
IFITMs from Naturally Infected Animal Species Exhibit Distinct Restriction Capacities against Toscana and Rift Valley Fever Viruses

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. IFITM Sequence Searches and Expression Plasmids
2.2. IFITM Phylogenetic Analysis and Protein Sequences Alignment
2.3. Cell Cultures
2.4. Immunofluorescence and Confocal Microscopy Experiments
2.5. Virus Stocks and Titrations
2.6. Transfection and Infection Assays
2.7. Western Blotting
2.8. Flow Cytometry
3. Results
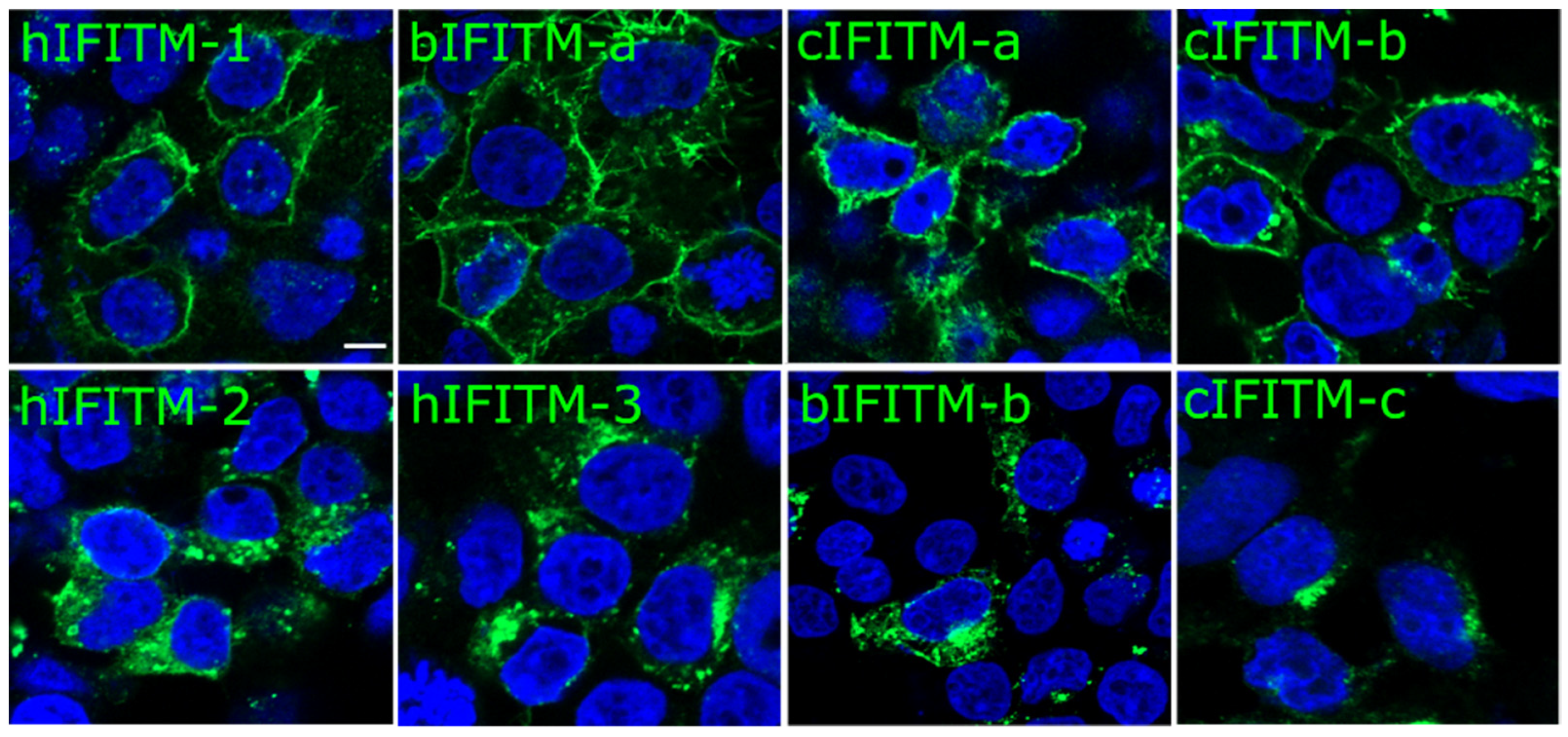
3.1. Characterization of Human, Bovine, and Camel IFITMs
3.2. IFITMs from Naturally Infected Species Differentially Restrict TOSV and RVFV
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cusi, M.G.; Savellini, G.G.; Zanelli, G. Toscana virus epidemiology: From Italy to beyond. Open Virol. J. 2010, 4, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Mari, J.M.; Palop-Borras, B.; Perez-Ruiz, M.; Sanbonmatsu-Gamez, S. Serosurvey study of Toscana virus in domestic animals, Granada, Spain. Vector Borne Zoonotic Dis. 2011, 11, 583–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartels, S.; de Boni, L.; Kretzschmar, H.A.; Heckmann, J.G. Lethal encephalitis caused by the Toscana virus in an elderly patient. J. Neurol. 2012, 259, 175–177. [Google Scholar] [CrossRef]
- Cusi, M.G.; Gandolfo, C.; Terrosi, C.; Gori Savellini, G.; Belmonte, G.; Miracco, C. Toscana virus infects dendritic and endothelial cells opening the way for the central nervous system. J. Neurovirol. 2015, 22, 307–315. [Google Scholar] [CrossRef]
- Vocale, C.; Bartoletti, M.; Rossini, G.; Macini, P.; Pascucci, M.G.; Mori, F.; Tampieri, A.; Lenzi, T.; Pavoni, M.; Giorgi, C.; et al. Toscana virus infections in northern Italy: Laboratory and clinical evaluation. Vector Borne Zoonotic Dis. 2012, 12, 526–529. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.R.; Coackley, W.; Roach, R.W.; Cowdy, N.R. Rift Valley fever in camels. J. Pathol. Bacteriol. 1963, 86, 229–231. [Google Scholar] [CrossRef]
- El-Harrak, M.; Martin-Folgar, R.; Llorente, F.; Fernandez-Pacheco, P.; Brun, A.; Figuerola, J.; Jimenez-Clavero, M.A. Rift Valley and West Nile virus antibodies in camels, North Africa. Emerg. Infect. Dis. 2011, 17, 2372–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegami, T.; Makino, S. The pathogenesis of Rift Valley fever. Viruses 2011, 3, 493–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linthicum, K.J.; Britch, S.C.; Anyamba, A. Rift Valley Fever: An Emerging Mosquito-Borne Disease. Annu. Rev. Entomol. 2016, 61, 395–415. [Google Scholar] [CrossRef]
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley fever virus (Bunyaviridae: Phlebovirus): An update on pathogenesis, molecular epidemiology, vectors, diagnostics and prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef]
- Laughlin, L.W.; Meegan, J.M.; Strausbaugh, L.J.; Morens, D.M.; Watten, R.H. Epidemic Rift Valley fever in Egypt: Observations of the spectrum of human illness. Trans. R. Soc. Trop. Med. Hyg. 1979, 73, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Madani, T.A.; Al-Mazrou, Y.Y.; Al-Jeffri, M.H.; Mishkhas, A.A.; Al-Rabeah, A.M.; Turkistani, A.M.; Al-Sayed, M.O.; Abodahish, A.A.; Khan, A.S.; Ksiazek, T.G.; et al. Rift Valley fever epidemic in Saudi Arabia: Epidemiological, clinical, and laboratory characteristics. Clin. Infect. Dis. 2003, 37, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, B.M.; Russell, D.; dos Santos, I.; Gear, J.H. Rift Valley fever in humans in South Africa. S. Afr. Med. J. 1980, 58, 803–806. [Google Scholar] [PubMed]
- Al-Hazmi, M.; Ayoola, E.A.; Abdurahman, M.; Banzal, S.; Ashraf, J.; El-Bushra, A.; Hazmi, A.; Abdullah, M.; Abbo, H.; Elamin, A.; et al. Epidemic Rift Valley fever in Saudi Arabia: A clinical study of severe illness in humans. Clin. Infect. Dis. 2003, 36, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shieh, W.J.; Paddock, C.D.; Lederman, E.; Rao, C.Y.; Gould, L.H.; Mohamed, M.; Mosha, F.; Mghamba, J.; Bloland, P.; Njenga, M.K.; et al. Pathologic studies on suspect animal and human cases of Rift Valley fever from an outbreak in Eastern Africa, 2006–2007. Am. J. Trop. Med. Hyg. 2010, 83 (Suppl. 2), 38–42. [Google Scholar] [CrossRef] [Green Version]
- Majdoul, S.; Compton, A.A. Lessons in self-defence: Inhibition of virus entry by intrinsic immunity. Nat. Rev. Immunol. 2022, 22, 339–352. [Google Scholar] [CrossRef]
- Zhao, X.; Li, J.; Winkler, C.A.; An, P.; Guo, J.T. IFITM Genes, Variants, and Their Roles in the Control and Pathogenesis of Viral Infections. Front. Microbiol. 2018, 9, 3228. [Google Scholar] [CrossRef] [Green Version]
- Tartour, K.; Appourchaux, R.; Gaillard, J.; Nguyen, X.N.; Durand, S.; Turpin, J.; Beaumont, E.; Roch, E.; Berger, G.; Mahieux, R.; et al. IFITM proteins are incorporated onto HIV-1 virion particles and negatively imprint their infectivity. Retrovirology 2014, 11, 103. [Google Scholar] [CrossRef] [Green Version]
- Tartour, K.; Nguyen, X.N.; Appourchaux, R.; Assil, S.; Barateau, V.; Bloyet, L.M.; Burlaud Gaillard, J.; Confort, M.P.; Escudero-Perez, B.; Gruffat, H.; et al. Interference with the production of infectious viral particles and bimodal inhibition of replication are broadly conserved antiviral properties of IFITMs. PLoS Pathog. 2017, 13, e1006610. [Google Scholar] [CrossRef] [Green Version]
- Chesarino, N.M.; McMichael, T.M.; Hach, J.C.; Yount, J.S. Phosphorylation of the antiviral protein interferon-inducible transmembrane protein 3 (IFITM3) dually regulates its endocytosis and ubiquitination. J. Biol. Chem. 2014, 289, 11986–11992. [Google Scholar] [CrossRef]
- Chesarino, N.M.; McMichael, T.M.; Yount, J.S. E3 Ubiquitin Ligase NEDD4 Promotes Influenza Virus Infection by Decreasing Levels of the Antiviral Protein IFITM3. PLoS Pathog. 2015, 11, e1005095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, R.; Xu, F.; Qian, J.; Yao, Y.; Miao, C.; Zheng, Y.M.; Liu, S.L.; Guo, F.; Geng, Y.; Qiao, W.; et al. Identification of an endocytic signal essential for the antiviral action of IFITM3. Cell Microbiol. 2014, 16, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Blyth, G.A.; Chan, W.F.; Webster, R.G.; Magor, K.E. Duck Interferon-Inducible Transmembrane Protein 3 Mediates Restriction of Influenza Viruses. J. Virol. 2016, 90, 103–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Ou, J.; Cai, S.; Lai, Z.; Zhong, L.; Yin, X.; Li, S. Canine Interferon-Inducible Transmembrane Protein Is a Host Restriction Factor That Potently Inhibits Replication of Emerging Canine Influenza Virus. Front. Immunol. 2021, 12, 710705. [Google Scholar] [CrossRef]
- Benfield, C.T.O.; Smith, S.E.; Wright, E.; Wash, R.S.; Ferrara, F.; Temperton, N.J.; Kellam, P. Bat and pig IFN-induced transmembrane protein 3 restrict cell entry by influenza virus and lyssaviruses. J. Gen. Virol. 2015, 96 Pt 5, 991–1005. [Google Scholar] [CrossRef] [Green Version]
- Everitt, A.R.; Clare, S.; Pertel, T.; John, S.P.; Wash, R.S.; Smith, S.E.; Chin, C.R.; Feeley, E.M.; Sims, J.S.; Adams, D.J.; et al. IFITM3 restricts the morbidity and mortality associated with influenza. Nature 2012, 484, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Gibson, M.S.; Wash, R.S.; Ferrara, F.; Wright, E.; Temperton, N.; Kellam, P.; Fife, M. Chicken interferon-inducible transmembrane protein 3 restricts influenza viruses and lyssaviruses in vitro. J. Virol. 2013, 87, 12957–12966. [Google Scholar] [CrossRef] [Green Version]
- Mudhasani, R.; Tran, J.P.; Retterer, C.; Radoshitzky, S.R.; Kota, K.P.; Altamura, L.A.; Smith, J.M.; Packard, B.Z.; Kuhn, J.H.; Costantino, J.; et al. IFITM-2 and IFITM-3 but not IFITM-1 restrict Rift Valley fever virus. J. Virol. 2013, 87, 8451–8464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marziali, F.; Delpeuch, M.; Kumar, A.; Appourchaux, R.; Dufloo, J.; Tartour, K.; Etienne, L.; Cimarelli, A. Functional Heterogeneity of Mammalian IFITM Proteins against HIV-1. J. Virol. 2021, 95, e0043921. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed] [Green Version]
- Lindenbach, B.D. Measuring HCV infectivity produced in cell culture and in vivo. Methods Mol. Biol. 2009, 510, 329–336. [Google Scholar] [PubMed]
- Alexander, A.J.T.; Confort, M.P.; Desloire, S.; Dunlop, J.I.; Kuchi, S.; Sreenu, V.B.; Mair, D.; Wilkie, G.S.; Da Silva Filipe, A.; Brennan, B.; et al. Development of a Reverse Genetics System for Toscana Virus (Lineage A). Viruses 2020, 12, 411. [Google Scholar] [CrossRef] [Green Version]
- Brennan, B.; Li, P.; Elliott, R.M. Generation and characterization of a recombinant Rift Valley fever virus expressing a V5 epitope-tagged RNA-dependent RNA polymerase. J. Gen. Virol. 2011, 92 Pt 12, 2906–2913. [Google Scholar] [PubMed] [Green Version]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, F.; Liu, F.; Cuconati, A.; Chang, J.; Block, T.M.; Guo, J.T. Interferon induction of IFITM proteins promotes infection by human coronavirus OC43. Proc. Natl. Acad. Sci. USA 2014, 111, 6756–6761. [Google Scholar] [CrossRef] [Green Version]
- Prelli Bozzo, C.; Nchioua, R.; Volcic, M.; Koepke, L.; Kruger, J.; Schutz, D.; Heller, S.; Sturzel, C.M.; Kmiec, D.; Conzelmann, C.; et al. IFITM proteins promote SARS-CoV-2 infection and are targets for virus inhibition in vitro. Nat. Commun. 2021, 12, 4584. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Confort, M.-P.; Duboeuf, M.; Thiesson, A.; Pons, L.; Marziali, F.; Desloire, S.; Ratinier, M.; Cimarelli, A.; Arnaud, F. IFITMs from Naturally Infected Animal Species Exhibit Distinct Restriction Capacities against Toscana and Rift Valley Fever Viruses. Viruses 2023, 15, 306. https://0-doi-org.brum.beds.ac.uk/10.3390/v15020306
Confort M-P, Duboeuf M, Thiesson A, Pons L, Marziali F, Desloire S, Ratinier M, Cimarelli A, Arnaud F. IFITMs from Naturally Infected Animal Species Exhibit Distinct Restriction Capacities against Toscana and Rift Valley Fever Viruses. Viruses. 2023; 15(2):306. https://0-doi-org.brum.beds.ac.uk/10.3390/v15020306
Chicago/Turabian StyleConfort, Marie-Pierre, Maëva Duboeuf, Adrien Thiesson, Léa Pons, Federico Marziali, Sophie Desloire, Maxime Ratinier, Andrea Cimarelli, and Frédérick Arnaud. 2023. "IFITMs from Naturally Infected Animal Species Exhibit Distinct Restriction Capacities against Toscana and Rift Valley Fever Viruses" Viruses 15, no. 2: 306. https://0-doi-org.brum.beds.ac.uk/10.3390/v15020306