARS-CoV-2 proteins. They found that chloroquine and hydroxychloroquine—antimalarial drugs that are also used to treat rheumatoid arthritis and lupus [35]— can bind to SARS-CoV-2 proteins like RdRp and have the potential to block viral replication by interfering with the catalytic active site. Moreover, derivatives such as CQN2H exhibited strong binding to RNA polymerase, as well as hydrogen bonding to RNA polymerase residues THR319 and THR394, suggesting that derivatives may also be potential SARS-CoV-2 inhibitors [36].
- 8.
- Maisonnasse et al. (2020) tested hydroxychloroquine’s antiviral properties against SARS-CoV-2 in Vero E6 cell cultures and in infected macaques. None of the analyzed tissues displayed significant inhibitory properties against SARS-CoV-2 either in vitro or in vivo. Hydroxychloroquine was tested as a prophylactic drug and as a possible treatment, both alone and in combination with azithromycin, an antibiotic used to fight bacterial infections. In vitro and in vivo results suggested that these therapeutic strategies may not be clinically beneficial, as the drugs do not significantly reduce viral loads or accelerate time to viral clearance [37].
- 9.
- Yao et al. (2020) assessed the antiviral and prophylactic properties of chloroquine and hydroxychloroquine against SARS-CoV-2 in Vero E6 cells in vitro. Physiologically based pharmacokinetic (PBPK) models were also used to analyze potential dosing regimens. Yao et al. found that hydroxychloroquine inhibits SARS-CoV-2 in vitro and displays stronger inhibitory behaviors than chloroquine against SARS-CoV-2. The most effective simulated dosing regimen was noted as a loading dose of 400 mg twice per day, followed by 4 days of 200 mg given twice daily. Yao et al. suggested that hydroxychloroquine’s prophylactic activities, antiviral potency against SARS-CoV-2, and safety profile make it a promising therapeutic candidate to combat cytokine storms in patients who are critically ill [38].
- 10.
- Gautret et al. (2020) studied the effect of hydroxychloroquine on respiratory viral loads in a non-randomized clinical trial with 36 patients. Validating prior research that chloroquine and its derivative hydroxychloroquine can inhibit SARS-CoV-2 viral replication in vitro, 70% of the patients who received hydroxychloroquine recovered from COVID-19 within six days compared to only 12.5% of patients in the untreated group. As such, researchers concluded hydroxychloroquine can successfully reduce SARS-CoV-2 viral loads, and the effects may be further bolstered by azithromycin [39].
3.2.5. Antioxidants
C60 Fullerene
- 11.
- Through molecular docking and simulations, Hurmach et al. (2021a) found that C60 fullerene, a compound often used as an antioxidant, is capable of inhibiting RdRp activity in silico. Dynamic simulations suggested two possible mechanisms of RdRp inhibition: (1) C60 fullerene blocks the RNA synthesis channel with its bulky carbon structure, thus thwarting normal replication processes, and (2) through stacking and steric interactions, C60 fullerene binds to two pockets that are critical to forming the RdRp-nsp8 complex, which assists in RNA synthesis. As SARS-CoV-2 RdRp (nsp12) cannot synthesize RNA on its own, it needs to form a complex with other nonstructural proteins, such as nsp7 and nsp8. Failure to create this complex results in an inability to replicate viral RNA [42].
- 12.
- Utilizing molecular dynamics and microscopic, spectroscopic, and in vitro methods, Hurmach et al. (2021b) evaluated the efficacy of nanostructure C60 fullerene against coronaviruses. In silico methods showed that the molecular structure of C60 fullerene is comprised of 60 carbon atoms that form a bulky, spherical shape. As such, C60 fullerene can block the coronavirus from entering the cell. It can also fill the RdRp binding pocket and effectively prevent RNA synthesis through stacking and steric interactions. Hurmach et al. found that C60 fullerene has the ability to decrease titers of infectious activity, can form stable complexes with SARS-CoV-2 RdRp through direct interactions, and can inhibit SARS-CoV-2 RdRp functionality [43].
Flavonoids (i.e., Quercetin, Theaflavin, and Luteolin)
- 13.
- Goc et al. (2022) evaluated the inhibition of RdRp activity from a mixture of natural compounds, consisting of vitamin C, N-acetylcysteine, resveratrol, theaflavin, curcumin, quercetin, naringenin, baicalin, and broccoli extract, in the hopes that a mixture of compounds could offer a multilayered protection effect against SARS-CoV-2. Using fluorescence-based RNA polymerase assay kits, Goc et al. found that the natural compound mixture is effective in inhibiting both the original and Omicron SARS-CoV-2 strains, though the strongest contributors to such inhibition are quercetin, theaflavin, and curcumin. Cytotoxicity and cell viability were tested in human alveolar epithelial cells, showing that quercetin and theaflavin are the most potent components targeting the RdRp complex. While the degree of inhibition was dose-dependent, an ideal dose of 0.1 mg/mL resulted in nearly 100% inhibition [45].
- 14.
- Munafò et al. (2022) explored whether quercetin and luteolin display antiviral properties against SARS-CoV-2 RdRp both in vitro and in silico. The anti-inflammatory properties of flavonoids suggest that SARS-CoV-2 inhibition may be demonstrated against several viral proteins, such as the Mpro and spike proteins. However, Munafò et al. observed that inhibitory behaviors are stronger against the RdRp protein than the Mpro and spike proteins at varying concentrations (total inhibition at 100 µM; 80% inhibition at 25 µM). Molecular dynamics simulations further validated the in vitro findings, suggesting that both quercetin and luteolin can form stable bonds at RdRp binding pockets [46].
Taroxaz-104
- 15.
- The Taroxaz-104 molecule is an antioxidant compound that is highly effective in transporting and mediating zinc uptake, which can play a direct role in inhibiting viral RdRp activity. Rabie (2021) noted that in silico predictions of Taroxaz-104 were highly effective against SARS-CoV-2 by interacting with several amino acids in the RdRp active site. In vitro, CPE-based antiviral bioassays in Vero E6 cells confirmed that Taroxaz-104 can successfully inhibit SARS-CoV-2 replication and transcription more than the reference compound, GS-443902, the active metabolite of RdRp inhibitor remdesivir. Furthermore, Taroxaz-104 demonstrates 43 times the potency of GS-443902 against the VOC-202012/01 SARS-CoV-2 variant. The ability of Taroxaz-104 to hinder SARS-CoV-2 RNA replication processes and inhibit CPEs indicates the powerful potential to repurpose this drug against SARS-CoV-2 variants. As fundamental mutations to the SARS-CoV-2 spike protein have been observed across variants, Rabie recommended utilizing drugs that inhibit key SARS-CoV-2 proteins that are not located on the viral surface, such as RdRp [47].
3.2.6. Antiparasitics
Suramin
- 16.
- Yin et al. (2021) explored whether and how suramin, a drug used to treat African sleeping sickness and river blindness [48], could inhibit SARS-CoV-2 replication through in silico and in vitro methods. Structural modeling demonstrated RdRp–suramin binding at two sites, both of which are needed for viral replication. At one site, where RNA template strands would bind, the RdRp–suramin complex blocks natural substrates from entering the active site, directly ceasing viral replication. Then, near the RdRp catalytic site, the RdRp–suramin complex conflicts with the RNA primer strand and indirectly prevents RdRp enzymatic processes from occurring. In vitro experiments in Vero E6 cells confirmed that suramin can inhibit SARS-CoV-2 viral replication [49].
3.2.7. Antivirals
BMS-986094 (Developed for Hepatitis C) [50]
- 17.
- Chemical structure modeling by Jimenez-Guardeño et al. (2022) suggested that BMS-986094, an RdRp inhibitor initially developed for the hepatitis C virus [51,52], and different forms of vitamin B12 may be effective against SARS-CoV-2. While quadratic unbounded binary optimization (QUBO) and Tanimoto models predicted inhibitory behaviors in silico, SARS-CoV-2 RNA polymerase and cytotoxicity assays in vitro also suggested that each tested compound could effectively disrupt the viral replication of four strains: SARS-CoV-2 Strain England 2 (England 02/2020/407073), B.1.1.7 (Alpha), B.1.351 (Beta), and B.1.617.2 (Delta) [53].
Remdesivir (Developed for the Ebola Virus) [54]
- 18.
- Pirzada et al. (2021) examined U.S. Food and Drug Administration-approved RdRp inhibitors as potential therapeutics for SARS-CoV-2. In silico methods suggested that remdesivir, a drug developed to treat Ebola [55,56], as well as two hepatitis C drugs, ledipasvir and paritaprevir [57,58], may exhibit strong interactions with SARS-CoV-2 RdRp. Subsequently, the antiviral properties of all three drugs were tested in Vero E6 cell cultures, and the findings indicated that, compared to ledipasvir and paritaprevir, remdesivir is the most potent SARS-CoV-2 RdRp inhibitor [59].
- 19.
- Vangeel et al. (2022) found that remdesivir, molnupiravir, and nirmatrelvir (the antiviral active ingredient in Paxlovid), three SARS-CoV-2 antivirals that target either the RdRp or Mpro proteins, demonstrate similar efficacy and potency in vitro across five of the main SARS-CoV-2 strains (Alpha, Beta, Gamma, Delta, and Omicron). Some of the structural characteristics that distinguish the original (Alpha) SARS-CoV-2 strain from variants of concern are associated with amino acid changes that are far from the RdRp and Mpro active sites. Thus, RdRp and Mpro inhibitor drugs can remain effective regardless of which SARS-CoV-2 strain one may be exposed to, because the target proteins are highly conserved. Vangeel et al. suggested that more antivirals that target proteins other than the commonly addressed spike protein, such as RdRp or Mpro, should be strongly considered, as the drugs are more likely to remain effective amidst new variants [60].
- 20.
- Through molecular docking methods, Choudhury et al. (2020) investigated which known RdRp inhibitors of other viruses may be effective in disrupting SARS-CoV-2 replication in silico. Ligand–receptor interaction docking scores predicted which compounds could form stable complexes and/or have high affinities for the RdRp active site and thus may interfere with the usual enzymatic activity processes. Remdesivir was noted as the strongest inhibitor, having the ability to compete with natural substrates and become incorporated into RNA chains, thereby terminating replication. Chlorhexidine, a drug commonly used for dental plaques, gingivitis, and tonsillitis, also demonstrated strong affinities for the RdRp active site, though the mechanism to inhibit RdRp was not by terminating replication via substrate competition but rather by actively blocking the RdRp cavity and preventing further replication. Additionally, Choudhury et al. predicted five other nucleoside analogs that are similar to remdesivir that could be effective in inhibiting SARS-CoV-2 RdRp activity: sofosbuvir, ribavirin, penciclovir, ganciclovir, and favipiravir [61].
- 21.
- Zhang and Zhou (2020) found that remdesivir may act as an effective inhibitor against SARS-CoV-2 in the same manner that it can stop reproduction of the Ebola virus. Once remdesivir is metabolized in the body (hydrolyzed and phosphorylated), it can act as a substitute for ATP, given its similar structure to adenosine, and thus may serve as a binding substrate that can obstruct the “grip” of SARS-CoV-2 RdRp, prompting the cessation of RdRp activity and RNA replication [62].
- 22.
- To better understand the interactions between remdesivir and SARS-CoV-2 RdRp residues that may contribute to viral replication inhibition, Koulgi et al. (2020) compared RdRp conformations in the apo form (unbound state) with the remdesivir-bound form in silico. Ensemble docking and molecular dynamics simulations revealed opposite movements: in the apo form, the template entry site is exposed to give natural nucleotides access; in the remdesivir-bound form, hydrogen bonding and residue contacts obstruct natural nucleotides from accessing the cavity, as the template entry site is filled by remdesivir, prompting viral replication termination [63].
- 23.
- Nguyen et al. (2020) were interested in how remdesivir binds to two key SARS-CoV-2 proteins, RdRp and Mpro, in silico. Molecular docking predicted that Mpro would have a stronger bond with remdesivir than RdRp would, but molecular dynamics simulations showed the converse to be true. Nguyen et al. suggested that this discrepancy may be due to molecular docking methods incorrectly estimating the binding energies compared to more advanced methods like molecular dynamics simulations. Since remdesivir’s binding site is close to the RdRp active site, remdesivir can easily form hydrogen bonds, as well as approach and obstruct the open RdRp cavity. Nguyen et al. also found that remdesivir binds to each key protein in a different manner: electrostatic interactions stabilize the RdRp–remdesivir complex, while van der Waals interactions are responsible for stabilizing Mpro binding [64].
- 24.
- Aranda et al. (2022) sought to characterize the mechanism of reaction in SARS-CoV-2 RdRp with natural triphosphates and remdesivir triphosphates in silico. Contrary to the popular hypothesis that remdesivir inhibits SARS-CoV-2 viral replication by competing with ATP, Aranda et al. suggested that a covalent bond with RdRp at position i+4 may be responsible for the observed delayed inhibition instead of the widely accepted steric clashing. Moreover, the remdesivir triphosphate incorporation rate inside RdRp can be slower than that of natural triphosphate (ATP). Through equilibrium trajectories, Aranda et al. noted that remdesivir triphosphate is the same size and shape as adenosine, exhibits strong interactions with uridine triphosphate (UTP), and is well suited for the RdRp active site; the simulations displayed remdesivir triphosphate incorporation into nascent RNA in front of UTP, contradicting findings that the substrate blocks RdRp. While the in silico findings suggested new mechanisms to explain inhibitory behaviors, the data nonetheless validated that remdesivir can be an effective drug against SARS-CoV-2 by targeting RdRp [65].
- 25.
- Tchesnokov et al. (2020) studied remdesivir’s role in SARS-CoV-2 viral replication inhibition in vitro. They found that remdesivir is capable of disrupting SARS-CoV-2 RdRp enzymatic processes once remdesivir triphosphate is incorporated into the RNA primer strand, as it is later used as a template for RNA synthesis. Once the remdesivir residue is in the template strand, nucleotide incorporation is notably diminished. Moreover, delayed chain termination may be observed with low nucleoside triphosphate (NTP) concentrations; when NTP is increased, the translocation equilibrium changes, which consequently unveils nucleotide binding sites. As such, increasing UTP and ATP, or the UTP, ATP, and cytidine triphosphate (CTP) concentrations, erodes the inhibitory properties, enhances read-through, and enables full-length RNA product development [66].
- 26.
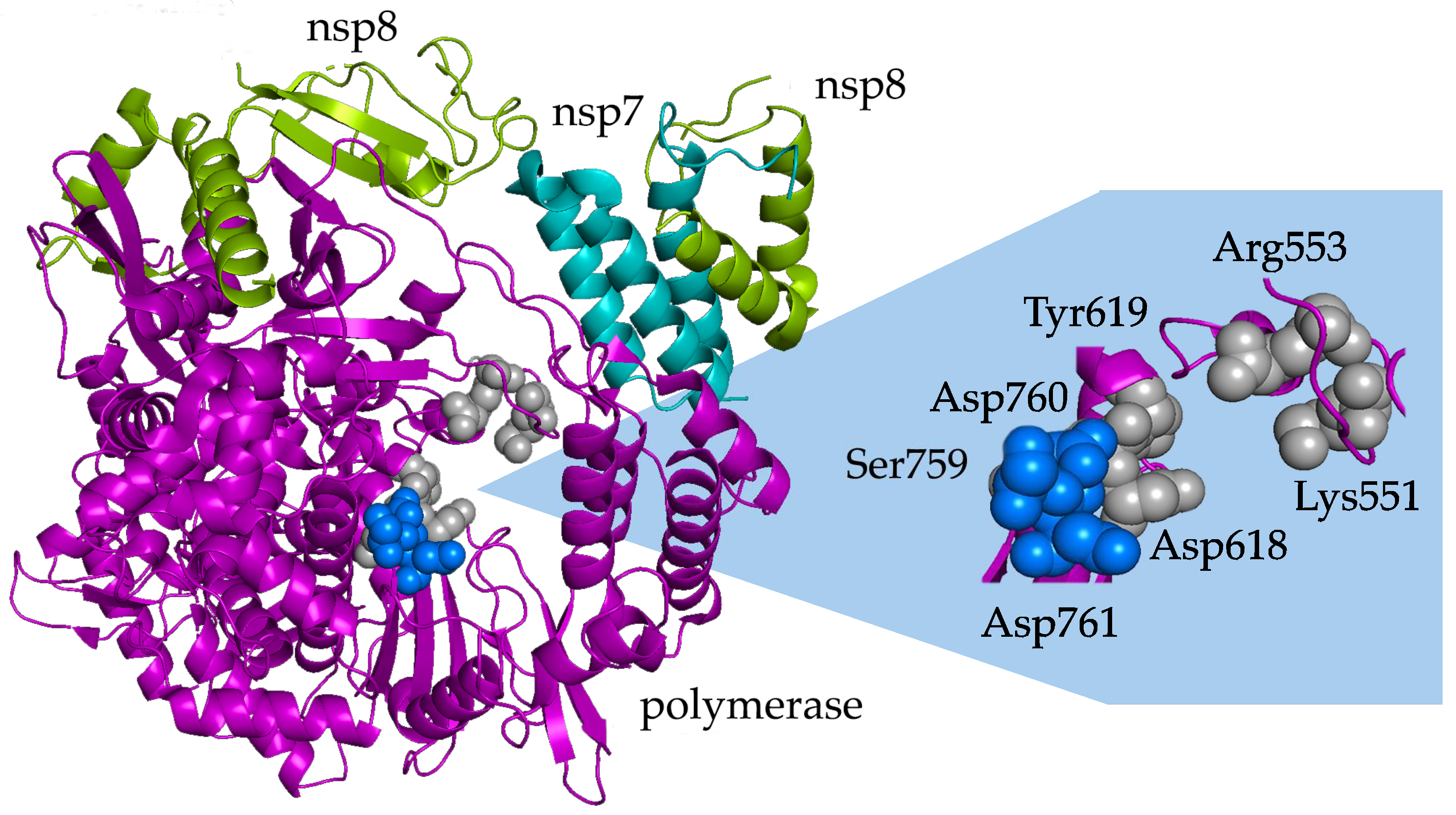
- Through sequence analyses, Stevens et al. (2022) explored nonsynonymous mutations (nucleotide sequence changes) in SARS-CoV-2 RdRp and sought to understand the possible mechanisms behind the resistance to remdesivir. Thus far, SARS-CoV-2 variants in the natural environment have not exhibited resistance to remdesivir, but there is a possibility that remdesivir resistance mutations could arise in minority variants. Stevens et al. found that the mutations V792I and S759A in SARS-CoV-2 RdRp could exhibit resistance to remdesivir. The V792I mutant may minimize the template-dependent inhibition of viral replication by lowering UTP concentrations. The S759A mutant is significantly less likely to use remdesivir triphosphate as a substrate compared to ATP (10.5-fold decrease), thereby diminishing the utility of the RdRp inhibitor. However, the S759A mutation was not found among the Delta and Omicron variant sequences, and other substitutions were identified at low frequencies, suggesting that contemporary variants do not show resistance to the RdRp inhibitor remdesivir [67].
Molnupiravir (Developed for the Influenza Virus) [71] and Derivatives (i.e., 4′-Fluorouridine and N4-Hydroxycytidine)
- 27.
- As mentioned above, Vangeel et al. (2022) found that molnupiravir has efficacy and potency comparable to remdesivir and nirmatrelvir (found in Paxlovid) in vitro across the SARS-CoV-2 strains (Alpha, Beta, Gamma, Delta, and Omicron). As the amino acid changes that differentiate strains are far enough away from the RdRp active site, RdRp inhibitors can continue to successfully interact with the RdRp protein and inhibit viral replication [60].
- 28.
- Sourimant et al. (2022) analyzed the antiviral properties of 4′-fluorouridine (4′-FlU; EIDD-2749), a ribonucleoside analog and derivative of molnupiravir, against several RNA viruses, including respiratory syncytial virus (RSV) and SARS-CoV-2. In vitro, Sourimant et al. found that 4′-FIU incorporation can prompt an antiviral effect of RNA polymerase stalling. In vivo, viral titers in nasal turbinate tissue revealed that a daily dose of 20 mg/kg of 4′-FIU can effectively reduce the viral load and inhibit SARS-CoV-2 replication in infected ferrets across the Alpha, Gamma, and Delta variants [74].
- 29.
- N4-hydroxycytidine (NHC; EIDD-1931), a ribonucleoside analog, has antiviral properties against several RNA viruses, such as influenzas A and B, Ebola, and Venezuelan equine encephalitis virus [75,76]. Sheahan et al. (2020) explored whether NHC is capable of inhibiting SARS-CoV-2 in vitro and in vivo. Utilizing human airway epithelial cell cultures, they found that NHC reduces infectious titers and viral SARS-CoV-2 RNA in a dose-dependent manner. Additionally, a prophylactic dose escalation in vivo study in mice suggested that an orally bioavailable NHC prodrug (EIDD-2801, also known as molnupiravir) notably decreases lung hemorrhaging, SARS-CoV lung titers, and viral loads. Surrogate markers for pulmonary obstruction can also be enhanced, though clinical improvement may be dependent on when NHC is administered post-infection [73].
- 30.
- Gordon et al. (2021) evaluated the mechanism of how molnupiravir (NHC 5′-triphosphate (NHC-TP)) promotes antiviral effects against SARS-CoV-2 in vitro. They observed that NHC-TP competes with CTP for incorporation (a C-analog), but when NHC is in the monophosphate form (NHC-MP) and embedded in the template strand, it can compete with CTP and UTP, thus favoring the incorporation of guanosine triphosphate (GTP) and ATP. Then, when GTP is incorporated opposite NHC-MP, subsequent nucleotides are unable to be incorporated into the strand (i.e., viral inhibition commences). When NHC-MP competes with UTP and favors the incorporation of ATP, NHC:A base pairs lead to transition mutations in G to A and in C to U. These mutation frequencies ultimately produce dysfunctional genomes that then disrupt normal replication processes. Thus, Gordon et al. showed that molnupiravir is responsible for mutagenesis when embedded into the template strand. Conversely, when NHC-MP is embedded in the RNA primer strand, it does not facilitate RNA synthesis inhibition [72].
- 31.
- Khoo et al. (2023) evaluated the safety and efficacy of molnupiravir against various SARS-CoV-2 strains in a phase II randomized, double blinded placebo-controlled clinical trial. Those who were treated with molnupiravir recovered from COVID-19 significantly faster than those who received the placebo (8 days versus 11 days, respectively). Additionally, no participants in the treatment group were hospitalized compared to four participants in the placebo group. While preliminary evidence indicates molnupiravir is well tolerated and may be effective against COVID-19, the predefined threshold was not reached, prompting Khoo et al. to conclude that the evidence may not be conclusive [77].
Nucleoside Analogs (i.e., Didanosine, Fludarabine, Vidarabine, and Favipiravir)
- 32.
- To understand how 2′- and 3′-ribose modifications of adenosine analogs may inhibit SARS-CoV-2 replication, Li et al. (2022) conducted molecular dynamics simulations of nucleotide analogs that target RdRp, including drugs used for leukemia (clofarabine and fludarabine) [78,79], HIV inhibitors (didanosine and 2′,3′-didehydro-2′,3′-dideoxyadenosine) [80,81], and inhibitors used against herpes simplex virus and varicella-zoster virus (vidarabine) [82]. They found that a 2′-ribose modification affects not only the binding stability but also the substrate incorporation efficiency into the new RNA strand. Additionally, 3′-ribose modifications, such as a 3′-hydroxyl group removal, as seen in didanosine and 2′,3′-didehydro-2’,3’-dideoxyadenosine, can also enable inhibition. Didanosine and cordycepin, natural adenosine analogs found in fungal species, were noted as the strongest agents to disrupt nucleotide addition at the SARS-CoV-2 RdRp active site [28].
- 33.
- Rabie (2022b) investigated the effectiveness of didanosine against SARS-CoV-2 RdRp both in vitro and in silico. Molecular docking and computational interpretations revealed that didanosine interacts with a fundamental pocket of the RdRp active site and is incorporated into new viral strands instead of natural substrates (ATP and GTP), thereby creating defective viral particles and prompting the termination of SARS-CoV-2 RNA synthesis. The molecular makeup of didanosine aids in these processes, as it lacks a 3′-hydroxyl group, and such a deficiency is often associated with premature RNA synthesis termination. In silico data confirmed what Rabie observed in vitro through antiviral assays in Vero E6 cells against the VOC-202012/01 SARS-CoV-2 variant strain: didanosine not only suppressed viral replication but also proved to be more potent (by 5.0-6.8 times) than remdesivir against the SARS-CoV-2 variant [68].
- 34.
- Sirijatuphat et al. (2022) investigated the efficacy of favipiravir, a nucleoside-based antiviral drug that is commonly used to treat influenza, among 96 patients that had COVID-19, had not received COVID-19 vaccinations, and did not have pneumonia at the onset of the study. The potential effects on viral clearance, their clinical condition, and the risk of developing pneumonia were evaluated. Validating prior in vitro analyses of inhibitory activity, patients who received favipiravir showed clinical improvements in two days compared to those in the control arm at fourteen days. Sirijatuphat et al. asserted favipiravir can expedite recovery in mild COVID-19 cases and is safe for short-term use [83].
- 35.
- Udwadia et al. (2020) conducted a phase III clinical trial testing the efficacy, safety, and clinical benefits of favipiravir among 150 patients. The median time of SARS-CoV-2 viral shedding was five days for those who received favipiravir compared to seven days for those in the control arm. While this difference was not statistically significant, the results were clinically meaningful for the secondary endpoint, the median time to a clinical cure, with three days for patients who received favipiravir and five days for those who did not. Researchers suggest favipiravir is not only safe and well-tolerated but may also be beneficial in mitigating the effects of mild-to-moderate COVID-19 [84].
- 36.
- Shinkai et al. (2021) assessed the efficacy and safety of favipiravir against COVID-19 among patients with moderate pneumonia in a randomized placebo-controlled Phase III trial. Researchers evaluated the median time of recovery, as measured by an improvement in temperature, oxygen saturation levels (SpO2), findings on chest imaging, and testing negative for SARS-CoV-2. Patients who were treated with favipiravir recovered significantly faster than those who received the placebo (11.9 days versus 14.7 days, respectively), suggesting favipiravir may be considered clinically effective for COVID-19 pneumonia patients [85].
Sofosbuvir (Developed for Hepatitis C) [86]
- 37.
- 38.
- Yuan et al. (2021) analyzed how five different 2′-modified nucleotides, including sofosbuvir and gemcitabine, a chemotherapy drug [90], may affect the polymerization process and/or inhibitory behaviors of SARS-CoV-2 RdRp. Considering 2′-O-methyl UTP, 2′-C-methyl CTP, gemcitabine, ara-UTP, and sofosbuvir, Yuan et al. found that the size and shape of 2′-methyl substitutions in 2′-C-methyl CTP and sofosbuvir greatly disrupt the polymerization processes once they are incorporated into the active site, so much so that the polymerase chain is instantly terminated from steric hindrance. On a lesser scale, 2′-O-methyl UTP may serve as a partial chain terminator, because molecular binding may be hindered upon incorporation. Furthermore, smaller 2′-substitutions, such as those in ara-UTP and gemcitabine, do not effectively inhibit viral replication [88].
- 39.
- Similarities between SARS-CoV-2 RdRp and hepatitis C enzymes NS5A (protease) and NS5B (RdRp) led Sacramento et al. (2021) to investigate whether drugs targeting these enzymes, such as daclatasvir and sofosbuvir [91,92,93], could also inhibit SARS-CoV-2 viral replication. Molecular docking suggested that both drugs can interact with SARS-CoV-2 RNA residues through hydrogen bonding and indicated that steric interactions exist for both compounds. In vitro assays in Vero E6 cells, human lung epithelial cells, and human hepatoma lineage cells demonstrated that daclatasvir can inhibit SARS-CoV-2 RNA synthesis and that sofosbuvir has antiviral capabilities, though the effects may be stronger when combined with daclatasvir than when administered alone. The inhibitory mechanisms differ between the two drugs: sofosbuvir acts as a chain terminator, and daclatasvir targets folds in the RNA secondary structures, inciting destabilization and disrupting polymerase reactions. Sacramento et al. suggested further investigation into dosing for SARS-CoV-2 treatment, as effective doses may be higher than those currently prescribed to treat hepatitis C [87].
- 40.
- SARS-CoV-2 has an exonuclease-based proofreader (nonstructural protein 14, nsp-14) that detects and removes incorrect nucleotides that have been incorporated into new viral strands. Many RdRp inhibitor drugs that exhibit antiviral properties against SARS-CoV-2 rely on the ability to mimic natural substrates, such as ATP, in order to become incorporated into viral RNA and prompt replication termination; therefore, their drug efficacy may be affected by nsp-14′s proofreading abilities. Jockusch et al. (2020) examined SARS-CoV-2 RNA that was terminated by either sofosbuvir, a hepatitis C inhibitor, or remdesivir, a therapeutic for Ebola, and compared which may be more resistant to nsp-14 and avoid removal so RNA synthesis could cease. Considering the active forms of each drug, Jockusch et al. found that sofosbuvir triphosphate is removed less frequently than remdesivir triphosphate, suggesting that sofosbuvir may be a promising therapeutic against SARS-CoV-2, because it is more resistant to nsp-14 than remdesivir [70].
Uracil Derivatives (Used for HIV and Hepatitis C) [94,95]
- 41.
- As non-nucleoside uracil derivatives have previously been effective antiviral agents against HIV, hepatitis C, and varicella zoster virus [96], Siniavin et al. (2022) investigated whether N1,N3-disubstituted derivatives of uracil could be utilized against SARS-CoV-2. In silico, compound 876 displayed the best score to predict the disruption of RdRp functionality (through placement in the hydrophobic pocket of the RNA cleft). In vitro, Siniavin et al. found that compound 876 can inhibit SARS-CoV-2 RdRp at varying concentrations (100% inhibition at 100 µM and 40% inhibition at 50 µM), and compounds 871, 874, and 1007 could decrease viral replication of the SARS-CoV-2 variants of concern (Beta, Delta, and Omicron) [97].
CRISPR Genome Editing
- 42.
- Abbott et al. (2020) proposed an alternative method to targeting SARS-CoV-2 to the currently available vaccines that aim to prime and help the immune system recognize proteins like the SARS-CoV-2 spike protein [98]. Instead, they suggested using prophylactic antiviral CRISPR in human cells (PAC-MAN) to edit the protein sequences of SARS-CoV-2 RNA so that intracellular viral genome templates that are needed for viral replication may degrade prophylactically. While in silico models identified specific, highly conserved SARS-CoV-2 genomic regions for the Cas13d enzyme to target, in vitro human lung epithelial cell cultures confirmed that PAC-MAN could inhibit viral gene expression across potential variants and mutations that may arise naturally through antigenic drift. Abbott et al. also employed this strategy against the influenza A virus and found that, by targeting the highly conserved RdRp genes, one could inhibit viral production and prompt viral genome deterioration, thereby reducing the viral load in human lung epithelial cells in vitro [99].
3.2.8. Vitamins, Natural Compounds, and Extracts
Vitamin B12
- 43.
- Narayanan and Nair (2020) utilized homology modeling and molecular dynamics simulations to identify vitamin B12 as a potential SARS-CoV-2 RdRp inhibitor. Since the binding site for vitamin B12 overlaps with that of NTPs, simulations predicted that vitamin B12 could bind to the SARS-CoV-2 RdRp active site in place of NTPs, therefore inhibiting polymerization processes. An analysis of the binding energies confirmed that vitamin B12 can bind to the RdRp active site with significant affinity [100].
- 44.
- As mentioned above, Jimenez-Guardeño et al. (2022) found that different forms of vitamin B12 can inhibit SARS-CoV-2 viral replication in silico and in vitro across variant strains, including SARS-CoV-2 Strain England 2 (England 02/2020/407073), B.1.1.7 (Alpha), B.1.351 (Beta), and B.1.617.2 (Delta) [53].
Gossypol (Used in Cancer Therapies) [101,102,103,104]
Hypericin (Used in Cancer Therapies) [107]
- 45.
- Matos et al. (2021) utilized structure-based virtual screening to identify potential therapeutics against SARS-CoV-2 that target proteins critical to the viral replication process, such as RdRp. They noted six compounds with high affinities for the SARS-CoV-2 RdRp active site, most notably hypericin, which is a naturally occurring substance found in St. John’s wort that has previously been used as an antiviral agent and in cancer therapies [108]. In a dose–response in vitro experiment, Matos et al. demonstrated that, at the highest tested concentrations, hypericin can reduce viral RNA synthesis up to 96%. Moreover, LDH cytotoxicity assays were conducted in cultured cells and showed no significant plasma membrane damage, further confirming that hypericin may be a promising therapeutic agent against COVID-19 [109].
Corilagin (Used in Cancer Therapies) [110]
- 46.
- Li et al. (2021) examined corilagin, a non-nucleoside RdRp inhibitor that is found in medicinal plants and has been used to diminish cancerous cell growth [111], and its ability to inhibit SARS-CoV-2 polymerization both in silico and in vitro. Molecular modeling predicted that corilagin would inhibit RdRp activity by docking at the palm subdomain of RdRp, thereby blocking NTP from entering the SARS-CoV-2 active site. In both cell-free and cell-based assays, corilagin can disrupt SARS-CoV-2 viral replication. Moreover, corilagin and remdesivir demonstrate dose–response relationships, and when both are used in treatments, there is an additive effect in inhibiting RdRp [112].
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caron, C. With Omicron Comes Uncertainty. Here’s How to Handle It. The New York Times, 2 December 2021. Available online: https://www.nytimes.com/2021/12/02/well/mind/omicron-variant-questions.html(accessed on 30 October 2023).
- COVID-19 Variants: What You Should Know. Johns Hopkins Medicine. Updated 8 April 2022. Available online: https://www.hopkinsmedicine.org/health/conditions-and-diseases/coronavirus/a-new-strain-of-coronavirus-what-you-should-know (accessed on 30 October 2023).
- Yewdell, J.W. Antigenic drift: Understanding COVID-19. Immunity 2021, 54, 2681–2687. [Google Scholar] [CrossRef] [PubMed]
- Offit, P.A. Bivalent COVID-19 Vaccines—A Cautionary Tale. N. Engl. J. Med. 2023, 388, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Tatar, M.; Shoorekchali, J.M.; Faraji, M.R.; Seyyedkolaee, M.A.; Pagán, J.A.; Wilson, F.A. COVID-19 vaccine inequality: A global per spective. J. Glob. Health 2022, 12, 03072. [Google Scholar] [CrossRef] [PubMed]
- Pathania, S.; Rawal, R.K.; Singh, P.K. RdRp (RNA-dependent RNA polymerase): A key target providing anti-virals for the man agement of various viral diseases. J. Mol. Struct. 2022, 1250, 131756. [Google Scholar] [CrossRef] [PubMed]
- Subissi, L.; Posthuma, C.C.; Collet, A.; Zevenhoven-Dobbe, J.C.; Gorbalenya, A.E.; Decroly, E.; Snijder, E.J.; Canard, B.; Imbert, I. One severe acute respiratory syndrome coronavirus protein complex integrates pro cessive RNA polymerase and exonuclease activities. Proc. Natl. Acad. Sci. USA 2014, 111, E3900–E3909. [Google Scholar] [CrossRef] [PubMed]
- Kirchdoerfer, R.N.; Ward, A.B. Structure of the SARS-CoV nsp12 polymerase bound to nsp7 and nsp8 co-factors. Nat. Commun. 2019, 10, 2342. [Google Scholar] [CrossRef] [PubMed]
- Ando, T.; Imamura, H.; Suzuki, R.; Aizaki, H.; Watanabe, T.; Wakita, T.; Suzuki, T. Visualization and measurement of ATP levels in living cells replicating hepatitis C vi rus genome RNA. PLoS Pathog. 2012, 8, e1002561. [Google Scholar] [CrossRef]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Mishra, A.; Rathore, A.S. RNA dependent RNA polymerase (RdRp) as a drug target for SARS-CoV-2. J. Biomol. Struct. Dyn. 2022, 40, 6039–6051. [Google Scholar] [CrossRef]
- Ago, H.; Adachi, T.; Yoshida, A.; Yamamoto, M.; Habuka, N.; Yatsunami, K.; Miyano, M. Crystal structure of the RNA-dependent RNA polymerase of hepatitis C virus. Structure 1999, 7, 1417–1426. [Google Scholar] [CrossRef]
- Hansen, J.L.; Long, A.M.; Schultz, S.C. Structure of the RNA-dependent RNA polymerase of poliovirus. Structure 1997, 5, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Yap, T.L.; Xu, T.; Chen, Y.L.; Malet, H.; Egloff, M.P.; Canard, B.; Vasudevan, S.G.; Lescar, J. Crystal structure of the dengue virus RNA-dependent RNA polymerase catalytic domain at 1.85-angstrom resolution. J. Virol. 2007, 81, 4753–4765. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Gong, P. A Structure-Function Diversity Survey of the RNA-Dependent RNA Polymerases From the Positive-Strand RNA Viruses. Front. Microbiol. 2019, 10, 1945. [Google Scholar] [CrossRef] [PubMed]
- Warren, T.K.; Kane, C.D.; Wells, J.; Stuthman, K.S.; Tongeren, S.A.V.; Garza, N.L.; Donnelly, G.; Steffens, J.; Gomba, L.; Weidner, J.M.; et al. Remdesivir is efficacious in rhesus monkeys exposed to aerosolized Ebola virus. Sci. Rep. 2021, 11, 19458. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Gottlieb, R.L.; Vaca, C.E.; Paredes, R.; Mera, J.; Webb, B.J.; Perez, G.; Oguchi, G.; Ryan, P.; Nielsen, B.U.; Brown, M.; et al. Early Remdesivir to Prevent Progression to Severe COVID-19 in Outpatients. N. Engl. J. Med. 2022, 386, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Amicone, M.; Borges, V.; Alves, M.J.; Isidro, J.; Zé-Zé, L.; Duarte, S.; Vieira, L.; Guiomar, R.; Gomes, J.P.; Gordo, I. Mutation rate of SARS-CoV-2 and emergence of mutators during experimental evolu tion. Evol. Med. Public Health 2022, 10, 142–155. [Google Scholar] [CrossRef]
- In Vitro. National Institutes of Health, National Cancer Institute. Reviewed 24 January 2023. Available online: https://www.cancer.gov/publications/dictionaries/cancer-terms/def/in-vitro (accessed on 30 October 2023).
- Aftab, S.O.; Ghouri, M.Z.; Masood, M.U.; Haider, Z.; Khan, Z.; Ahmad, A.; Munawar, N. Analysis of SARS-CoV-2 RNA-dependent RNA polymerase as a potential thera peutic drug target using a computational approach. J. Transl. Med. 2020, 18, 275. [Google Scholar] [CrossRef]
- Singh, P.; Tripathi, M.K.; Yasir, M.; Khare, R.; Shrivastava, R. In silico identification of promising inhibitor against RNA-dependent RNA polymerase target of SARS-CoV-2. Mol. Biol. Res. Commun. 2021, 10, 131–140. [Google Scholar] [CrossRef]
- Vicenti, I.; Zazzi, M.; Saladini, F. SARS-CoV-2 RNA-dependent RNA polymerase as a therapeutic target for COVID-19. Expert. Opin. Ther. Pat. 2021, 31, 325–337. [Google Scholar] [CrossRef]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic re views. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.; Hasan, M.M.; Wang, Y.B.; Papadakos, S.P.; Yu, H. Cordycepin as a Promising Inhibitor of SARS-CoV-2 RNA Dependent RNA Polymerase (RdRp). Article. Curr. Med. Chem. 2022, 29, 152–162. [Google Scholar] [CrossRef]
- Rabie, A.M. Potent Inhibitory Activities of the Adenosine Analogue Cordycepin on SARS-CoV-2 Replication. ACS Omega 2022, 7, 2960–2969. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, D.; Gao, X.; Wang, X.; Zhang, L. 2′- and 3′-Ribose Modifications of Nucleotide Analogues Establish the Structural Basis to Inhibit the Viral Replication of SARS-CoV-2. J. Phys. Chem. Lett. 2022, 13, 4111–4118. [Google Scholar] [CrossRef] [PubMed]
- Glab-Ampai, K.; Kaewchim, K.; Thavorasak, T.; Saenlom, T.; Thepsawat, W.; Mahasongkram, K.; Thueng-In, K.; Sookrung, N.; Chaicumpa, W.; Chulanetra, M. Targeting Emerging RNA Viruses by Engineered Human Super-antibody to Hepatitis C Virus RNA-Dependent RNA Polymerase. Front. Microbiol. 2022, 13, 926929. [Google Scholar] [CrossRef]
- Comunale, B.A.; Engineer, L.; Jiang, Y.; Andrews, J.C.; Liu, Q.; Ji, L.; Yurkovich, J.T.; Comunale, R.A.; Xie, Q. Poliovirus Vaccination Induces a Humoral Immune Response That Cross Reacts With SARS-CoV-2. Front. Med. 2021, 8, 1285. [Google Scholar] [CrossRef] [PubMed]
- Tenforde, M.W.; Shapiro, A.E.; Rouse, B.; Jarvis, J.N.; Li, T.; Eshun-Wilson, I.; Ford, N. Treatment for HIV-associated cryptococcal meningitis. Cochrane Database Syst. Rev. 2018, 7, CD005647. [Google Scholar] [CrossRef]
- Konopka, K.; Guo, L.S.; Düzgüneş, N. Anti-HIV activity of amphotericin B-cholesteryl sulfate colloidal dispersion in vitro. Antivir. Res. 1999, 42, 197–209. [Google Scholar] [CrossRef]
- Al-Khikani, F.H.O. Amphotericin B as antiviral drug: Possible efficacy against COVID-19. Ann. Thorac. Med. 2020, 15, 118–124. [Google Scholar] [CrossRef]
- Park, J.H.; Park, H.S.; Nah, H.J.; Kang, S.H.; Choi, S.S.; Kim, E.S. Streptomyces BAC Cloning of a Large-Sized Biosynthetic Gene Clus ter of NPP B1, a Potential SARS-CoV-2 RdRp Inhibitor. J. Microbiol. Biotechnol. 2022, 32, 911–917. [Google Scholar] [CrossRef]
- Dörner, T. Therapy: Hydroxychloroquine in SLE: Old drug, new perspectives. Nat. Rev. Rheumatol. 2010, 6, 10–11, Erratum in Nat. Rev. Rheumatol. 2010, 6, 118. [Google Scholar] [CrossRef] [PubMed]
- Nimgampalle, M.; Devanathan, V.; Saxena, A. Screening of Chloroquine, Hydroxychloroquine and its derivatives for their binding affinity to multiple SARS-CoV-2 protein drug targets. J. Biomol. Struct. Dyn. 2021, 39, 4949–4961. [Google Scholar] [CrossRef]
- Maisonnasse, P.; Guedj, J.; Contreras, V.; Behillil, S.; Solas, C.; Marlin, R.; Naninck, T.; Pizzorno, A.; Lemaitre, J.; Gonçalves, A.; et al. Hydroxychloroquine use against SARS-CoV-2 infection in non-human primates. Nature 2020, 585, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Ye, F.; Zhang, M.; Cui, C.; Huang, B.; Niu, P.; Liu, X.; Zhao, L.; Dong, E.; Song, C.; et al. In Vitro Antiviral Activity and Projection of Optimized Dosing Design of Hydroxychloroquine for the Treatment of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Clin. Infect. Dis. 2020, 71, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Gautret, P.; Lagier, J.C.; Parola, P.; Hoang, V.T.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020, 56, 105949. [Google Scholar] [CrossRef] [PubMed]
- Chistyakov, V.A.; Smirnova, Y.O.; Prazdnova, E.V.; Soldatov, A.V. Possible mechanisms of fullerene C60 antioxidant action. Biomed. Res. Int. 2013, 2013, 821498. [Google Scholar] [CrossRef] [PubMed]
- Bakry, R.; Vallant, R.M.; Najam-ul-Haq, M.; Rainer, M.; Szabo, Z.; Huck, C.W.; Bonn, G.K. Medicinal applications of fullerenes. Int. J. Nanomed. 2007, 2, 639–649. [Google Scholar]
- Hurmach, V.V.; Platonov, M.O.; Prylutska, S.V.; Scharff, P.; Prylutskyy, Y.I.; Ritter, U. C60 fullerene against SARS-CoV-2 corona virus: An in silico insight. Sci. Rep. 2021, 11, 17748. [Google Scholar] [CrossRef]
- Hurmach, V.; Platonov, M.; Prylutska, S.; Scharff, P.; Prylutskyy, Y.I.; Ritter, U. Anticoronavirus Activity of Water-Soluble Pristine C60 Fullerenes: In Vitro and In Silico Screenings. Adv. Exp. Med. Biol. 2021, 1352, 159–172. [Google Scholar] [CrossRef]
- Guardia, T.; Rotelli, A.E.; Juarez, A.O.; Pelzer, L.E. Anti-inflammatory properties of plant flavonoids. Effects of rutin, quercetin and hesperidin on adjuvant arthritis in rat. Farmaco 2001, 56, 683–687. [Google Scholar] [CrossRef]
- Goc, A.; Rath, M.; Niedzwiecki, A. Composition of naturally occurring compounds decreases activity of Omicron and SARS-CoV-2 RdRp complex. Eur. J. Microbiol. Immunol. 2022, 12, 39–45. [Google Scholar] [CrossRef]
- Munafò, F.; Donati, E.; Brindani, N.; Ottonello, G.; Armirotti, A.; De Vivo, M. Quercetin and luteolin are single-digit micromolar inhibitors of the SARS-CoV-2 RNA-dependent RNA polymerase. Sci. Rep. 2022, 12, 10571. [Google Scholar] [CrossRef]
- Rabie, A.M. Discovery of Taroxaz-104: The first potent antidote of SARS-CoV-2 VOC-202012/01 strain. J. Mol. Struct. 2021, 1246, 131106. [Google Scholar] [CrossRef]
- Wiedemar, N.; Hauser, D.A.; Mäser, P. 100 Years of Suramin. Antimicrob. Agents Chemother. 2020, 64, e01168-19. [Google Scholar] [CrossRef]
- Yin, W.; Luan, X.; Li, Z.; Zhou, Z.; Wang, Q.; Gao, M.; Wang, X.; Zhou, F.; Shi, J.; You, E.; et al. Structural basis for inhibition of the SARS-CoV-2 RNA polymerase by suramin. Nat. Struct. Mol. Biol. Mar. 2021, 28, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Squibb, B.-M. A Follow-up Evaluation for Safety in Subjects Who Participated in a Phase 1 Study With BMS-986094 (INX-08189). National Institutes of Health, U.S. National Library of Medicine. Updated 15 August 2014. Available online: https://clinicaltrials.gov/ct2/show/NCT01732848 (accessed on 30 October 2023).
- Pawlotsky, J.M. New antiviral agents for hepatitis C. F1000 Biol. Rep. 2012, 4, 5. [Google Scholar] [CrossRef]
- Delang, L.; Neyts, J.; Vliegen, I.; Abrignani, S.; Neddermann, P.; De Francesco, R. Hepatitis C virus-specific directly acting antiviral drugs. Curr. Top. Microbiol. Immunol. 2013, 369, 289–320. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Guardeño, J.M.; Ortega-Prieto, A.M.; Menendez Moreno, B.; Maguire, T.J.A.; Richardson, A.; Diaz-Hernandez, J.I.; Perez, J.D.; Zuckerman, M.; Playa, A.M.; Deline, C.C.; et al. Drug repurposing based on a quantum-inspired method versus classical fingerprinting uncovers potential antivirals against SARS-CoV-2. PLoS Comput. Biol. 2022, 18, e1010330. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Allergy and Infectious Diseases (NIAID). GS-5734 to Assess the Antiviral Activity, Longer-Term Clear ance of Ebola Virus, and Safety in Male Ebola Survivors With Evidence of Ebola Virus Persistence in Semen. National Insti tutes of Health, U.S. National Library of Medicine. Updated 15 February 2022. Available online: https://clinicaltrials.gov/ct2/show/NCT02818582 (accessed on 30 October 2023).
- Higgs, E.S.; Gayedyu-Dennis, D.; Fischer Ii, W.A.; Nason, M.; Reilly, C.; Beavogui, A.H.; Aboulhab, J.; Nordwall, J.; Lobbo, P.; Wachekwa, I.; et al. PREVAIL IV: A Randomized, Double-Blind, 2-Phase, Phase 2 Trial of Remdesivir vs Placebo for Reduction of Ebola Virus RNA in the Semen of Male Survivors. Clin. Infect. Dis. 2021, 73, 1849–1856. [Google Scholar] [CrossRef]
- Pardo, J.; Shukla, A.M.; Chamarthi, G.; Gupte, A. The journey of remdesivir: From Ebola to COVID-19. Drugs Context. 2020, 9, 2020-4-14. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Ledipasvir/Sofosbuvir: A Review in Chronic Hepatitis C. Drugs 2018, 78, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.M.; Polepally, A.R.; Khatri, A.; Awni, W.M.; Dutta, S. Clinical Pharmacokinetics of Paritaprevir. Clin. Pharmacokinet. 2017, 56, 1125–1137. [Google Scholar] [CrossRef]
- Pirzada, R.H.; Haseeb, M.; Batool, M.; Kim, M.; Choi, S. Remdesivir and Ledipasvir among the FDA-Approved Antiviral Drugs Have Potential to Inhibit SARS-CoV-2 Replication. Cells 2021, 10, 1052. [Google Scholar] [CrossRef] [PubMed]
- Vangeel, L.; Chiu, W.; De Jonghe, S.; Maes, P.; Slechten, B.; Raymenants, J.; André, E.; Leyssen, P.; Neyts, J.; Jochmans, D. Remdesivir, Molnupiravir and Nirmatrelvir remain active against SARS-CoV-2 Omi cron and other variants of concern. Antiviral Res. 2022, 198, 105252. [Google Scholar] [CrossRef]
- Choudhury, S.; Moulick, D.; Saikia, P.; Mazumder, M.K. Evaluating the potential of different inhibitors on RNA-dependent RNA polymerase of severe acute respiratory syndrome coronavirus 2: A molecular modeling approach. Med. J. Armed Forces India 2021, 77 (Suppl. S2), S373–S378. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, R. Structural Basis of the Potential Binding Mechanism of Remdesivir to SARS-CoV-2 RNA-Dependent RNA Polymerase. J. Phys. Chem. B 2020, 124, 6955–6962. [Google Scholar] [CrossRef]
- Koulgi, S.; Jani, V.; Uppuladinne, M.V.N.; Sonavane, U.; Joshi, R. Remdesivir-bound and ligand-free simulations reveal the proba ble mechanism of inhibiting the RNA dependent RNA polymerase of severe acute respiratory syndrome coronavirus 2. RSC Adv. 2020, 10, 26792–26803. [Google Scholar] [CrossRef]
- Nguyen, H.L.; Thai, N.Q.; Truong, D.T.; Li, M.S. Remdesivir Strongly Binds to Both RNA-Dependent RNA Polymerase and Main Protease of SARS-CoV-2: Evidence from Molecular Simulations. J. Phys. Chem. B 2020, 124, 11337–11348. [Google Scholar] [CrossRef]
- Aranda, J.; Wieczór, M.; Terrazas, M.; Brun-Heath, I.; Orozco, M. Mechanism of reaction of RNA-dependent RNA polymerase from SARS-CoV-2. Chem. Catal. 2022, 2, 1084–1099. [Google Scholar] [CrossRef]
- Tchesnokov, E.P.; Gordon, C.J.; Woolner, E.; Kocinkova, D.; Perry, J.K.; Feng, J.Y.; Porter, D.P.; Götte, M. Template-dependent inhibition of coronavirus RNA-dependent RNA polymer ase by remdesivir reveals a second mechanism of action. J. Biol. Chem. 2020, 295, 16156–16165. [Google Scholar] [CrossRef] [PubMed]
- Stevens, L.J.; Pruijssers, A.J.; Lee, H.W.; Gordon, C.J.; Tchesnokov, E.P.; Gribble, J.; George, A.S.; Hughes, T.M.; Lu, X.; Li, J.; et al. Mutations in the SARS-CoV-2 RNA-dependent RNA polymerase confer resistance to remdesivir by distinct mechanisms. Sci. Transl. Med. 2022, 14, eabo0718. [Google Scholar] [CrossRef] [PubMed]
- Rabie, A.M. Efficacious Preclinical Repurposing of the Nucleoside Analogue Didanosine against COVID-19 Polymerase and Exonuclease. ACS Omega 2022, 7, 21385–21396. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, W.; Wen, Z.; Wang, C.; Liu, W.; Zhang, Y.; Liu, J.; Ding, T.; Shuai, L.; Zhong, G.; et al. Gossypol Broadly Inhibits Coronaviruses by Targeting RNA-Dependent RNA Polymerases. Adv. Sci. 2022, 9, e2203499. [Google Scholar] [CrossRef] [PubMed]
- Jockusch, S.; Tao, C.; Li, X.; Chien, M.; Kumar, S.; Morozova, I.; Kalachikov, S.; Russo, J.J.; Ju, J. Sofosbuvir terminated RNA is more resistant to SARS-CoV-2 proofreader than RNA termi nated by Remdesivir. Sci. Rep. 2020, 10, 16577. [Google Scholar] [CrossRef] [PubMed]
- Toots, M.; Yoon, J.J.; Cox, R.M.; Hart, M.; Sticher, Z.M.; Makhsous, N.; Plesker, R.; Barrena, A.H.; Reddy, P.G.; Mitchell, D.G.; et al. Characterization of orally efficacious influenza drug with high resistance barrier in ferrets and human airway epithelia. Sci. Transl. Med. 2019, 11, eaax5866. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Tchesnokov, E.P.; Schinazi, R.F.; Götte, M. Molnupiravir promotes SARS-CoV-2 mutagenesis via the RNA template. J. Biol. Chem. 2021, 297, 100770. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Zhou, S.; Graham, R.L.; Pruijssers, A.J.; Agostini, M.L.; Leist, S.R.; Schäfer, A.; Dinnon III, K.H.; Stevens, L.J.; et al. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci. Transl. Med. 2020, 12, eabb5883. [Google Scholar] [CrossRef]
- Sourimant, J.; Lieber, C.M.; Aggarwal, M.; Cox, R.M.; Wolf, J.D.; Yoon, J.J.; Toots, M.; Ye, C.; Sticher, Z.; Kolykhalov, A.A.; et al. 4′-Fluorouridine is an oral antiviral that blocks respiratory syncytial virus and SARS-CoV-2 replication. Science 2022, 375, 161–167. [Google Scholar] [CrossRef]
- Reynard, O.; Nguyen, X.N.; Alazard-Dany, N.; Barateau, V.; Cimarelli, A.; Volchkov, V.E. Identification of a New Ribonucleoside Inhibitor of Ebola Virus Replication. Viruses 2015, 7, 6233–6240, Erratum in Viruses 2016, 8, 137. [Google Scholar] [CrossRef]
- Agostini, M.L.; Pruijssers, A.J.; Chappell, J.D.; Gribble, J.; Lu, X.; Andres, E.L.; Bluemling, G.R.; Lockwood, M.A.; Sheahan, T.P.; Sims, A.C.; et al. Small-Molecule Antiviral β-d-N4-Hydroxycytidine Inhibits a Proofreading-Intact Coronavirus with a High Genetic Barrier to Resistance. J. Virol. 2019, 93, e01348-19. [Google Scholar] [CrossRef] [PubMed]
- Khoo, S.H.; FitzGerald, R.; Saunders, G.; Middleton, C.; Ahmad, S.; Edwards, C.J.; Hadjiyiannakis, D.; Walker, L.; Lyon, R.; Shaw, V.; et al. Molnupiravir versus placebo in unvaccinated and vaccinated patients with early SARS-CoV-2 infection in the UK (AGILE CST-2): A randomised, placebo-controlled, double-blind, phase 2 trial. Lancet Infect. Dis. 2023, 23, 183–195, Erratum in Lancet Infect. Dis. 2023, 23, e1. [Google Scholar] [CrossRef] [PubMed]
- Ricci, F.; Tedeschi, A.; Morra, E.; Montillo, M. Fludarabine in the treatment of chronic lymphocytic leukemia: A review. Ther. Clin. Risk Manag. 2009, 5, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, H.; Kantarjian, H.; Ohanian, M.; Jabbour, E. The role of clofarabine in acute myeloid leukemia. Leuk. Lymphoma 2013, 54, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Coull, J.J.; Drusano, G.L.; Margolis, D.M. Dose proportional inhibition of HIV-1 replication by mycophenolic acid and synergistic inhibition in combination with abacavir, didanosine, and tenofovir. Antiviral Res. 2002, 55, 41–52. [Google Scholar] [CrossRef]
- Balzarini, J.; Robins, M.J.; Zou, R.M.; Herdewijn, P.; De Clercq, E. The 2′,3′-dideoxyriboside of 2,6-diaminopurine and its 2′,3′-dide hydro derivative inhibit the deamination of 2′,3′-dideoxyadenosine, an inhibitor of human immunodeficiency virus (HIV) replication. Biochem. Biophys. Res. Commun. 1987, 145, 277–283. [Google Scholar] [CrossRef]
- Suzuki, M.; Okuda, T.; Shiraki, K. Synergistic antiviral activity of acyclovir and vidarabine against herpes simplex virus types 1 and 2 and varicella-zoster virus. Antiviral Res. 2006, 72, 157–161. [Google Scholar] [CrossRef]
- Sirijatuphat, R.; Manosuthi, W.; Niyomnaitham, S.; Owen, A.; Kradangna Copeland, K.; Charoenpong, L.; Rattanasompattikul, M.; Mahasirimongkol, S.; Wichukchinda, N.; Chokephaibulkit, K. Early treatment of Favipiravir in COVID-19 patients without pneumo nia: A multicentre, open-labelled, randomized control study. Emerg. Microbes Infect. 2022, 11, 2197–2206. [Google Scholar] [CrossRef]
- Udwadia, Z.F.; Singh, P.; Barkate, H.; Patil, S.; Rangwala, S.; Pendse, A.; Kadam, J.; Wu, W.; Caracta, C.F.; Tandon, M. Efficacy and safety of favipiravir, an oral RNA-dependent RNA polymerase inhibitor, in mild-to-moderate COVID-19: A randomized, comparative, open-label, multicenter, phase 3 clinical trial. Int. J. Infect. Dis. 2021, 103, 62–71. [Google Scholar] [CrossRef]
- Shinkai, M.; Tsushima, K.; Tanaka, S.; Hagiwara, E.; Tarumoto, N.; Kawada, I.; Hirai, Y.; Fujiwara, S.; Komase, Y.; Saraya, T.; et al. Efficacy and Safety of Favipiravir in Moderate COVID-19 Pneumonia Patients with out Oxygen Therapy: A Randomized, Phase III Clinical Trial. Infect. Dis. Ther. 2021, 10, 2489–2509. [Google Scholar] [CrossRef]
- National Institute of Allergy and Infectious Diseases (NIAID). GS-7977 With Ribavirin for Hepatitis C (SPARE). National Institutes of Health, U.S. National Library of Medicine. Updated 26 September 2014. Available online: https://clinicaltrials.gov/ct2/show/NCT01441180 (accessed on 30 October 2023).
- Sacramento, C.Q.; Fintelman-Rodrigues, N.; Temerozo, J.R.; Dias Da Silva, A.d.P.; Da Silva Gomes Dias, S.; Dos Santos da Silva, C.; Ferreira, A.C.; Mattos, M.; Pão, C.R.R.; De Freitas, C.S.; et al. In vitro antiviral activity of the anti-HCV drugs daclatasvir and sofosbuvir against SARS-CoV-2, the aetiological agent of COVID-19. J. Antimicrob. Chemother. 2021, 76, 1874–1885. [Google Scholar] [CrossRef]
- Yuan, C.; Goonetilleke, E.C.; Unarta, I.C.; Huang, X. Incorporation efficiency and inhibition mechanism of 2′-substituted nucleo tide analogs against SARS-CoV-2 RNA-dependent RNA polymerase. Phys. Chem. Chem. Phys. 2021, 23, 20117–20128. [Google Scholar] [CrossRef]
- Bhatia, H.K.; Singh, H.; Grewal, N.; Natt, N.K. Sofosbuvir: A novel treatment option for chronic hepatitis C infection. J. Pharmacol. Pharmacother. 2014, 5, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Toschi, L.; Finocchiaro, G.; Bartolini, S.; Gioia, V.; Cappuzzo, F. Role of gemcitabine in cancer therapy. Future Oncol. 2005, 1, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Regal, R.E.; Mohammad, R.A. Daclatasvir: A NS5A Replication Complex Inhibitor for Hepatitis C Infection. Ann. Pharmacother. 2016, 50, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Stedman, C. Sofosbuvir, a NS5B polymerase inhibitor in the treatment of hepatitis C: A review of its clinical potential. Ther. Adv. Gastroenterol. 2014, 7, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Keating, G.M. Sofosbuvir: A review of its use in patients with chronic hepatitis C. Drugs 2014, 74, 1127–1146. [Google Scholar] [CrossRef]
- Novikov, M.S.; Ivanova, O.N.; Ivanov, A.V.; Ozerov, A.A.; Valuev-Elliston, V.T.; Temburnikar, K.; Gurskaya, G.V.; Kochetkov, S.N.; Pannecouque, C.; Balzarini, J.; et al. 1-[2-(2-Benzoyl- and 2-benzylphenoxy)ethyl]uracils as potent anti-HIV-1 agents. Bioorg. Med. Chem. 2011, 19, 5794–5802. [Google Scholar] [CrossRef]
- Magri, A.; Ozerov, A.A.; Tunitskaya, V.L.; Valuev-Elliston, V.T.; Wahid, A.; Pirisi, M.; Simmonds, P.; Ivanov, A.V.; Novikov, M.S.; Patel, A.H. Exploration of acetanilide derivatives of 1-(ω-phenoxyalkyl)uracils as novel in hibitors of Hepatitis C Virus replication. Sci Rep. 2016, 6, 29487, Erratum in Sci. Rep. 2016, 6, 31529. [Google Scholar] [CrossRef]
- Kezin, V.A.; Matyugina, E.S.; Novikov, M.S.; Chizhov, A.O.; Snoeck, R.; Andrei, G.; Kochetkov, S.N.; Khandazhinskaya, A.L. New Derivatives of 5-Substituted Uracils: Potential Agents with a Wide Spec trum of Biological Activity. Molecules 2022, 27, 2866. [Google Scholar] [CrossRef]
- Siniavin, A.E.; Novikov, M.S.; Gushchin, V.A.; Terechov, A.A.; Ivanov, I.A.; Paramonova, M.P.; Gureeva, E.S.; Russu, L.I.; Kuznetsova, N.A.; Shidlovskaya, E.V.; et al. Antiviral Activity of N(1),N(3)- Disubstituted Uracil Derivatives against SARS-CoV-2 Variants of Concern. Int. J. Mol. Sci. 2022, 23, 10171. [Google Scholar] [CrossRef] [PubMed]
- Angeli, F.; Spanevello, A.; Reboldi, G.; Visca, D.; Verdecchia, P. SARS-CoV-2 vaccines: Lights and shadows. Eur. J. Intern. Med. 2021, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Abbott, T.R.; Dhamdhere, G.; Liu, Y.; Lin, X.; Goudy, L.; Zeng, L.; Chemparathy, A.; Chmura, S.; Heaton, N.S.; Debs, R.; et al. Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influ enza. Cell 2020, 181, 865–876.e12. [Google Scholar] [CrossRef]
- Narayanan, N.; Nair, D.T. Vitamin B12 may inhibit RNA-dependent-RNA polymerase activity of nsp12 from the SARS-CoV-2 virus. IUBMB Life 2020, 72, 2112–2120. [Google Scholar] [CrossRef] [PubMed]
- Bushunow, P.; Reidenberg, M.M.; Wasenko, J.; Winfield, J.; Lorenzo, B.; Lemke, S.; Himpler, B.; Corona, R.; Coyle, T. Gossypol treatment of recurrent adult malignant gliomas. J. Neurooncol. 1999, 43, 79–86. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute (NCI). Gossypol in Treating Patients With Progressive or Recurrent Glioblastoma Multiforme. National Institutes of Health, U.S. National Library of Medicine. Updated 11 August 2017. Available online: https://clinicaltrials.gov/ct2/show/NCT00540722 (accessed on 30 October 2023).
- Wong, F.Y.; Liem, N.; Xie, C.; Yan, F.L.; Wong, W.C.; Wang, L.; Yong, W.P. Combination therapy with gossypol reveals synergism against gemcitabine resistance in can cer cells with high BCL-2 expression. PLoS ONE 2012, 7, e50786. [Google Scholar] [CrossRef]
- Lan, L.; Appelman, C.; Smith, A.R.; Yu, J.; Larsen, S.; Marquez, R.T.; Liu, H.; Wu, X.; Gao, P.; Roy, A.; et al. Natural product (-)-gossypol inhibits colon cancer cell growth by targeting RNA-bind ing protein Musashi-1. Mol. Oncol. 2015, 9, 1406–1420. [Google Scholar] [CrossRef]
- Pal, D.; Sahu, P.; Sethi, G.; Wallace, C.E.; Bishayee, A. Gossypol and Its Natural Derivatives: Multitargeted Phytochemicals as Po tential Drug Candidates for Oncologic Diseases. Pharmaceutics 2022, 14, 2624. [Google Scholar] [CrossRef]
- Zhong, X.; He, X.; Wang, Y.; Hu, Z.; Huang, H.; Zhao, S.; Wei, P.; Li, D. Warburg effect in colorectal cancer: The emerging roles in tumor microenvironment and thera peutic implications. J. Hematol. Oncol. 2022, 15, 160. [Google Scholar] [CrossRef]
- Agostinis, P.; Vantieghem, A.; Merlevede, W.; de Witte, P.A. Hypericin in cancer treatment: More light on the way. Int. J. Biochem. Cell Biol. 2002, 34, 221–241. [Google Scholar] [CrossRef]
- Dong, X.; Zeng, Y.; Zhang, Z.; Fu, J.; You, L.; He, Y.; Hao, Y.; Gu, Z.; Yu, Z.; Qu, C.; et al. Hypericin-mediated photodynamic therapy for the treatment of cancer: A review. J. Pharm. Pharmacol. 2021, 73, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.D.R.; Caetano, B.C.; de Almeida Filho, J.L.; Santa Cruz de Carvalho Martins, J.; Pacheco de Oliveira, M.G.; das Chagas Sousa, T.; Pareira Horta, M.A.; Mendonça Siqueira, M.; Hernandez Fernandez, J. Identification of Hypericin as a Candidate Repurposed Therapeutic Agent for COVID-19 and Its Potential Anti-SARS-CoV-2 Activity. Front. Microbiol. 2022, 13, 828984. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, A.K.; Kumar, R.; Gaguly, R.; Rana, H.K.; Pandey, P.K.; Sethi, G.; Bishayee, A.; Pandey, A.K. Corilagin in Cancer: A Critical Evaluation of Anticancer Activities and Molecular Mech anisms. Molecules 2019, 24, 3399. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zhang, G.; Li, Y.; Xu, J.; Yuan, J.; Zhang, B.; Hu, T.; Song, G. Corilagin inhibits breast cancer growth via reactive oxygen species-dependent apoptosis and autophagy. J. Cell Mol. Med. 2018, 22, 3795–3807. [Google Scholar] [CrossRef]
- Li, Q.; Yi, D.; Lei, X.; Zhao, J.; Zhang, Y.; Cui, X.; Xiao, X.; Jiao, T.; Dong, X.; Zhao, X.; et al. Corilagin inhibits SARS-CoV-2 replication by targeting viral RNA-dependent RNA polymerase. Acta Pharm. Sin. B 2021, 11, 1555–1567. [Google Scholar] [CrossRef]
- Negi, S.S.; Schein, C.H.; Braun, W. Regional and temporal coordinated mutation patterns in SARS-CoV-2 spike protein revealed by a clustering and network analysis. Sci. Rep. 2022, 12, 1128. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Drug/Compound Class Specific Agent (Reference) | RdRp Inhibition | RdRp Mechanism | SARS-CoV-2 Variants * | ||||||
|---|---|---|---|---|---|---|---|---|---|
| In Silico | In Vitro | In Vivo | Human Trial | In Silico | In Vitro | In Vivo | |||
| Adenosine-like fungal species | |||||||||
| Cordycepin (Bibi et al. (2022); Li et al. (2022); Rabie (2022a)) | 2 | 1 | 1 | 1 | |||||
| Antibodies | |||||||||
| Engineered super-Ab to hepatitis C RdRp (Glab-ampai et al. (2022)) | 1 | 1 | 1 | ||||||
| Poliovirus vaccine that induces Ab against poliovirus RdRp (Comunale et al. (2021)) | 1 | ||||||||
| Antifungal Antibiotics | |||||||||
| Biosynthetic gene cluster of NPP B1 (Park et al. (2022)) | 1 | ||||||||
| Antimalarials | |||||||||
| Chloroquine; derivatives (i.e., Hydroxychloroquine) (Gautret et al. (2021); Nimgampalle et al. (2021); Maisonnasse et al. (2020); Yao et al. (2020)) | 2 | 2 | 1 | 1 | |||||
| Antioxidants | |||||||||
| C60 Fullerene (Hurmach et al. (2021a); Hurmach et al. (2021b)) | 2 | 1 | |||||||
| Quercetin and Theaflavin (Goc et al. (2022)) | 1 | 1 | |||||||
| Quercetin and Luteolin (Munafò et al. (2022)) | 1 | 1 | |||||||
| Taroxaz-104 (Rabie (2021)) | 1 | 1 | 1 | ||||||
| Antiparasitics | |||||||||
| Suramin (Yin et al. (2021)) | 1 | 1 | |||||||
| Antivirals | |||||||||
| BMS-986094 (developed for hepatitis C) (Jimenez-Guardeño et al. (2022)) | 1 | 1 | 1 | ||||||
| Remdesivir (developed for Ebola virus) (Aranda et al. (2022); Choudhury et al. (2020); Koulgi et al. (2020); Nguyen et al. (2020); Pirzada et al. (2021); Stevens et al. (2022); Tchesnokov et al. (2020); Vangeel et al. (2022); Zhang & Zhou (2020)) | 1 | 2 | 7 | 2 | 2 | ||||
| Molnupiravir (developed for influenza virus); derivatives (i.e., 4′-fluorouridine, NHC) (Gordon et al. (2021); Khoo et al. (2023); Sheahan et al. (2020); Sourimant et al. (2022); Vangeel et al. (2022)) | 3 | 2 | 1 | 1 | 3 | ||||
| Nucleoside analogs (i.e., didanosine, fludarabine, vidarabine, favipiravir) (Li et al. (2022); Rabie (2022b); Shinkai et al. (2021); Sirijatuphat et al. (2022); Udwadia et al. (2021)) | 3 | 2 | 1 | 1 | |||||
| Sofosbuvir (developed for Hepatitis C) (Jockusch et al. (2020); Sacramento et al. (2021); Yuan et al. (2021)) | 2 | 2 | |||||||
| Uracil derivatives (used for HIV and Hepatitis C) (Siniavin et al. (2022)) | 1 | 1 | 1 | ||||||
| Vitamins, Natural Compounds, and Extracts | |||||||||
| Vitamin B12 (Jimenez-Guardeño et al. (2022); Narayanan & Nair (2020)) | 2 | 1 | 1 | ||||||
| Gossypol (used in cancer therapies) (Wang et al. (2022)) | 1 | 1 | 1 | 1 | |||||
| Hypericin (used in cancer therapies) (Matos et al. (2021)) | 1 | 1 | |||||||
| Corilagin (used in cancer therapies) (Li et al. (2021)) | 1 | 1 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comunale, B.A.; Larson, R.J.; Jackson-Ward, E.; Singh, A.; Koback, F.L.; Engineer, L.D. The Functional Implications of Broad Spectrum Bioactive Compounds Targeting RNA-Dependent RNA Polymerase (RdRp) in the Context of the COVID-19 Pandemic. Viruses 2023, 15, 2316. https://0-doi-org.brum.beds.ac.uk/10.3390/v15122316
Comunale BA, Larson RJ, Jackson-Ward E, Singh A, Koback FL, Engineer LD. The Functional Implications of Broad Spectrum Bioactive Compounds Targeting RNA-Dependent RNA Polymerase (RdRp) in the Context of the COVID-19 Pandemic. Viruses. 2023; 15(12):2316. https://0-doi-org.brum.beds.ac.uk/10.3390/v15122316
Chicago/Turabian StyleComunale, Brittany A., Robin J. Larson, Erin Jackson-Ward, Aditi Singh, Frances L. Koback, and Lilly D. Engineer. 2023. "The Functional Implications of Broad Spectrum Bioactive Compounds Targeting RNA-Dependent RNA Polymerase (RdRp) in the Context of the COVID-19 Pandemic" Viruses 15, no. 12: 2316. https://0-doi-org.brum.beds.ac.uk/10.3390/v15122316