
SARS-CoV-2 Vaccine-Induced T-Cell Response after Three Doses in People Living with HIV on Antiretroviral Therapy Compared to Seronegative Controls (CTN 328 COVAXHIV Study)

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
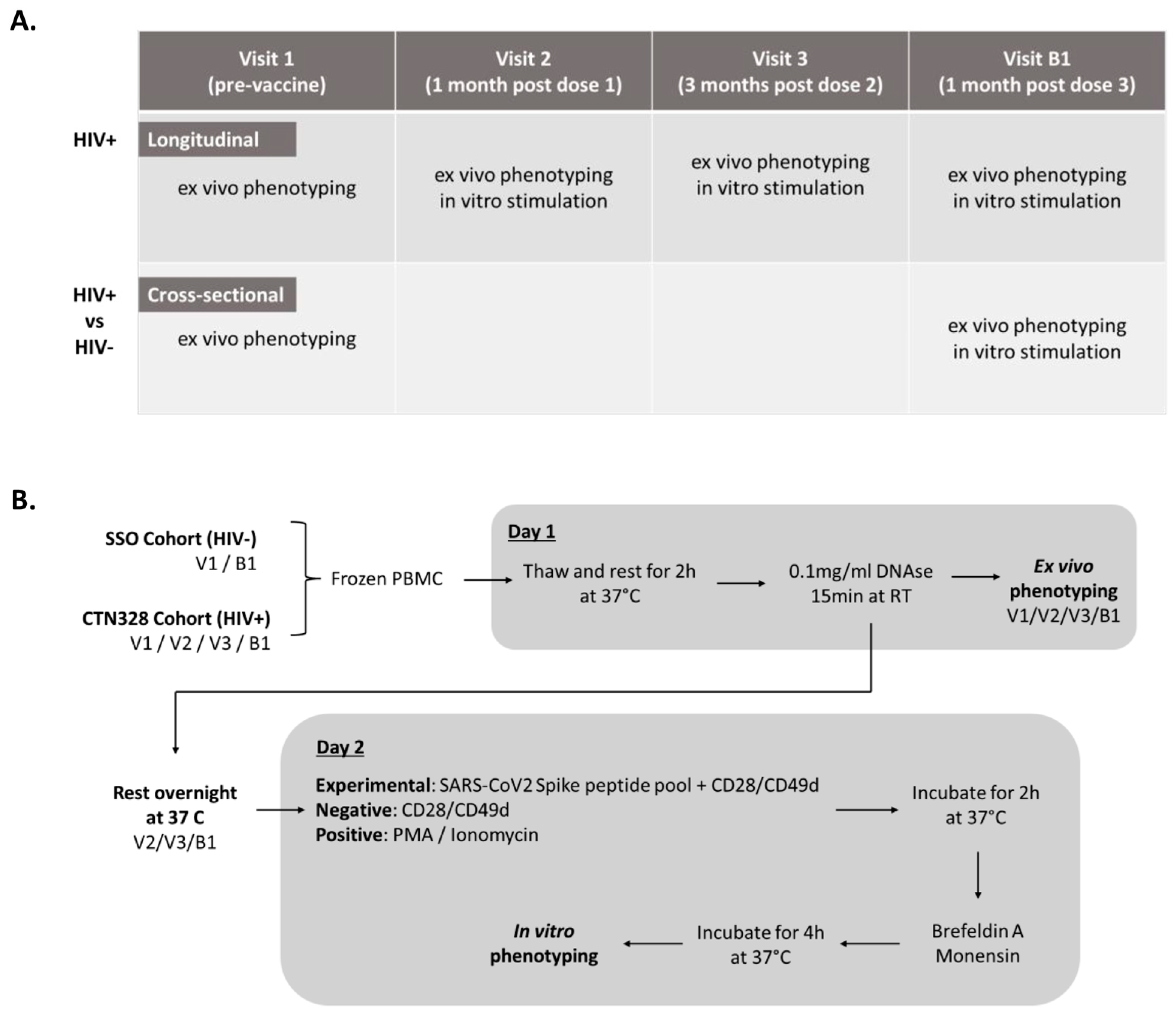
2.1. Study Population
2.2. Vaccine Doses and Study Visits
2.3. Ex Vivo Immunophenotyping of T-Cells
2.4. Intracellular Cytokine Production upon In Vitro SARS-CoV-2 Peptide Stimulation
2.5. Ethical Statement
2.6. Statistical Analyses
3. Results
3.1. Study Population
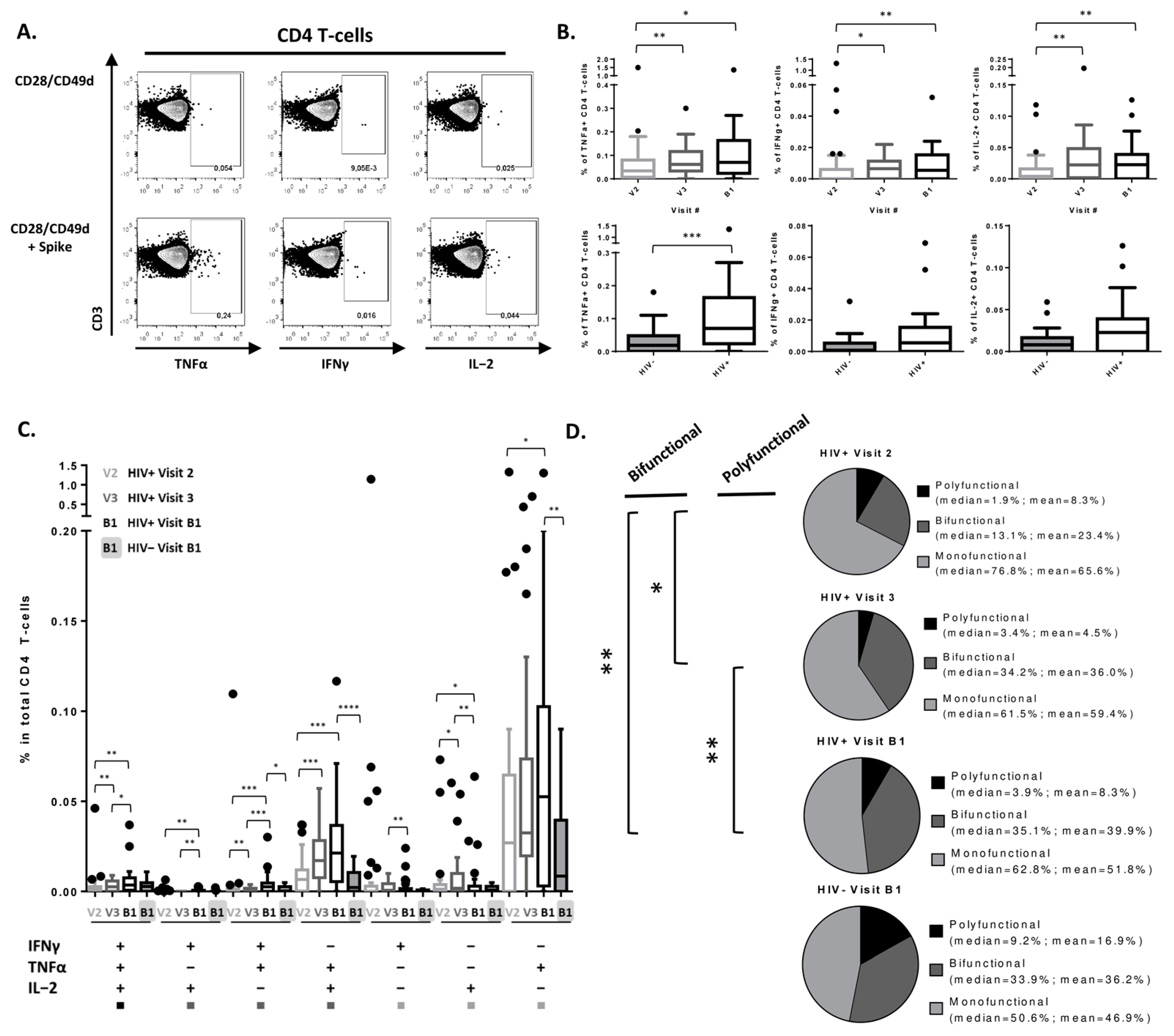
3.2. Peak SARS-CoV-2-Specific CD4 T-Cell Cytokine Secretion Response Achieved after 2nd Vaccine Dose in PLWH
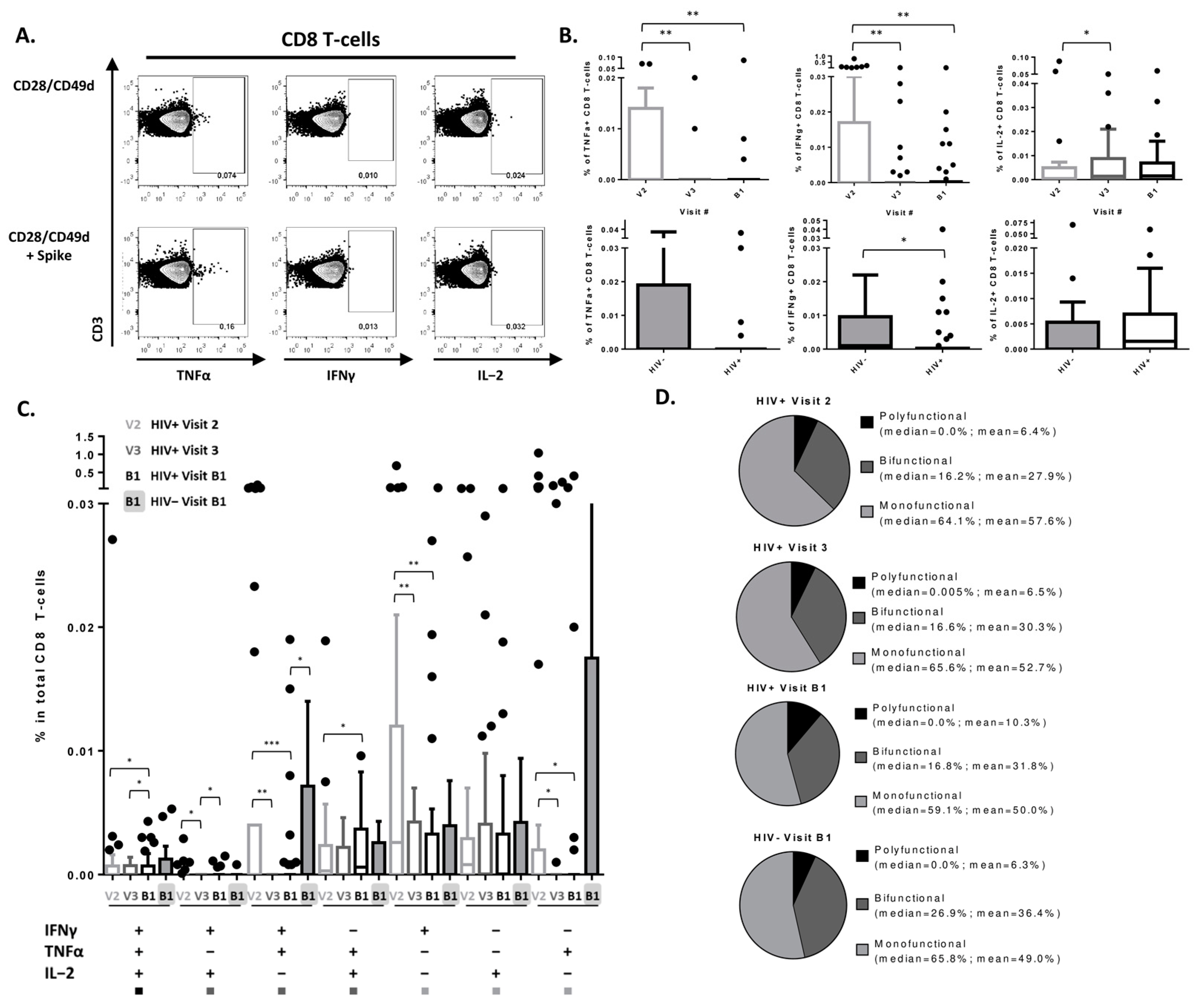
3.3. SARS-CoV-2 Vaccine Induces a Detectable CD8 T-Cell Response in PLWH
3.4. Changes in CD4 T-Cell Subsets following 3 Doses of SARS-CoV-2 Vaccination
3.5. Changes in CD8 T-Cell Subsets following 3 Doses of SARS-CoV-2 Vaccination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spinelli, M.A.; Jones, B.L.H.; Gandhi, M. COVID-19 Outcomes and Risk Factors Among People Living with HIV. Curr. HIV/AIDS Rep. 2022, 19, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Alexandrova, Y.; Costiniuk, C.T.; Jenabian, M.A. Pulmonary Immune Dysregulation and Viral Persistence during HIV Infection. Front Immunol. 2021, 12, 808722. [Google Scholar] [CrossRef]
- Tebas, P.; Frank, I.; Lewis, M.; Quinn, J.; Zifchak, L.; Thomas, A.; Kenney, T.; Kappes, R.; Wagner, W.; Maffei, K.; et al. Poor immunogenicity of the H1N1 2009 vaccine in well controlled HIV-infected individuals. AIDS 2010, 24, 2187–2192. [Google Scholar] [CrossRef]
- Farooq, P.D.; Sherman, K.E. Hepatitis B Vaccination and Waning Hepatitis B Immunity in Persons Living with HIV. Curr. HIV/AIDS Rep. 2019, 16, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Alrubayyi, A.; Gea-Mallorquí, E.; Touizer, E.; Hameiri-Bowen, D.; Kopycinski, J.; Charlton, B.; Fisher-Pearson, N.; Muir, L.; Rosa, A.; Roustan, C.; et al. Characterization of humoral and SARS-CoV-2 specific T cell responses in people living with HIV. Nat. Commun. 2021, 12, 5839. [Google Scholar] [CrossRef] [PubMed]
- Antinori, A.; Cicalini, S.; Meschi, S.; Bordoni, V.; Lorenzini, P.; Vergori, A.; Lanini, S.; De Pascale, L.; Matusali, G.; Mariotti, D.; et al. Humoral and Cellular Immune Response Elicited by mRNA Vaccination against Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in People Living with Human Immunodeficiency Virus Receiving Antiretroviral Therapy Based on Current CD4 T-Lymphocyte Count. Clin. Infect. Dis. 2022, 75, e552–e563. [Google Scholar]
- Coburn, S.B.; Humes, E.; Lang, R.; Stewart, C.; Hogan, B.C.; Gebo, K.A.; Napravnik, S.; Edwards, J.K.; Browne, L.E.; Park, L.S.; et al. Analysis of Postvaccination Breakthrough COVID-19 Infections among Adults with HIV in the United States. JAMA Netw. Open 2022, 5, e2215934. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Singer, J.; Langlois, M.A.; Kulic, I.; Needham, J.; Burchell, A.; Jenabian, M.A.; Walmsley, S.; Ostrowski, M.; Kovacs, C.; et al. CTN 328: Immunogenicity outcomes in people living with HIV in Canada following vaccination for COVID-19 (HIV-COV): Protocol for an observational cohort study. BMJ Open 2021, 11, e054208. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Singer, J.; Lee, T.; Langlois, M.-A.; Arnold, C.; Galipeau, Y.; Needham, J.; Kulic, I.; Jenabian, M.-A.; Burchell, A.N.; et al. COVID-19 vaccine immunogenicity in people living with HIV (CIHR Canadian HIV Trials Network 328). AIDS 2022, 37, F1–F10. [Google Scholar] [CrossRef]
- Cai, S.; Liao, G.; Yu, T.; Gao, Q.; Zou, L.; Zhang, H.; Xu, X.; Chen, J.; Lu, A.; Wu, Y.; et al. Immunogenicity and safety of an inactivated SARS-CoV-2 vaccine in people living with HIV: A cross-sectional study. J. Med. Virol. 2022, 94, 4224–4233. [Google Scholar] [CrossRef]
- Vergori, A.; Cozzi Lepri, A.; Cicalini, S.; Matusali, G.; Bordoni, V.; Lanini, S.; Meschi, S.; Iannazzo, R.; Mazzotta, V.; Colavita, F.; et al. Immunogenicity to COVID-19 mRNA vaccine third dose in people living with HIV. Nat. Commun. 2022, 13, 4922. [Google Scholar] [CrossRef]
- Collins, E.; Galipeau, Y.; Arnold, C.; Bosveld, C.; Heiskanen, A.; Keeshan, A.; Nakka, K.; Shir-Mohammadi, K.; St-Denis-Bissonnette, F.; Tamblyn, L.; et al. Cohort profile: Stop the Spread Ottawa (SSO)—A community-based prospective cohort study on antibody responses, antibody neutralisation efficiency and cellular immunity to SARS-CoV-2 infection and vaccination. BMJ Open 2022, 12, e062187. [Google Scholar] [CrossRef] [PubMed]
- Ao, L.; Lu, T.; Cao, Y.; Chen, Z.; Wang, Y.; Li, Z.; Ren, X.; Xu, P.; Peng, M.; Chen, M.; et al. Safety and immunogenicity of inactivated SARS-CoV-2 vaccines in people living with HIV. Emerg. Microbes Infect. 2022, 11, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Lamacchia, G.; Salvati, L.; Kiros, S.T.; Mazzoni, A.; Vanni, A.; Capone, M.; Carnasciali, A.; Farahvachi, P.; Lagi, F.; Di Lauria, N.; et al. Fourth Dose of mRNA COVID-19 Vaccine Transiently Reactivates Spike-Specific Immunological Memory in People Living with HIV (PLWH). Biomedicines 2022, 10, 3261. [Google Scholar] [CrossRef]
- Lang, R.; Humes, E.; Coburn, S.B.; Horberg, M.A.; Fathi, L.F.; Watson, E.; Jefferson, C.R.; Park, L.S.; Gordon, K.S.; Akgün, K.M.; et al. Analysis of Severe Illness after Postvaccination COVID-19 Breakthrough among Adults with and without HIV in the US. JAMA Netw. Open 2022, 5, e2236397. [Google Scholar] [CrossRef] [PubMed]
- De Pablo-Bernal, R.S.; Ruiz-Mateos, E.; Rosado, I.; Dominguez-Molina, B.; Alvarez-Ríos, A.I.; Carrillo-Vico, A.; De La Rosa, R.; Delgado, J.; Muñoz-Fernández, M.A.; Leal, M.; et al. TNF-α levels in HIV-infected patients after long-term suppressive cART persist as high as in elderly, HIV-uninfected subjects. J. Antimicrob. Chemother. 2014, 69, 3041–3046. [Google Scholar] [CrossRef] [Green Version]
- Collora, J.A.; Pinto-Santini, D.; Pasalar, S.; Ravindra, N.; Ganoza, C.; Lama, J.; Alfaro, R.; Chiarella, J.; Spudich, S.; van Dijk, D.; et al. Single-cell immune profiling reveals the impact of antiretroviral therapy on HIV-1-induced immune dysfunction, T cell clonal expansion, and HIV-1 persistence in vivo. bioRxiv 2021. [Google Scholar] [CrossRef]
- Osuji, F.N.; Onyenekwe, C.C.; Ahaneku, J.E.; Ukibe, N.R. The effects of highly active antiretroviral therapy on the serum levels of pro-inflammatory and anti-inflammatory cytokines in HIV infected subjects. J. Biomed. Sci. 2018, 25, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.L.; Shive, C.L.; Nguyen, T.P.; Younes, S.A.; Panigrahi, S.; Lederman, M.M. Cytokines and T-Cell Homeostasis in HIV Infection. J. Infect Dis. 2016, 214, S51–S57. [Google Scholar] [CrossRef] [Green Version]
- Day, C.L.; Kaufmann, D.E.; Kiepiela, P.; Brown, J.A.; Moodley, E.S.; Reddy, S.; Mackey, E.W.; Miller, J.D.; Leslie, A.J.; DePierres, C.; et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 2006, 443, 350–354. [Google Scholar] [CrossRef]
- Petrovas, C.; Casazza, J.P.; Brenchley, J.M.; Price, D.A.; Gostick, E.; Adams, W.C.; Precopio, M.L.; Schacker, T.; Roederer, M.; Douek, D.C.; et al. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. J. Exp. Med. 2006, 203, 2281–2292. [Google Scholar] [CrossRef]
- Thieme, C.J.; Anft, M.; Paniskaki, K.; Blazquez-Navarro, A.; Doevelaar, A.; Seibert, F.S.; Hoelzer, B.; Konik, M.J.; Brenner, T.; Tempfer, C.; et al. The SARS-CoV-2 T-cell immunity is directed against the spike, membrane, and nucleocapsid protein and associated with COVID 19 severity. medRxiv 2020. [Google Scholar] [CrossRef]
- Deming, M.E.; Lyke, K.E. A ‘mix and match’ approach to SARS-CoV-2 vaccination. Nat. Med. 2021, 27, 1510–1511. [Google Scholar] [CrossRef]
- Zhang, Z.; Mateus, J.; Coelho, C.H.; Dan, J.M.; Moderbacher, C.R.; Gálvez, R.I.; Cortes, F.H.; Grifoni, A.; Tarke, A.; Chang, J.; et al. Humoral and cellular immune memory to four COVID-19 vaccines. Cell 2022, 185, 2434–2451.e17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, Y.; Kang, Y. Naïve T cells may be key to the low mortality of children with COVID-19. J. Evid.-Based Med. 2022, 15, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.D.; Emerson, S.G.; Punt, J.; Goff, W.D. Decreased Naïve T-cell Production Leading to Cytokine Storm as Cause of Increased COVID-19 Severity with Comorbidities. Aging Dis. 2020, 11, 742–745. [Google Scholar] [CrossRef]
- Vitalle, J.; Perez-Gomez, A.; Ostos, F.J.; Gasca-Capote, C.; Jimenez-Leon, M.R.; Bachiller, S.; Rivas-Jeremias, I.; Silva-Sanchez, M.D.M.; Ruiz-Mateos, A.M.; Martin-Sanchez, M.A.; et al. Immune defects associated with lower SARS-CoV-2 BNT162b2 mRNA vaccine response in aged people. JCI Insight 2022, 7, 17. [Google Scholar] [CrossRef]
- Amu, S.; Lantto Graham, R.; Bekele, Y.; Nasi, A.; Bengtsson, C.; Rethi, B.; Sorial, S.; Meini, G.; Zazzi, M.; Hejdeman, B.; et al. Dysfunctional phenotypes of CD4+ and CD8+ T cells are comparable in patients initiating ART during early or chronic HIV-1 infection. Medicine 2016, 95, 23. [Google Scholar] [CrossRef] [Green Version]
- Mold, J.E.; Réu, P.; Olin, A.; Bernard, S.; Michaëlsson, J.; Rane, S.; Yates, A.; Khosravi, A.; Salehpour, M.; Possnert, G.; et al. Cell generation dynamics underlying naive T-cell homeostasis in adult humans. PLoS Biol. 2019, 17, e3000383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, M.; Schulte, S.; Wildner, N.H.; Wittner, M.; Brehm, T.T.; Ramharter, M.; Woost, R.; Lohse, A.W.; Jacobs, T.; Schulze zur Wiesch, J. Analysis of Co-inhibitory Receptor Expression in COVID-19 Infection Compared to Acute Plasmodium falciparum Malaria: LAG-3 and TIM-3 Correlate with T Cell Activation and Course of Disease. Front. Immunol. 2020, 11, 1870. [Google Scholar] [CrossRef] [PubMed]
- Yoshie, O.; Matsushima, K. CCR4 and its ligands: From bench to bedside. Int. Immunol. 2015, 27, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spoerl, S.; Kremer, A.N.; Aigner, M.; Eisenhauer, N.; Koch, P.; Meretuk, L.; Löffler, P.; Tenbusch, M.; Maier, C.; Überla, K.; et al. Upregulation of CCR4 in activated CD8+ T cells indicates enhanced lung homing in patients with severe acute SARS-CoV-2 infection. Eur. J. Immunol. 2021, 51, 1436–1448. [Google Scholar] [CrossRef] [PubMed]
- Guerrera, G.; Picozza, M.; D’Orso, S.; Placido, R.; Pirronello, M.; Verdiani, A.; Termine, A.; Fabrizio, C.; Giannessi, F.; Sambucci, M.; et al. BNT162b2 vaccination induces durable SARS-CoV-2–specific T cells with a stem cell memory phenotype. Sci. Immunol. 2021, 6, eabl5344. [Google Scholar] [CrossRef] [PubMed]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.G.; Mendoza, A.; Hemmers, S.; Moltedo, B.; Niec, R.E.; Schizas, M.; Hoyos, B.E.; Putintseva, E.V.; Chaudhry, A.; Dikiy, S.; et al. Stability and function of regulatory T cells expressing the transcription factor T-bet. Nature 2017, 546, 421–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Y.; Fu, J.; Xing, S.; Fu, B.; Zhang, Z.; Shi, M.; Wang, X.; Zhang, J.; Jin, L.; Kang, F.; et al. The decrease of regulatory T cells correlates with excessive activation and apoptosis of CD8+ T cells in HIV-1-infected typical progressors, but not in long-term non-progressors. Immunology 2009, 128, e366–e375. [Google Scholar] [CrossRef]
- Machicote, A.; Belén, S.; Baz, P.; Billordo, L.A.; Fainboim, L. Human CD8. Front. Immunol. 2018, 9, 2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.K.; Kagari, T.; Clingan, J.M.; Matloubian, M. Expression of chemokine receptor CXCR3 on T cells affects the balance between effector and memory CD8 T-cell generation. Proc. Natl. Acad. Sci. USA 2011, 108, E118–E127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, M.; Dölen, Y.; van Dinther, E.; Vimeux, L.; Fallet, M.; Feuillet, V.; Figdor, C.G. Cross-talk between iNKT cells and CD8 T cells in the spleen requires the IL-4/CCL17 axis for the generation of short-lived effector cells. Proc. Natl. Acad. Sci. USA 2019, 116, 25816–25827. [Google Scholar] [CrossRef]
- Bayry, J. Regulatory T cells as adjuvant target for enhancing the viral disease vaccine efficacy. VirusDisease 2014, 25, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Jiang, X.; Dai, X.; Li, B. The Dynamic Role of FOXP3(+) Tregs and Their Potential Therapeutic Applications During SARS-CoV-2 Infection. Front. Immunol. 2022, 13, 916411. [Google Scholar] [CrossRef] [PubMed]
- Kidani, Y.; Nogami, W.; Yasumizu, Y.; Kawashima, A.; Tanaka, A.; Sonoda, Y.; Tona, Y.; Nashiki, K.; Matsumoto, R.; Hagiwara, M.; et al. CCR8-targeted specific depletion of clonally expanded Treg cells in tumor tissues evokes potent tumor immunity with long-lasting memory. Proc. Natl. Acad. Sci. USA 2022, 119, e2114282119. [Google Scholar] [CrossRef]
- Moore, A.C.; Gallimore, A.; Draper, S.J.; Watkins, K.R.; Gilbert, S.C.; Hill, A.V.S. Anti-CD25 Antibody Enhancement of Vaccine-Induced Immunogenicity: Increased Durable Cellular Immunity with Reduced Immunodominance1. J. Immunol. 2005, 175, 7264–7273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Haskó, G. CD39 and CD73 in immunity and inflammation. Trends Mol. Med. 2013, 19, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yero, A.; Farnos, O.; Rabezanahary, H.; Racine, G.; Estaquier, J.; Jenabian, M.A. Differential Dynamics of Regulatory T-Cell and Th17 Cell Balance in Mesenteric Lymph Nodes and Blood following Early Antiretroviral Initiation during Acute Simian Immunodeficiency Virus Infection. J. Virol. 2019, 93, e00371-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrière, M.; Lacabaratz, C.; Kök, A.; Benne, C.; Jenabian, M.A.; Casartelli, N.; Hüe, S.; Hocqueloux, L.; Lelièvre, J.D.; Lévy, Y. HIV “elite controllers” are characterized by a high frequency of memory CD8+ CD73+ T cells involved in the antigen-specific CD8+ T-cell response. J. Infect Dis. 2014, 209, 1321–1330. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Godec, J.; Wolski, D.; Adland, E.; Yates, K.; Pauken, K.E.; Cosgrove, C.; Ledderose, C.; Junger, W.G.; Robson, S.C.; et al. CD39 Expression Identifies Terminally Exhausted CD8+ T Cells. PLoS Pathog. 2015, 11, e1005177. [Google Scholar] [CrossRef] [Green Version]
- Bai, A.; Moss, A.; Rothweiler, S.; Serena Longhi, M.; Wu, Y.; Junger, W.G.; Robson, S.C. NADH oxidase-dependent CD39 expression by CD8+ T cells modulates interferon gamma responses via generation of adenosine. Nat. Commun. 2015, 6, 8819. [Google Scholar] [CrossRef] [Green Version]
- Steiner, K.; Waase, I.; Rau, T.; Dietrich, M.; Fleischer, B.; Bröker, B.M. Enhanced expression of CTLA-4 (CD152) on CD4+ T cells in HIV infection. Clin. Exp. Immunol. 1999, 115, 451–457. [Google Scholar] [CrossRef]
- Jubel, J.M.; Barbati, Z.R.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. The Role of PD-1 in Acute and Chronic Infection. Front. Immunol. 2020, 11, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusnadi, A.; Ramírez-Suástegui, C.; Fajardo, V.; Chee, S.J.; Meckiff, B.J.; Simon, H.; Pelosi, E.; Seumois, G.; Ay, F.; Vijayanand, P.; et al. Severely ill COVID-19 patients display impaired exhaustion features in SARS-CoV-2-reactive CD8(+) T cells. Sci. Immunol. 2021, 6, eabe4782. [Google Scholar] [CrossRef]
- Lin, H.-S.; Lin, X.-H.; Wang, J.-W.; Wen, D.-N.; Xiang, J.; Fan, Y.-Q.; Li, H.-D.; Wu, J.; Lin, Y.; Lin, Y.-L.; et al. Exhausting T Cells During HIV Infection May Improve the Prognosis of Patients with COVID-19. Front. Cell. Infect. Microbiol. 2021, 11, 564938. [Google Scholar] [CrossRef]
- Rha, M.-S.; Jeong, H.W.; Ko, J.-H.; Choi, S.J.; Seo, I.-H.; Lee, J.S.; Sa, M.; Kim, A.R.; Joo, E.-J.; Ahn, J.Y.; et al. PD-1-Expressing SARS-CoV-2-Specific CD8+ T Cells Are Not Exhausted, but Functional in Patients with COVID-19. Immunity 2021, 54, 44–52.e3. [Google Scholar] [CrossRef]
- Garcia-Bates, T.M.; Palma, M.L.; Shen, C.; Gambotto, A.; Macatangay, B.J.C.; Ferris, R.L.; Rinaldo, C.R.; Mailliard, R.B. Contrasting Roles of the PD-1 Signaling Pathway in Dendritic Cell-Mediated Induction and Regulation of HIV-1-Specific Effector T Cell Functions. J. Virol. 2019, 93, e02035-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, C.L.; Abrahams, D.A.; Bunjun, R.; Stone, L.; de Kock, M.; Walzl, G.; Wilkinson, R.J.; Burgers, W.A.; Hanekom, W.A. PD-1 Expression on Mycobacterium tuberculosis-Specific CD4 T Cells Is Associated with Bacterial Load in Human Tuberculosis. Front. Immunol. 2018, 9, 1995. [Google Scholar] [CrossRef] [PubMed]
- Breton, G.; Chomont, N.; Takata, H.; Fromentin, R.; Ahlers, J.; Filali-Mouhim, A.; Riou, C.; Boulassel, M.R.; Routy, J.P.; Yassine-Diab, B.; et al. Programmed death-1 is a marker for abnormal distribution of naive/memory T cell subsets in HIV-1 infection. J. Immunol. 2013, 191, 2194–2204. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.-M.; Bai, P.; He, W.; Wu, F.; Liu, X.-F.; Han, D.-M.; Liu, S.; Yang, J.-K. Gender Differences in Patients with COVID-19: Focus on Severity and Mortality. Front. Public Health 2020, 8, 152. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.J.; Swadling, L.; Gibbons, J.M.; Pade, C.; Jensen, M.P.; Diniz, M.O.; Schmidt, N.M.; Butler, D.K.; Amin, O.E.; Bailey, S.N.L.; et al. Discordant neutralizing antibody and T cell responses in asymptomatic and mild SARS-CoV-2 infection. Sci. Immunol. 2020, 5, eabf3698. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | PLWH, N = 38 1 | Uninfected, N = 24 1 | p-Value |
|---|---|---|---|
| Age, years, median (IQR) | 43 (36, 57) | 44 (38, 56) | 0.8 2 |
| Male sex, n (%) | 33 (87%) | 12 (50%) | 0.002 3 |
| Ethnicity, n (%) | 0.008 4 | ||
| Caucasian | 21 (55%) | 24 (100%) | |
| Black | 2 (5.3%) | 0 (0%) | |
| Chinese | 4 (11%) | 0 (0%) | |
| Filipino | 2 (5.3%) | 0 (0%) | |
| Japanese | 1 (2.6%) | 0 (0%) | |
| Latin American | 5 (13%) | 0 (0%) | |
| Other | 3 (7.9%) | 0 (0%) | |
| SARS-CoV2 vaccine type received, n (%) | 0.019 4 | ||
| 3 doses of mRNA (Pfizer or Moderna) | 38 (100%) | 20 (83%) | |
| 1st dose Adenovirus vector (Astrazeneca), 2nd/3d dose mRNA (Pfizer or Moderna) | 0 (0%) | 4 (17%) | |
| CD4 count (n = 35), cells/mm3, median (IQR) | 700 (480, 839) | N/A | N/A |
| CD4/CD8 ratio (n = 35) | 0.81 (0.59, 1.01) | ||
| CD4 nadir, cells/mm3, median (IQR) (n = 37) | 290 (167, 420) | ||
| Duration of HIV infection, years (IRQ) (n = 37) | 6.5 (5.0, 21.0) | ||
| Undetectable viral load for at least 6 months, n (%) | 30 (79%) | ||
| If detectable, highest viral load over past 6 months (n = 8), median (IQR) (copies/mL) | 182 (56, 2, 040) | ||
| ART regimen, n (%) | |||
| INSTI-based regimen | 28 (74%) | N/A | |
| NNRTI-based regimen | 5 (13%) | ||
| NRTI-based regimen | 2 (5.3%) | ||
| Other | 3 (7.9%) | ||
| Coinfections, n (%) | N/A | ||
| Hepatitis B virus | 0 (0%) | 0 (0%) | |
| Hepatitis C virus | 0 (0%) | 0 (0%) | |
| Characteristics | PLWH Longitudinal | Uninfected Longitudinal | PLWH vs. Uninfected Cross-Sectional | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V1 N = 38 1 | V2 N = 38 1 | V3 N = 38 1 | B1 N = 38 1 | V1 vs. V2 2 | V1 vs. V3 2 | V1 vs. B1 2 | V1 N = 24 1 | B1 N = 24 1 | V1 vs. B1 2 | V1 3 | B1 3 | |
| Memory T-Cell Subsets | ||||||||||||
| Naive (CD45RA+CD28+CCR7+) CD4 T-cells | 19.55 (14.62, 27.73) | 26.35 (19.67, 34.05) | 29.05 (18.27, 35.15) | 28.45 (21.33, 33.70) | <0.001 | <0.001 | <0.001 | 11.45 (6.65, 17.12) | 21.60 (8.47, 29.63) | 0.001 | 0.002 | 0.017 |
| Central Memory (CD45RA-CD28+CCR7+) CD4 T-cells | 42.35 (35.00, 48.13) | 42.60 (37.00, 49.88) | 42.90 (38.30, 50.08) | 44.10 (37.55, 48.95) | 0.14 | 0.3 | 0.5 | 48.05 (40.00, 51.85) | 48.20 (39.70, 55.08) | 0.2 | 0.048 | 0.043 |
| Transitional Memory (CD45RA-CD28+CCR7-) CD4 T-cells | 19.65 (15.50, 22.30) | 17.20 (14.83, 20.15) | 16.55 (14.33, 19.70) | 16.70 (13.55, 21.20) | 0.027 | 0.001 | <0.001 | 20.45 (16.25, 22.65) | 16.50 (13.78, 21.20) | 0.075 | 0.8 | 0.9 |
| Effector Memory (CD45RA-CD28-CCR7-) CD4 T-cells | 3.23 (1.62, 6.33) | 2.00 (1.27, 4.22) | 1.68 (1.01, 3.31) | 1.65 (0.98, 3.40) | <0.001 | <0.001 | <0.001 | 3.98 (2.61, 5.62) | 1.37 (0.76, 3.43) | 0.008 | 0.5 | 0.8 |
| Terminally Differentiated (CD45RA+CD28-CCR7-) CD4 T-cells | 0.35 (0.12, 0.70) | 0.23 (0.11, 0.45) | 0.23 (0.07, 0.49) | 0.16 (0.08, 0.43) | 0.004 | <0.001 | <0.001 | 0.28 (0.19, 0.47) | 0.12 (0.05, 0.32) | 0.025 | 0.7 | 0.4 |
| T-cell function | ||||||||||||
| % HLA-DR+CD38+ in CD4 T-cells | 2.32 (1.93, 3.67) | 2.18 (1.65, 2.86) | 2.04 (1.55, 2.69) | 2.23 (1.74, 2.96) | 0.010 | 0.016 | 0.013 | 2.57 (1.49, 4.92) | 2.36 (1.34, 3.83) | 0.2 | >0.9 | 0.6 |
| % Ki-67+ in CD4 T-cells | 2.70 (2.19, 3.98) | 2.30 (2.00, 3.11) | 2.32 (1.86, 2.72) | 2.39 (1.87, 2.85) | 0.074 | <0.001 | <0.001 | 2.36 (1.97, 3.03) | 2.12 (1.59, 2.80) | 0.3 | 0.070 | 0.4 |
| % PD-1+ in CD4 T-cells | 18.60 (14.67, 25.33) | 19.55 (15.12, 23.98) | 22.20 (16.35, 25.65) | 20.70 (16.12, 25.97) | 0.8 | 0.006 | 0.032 | 9.32 (6.68, 13.03) | 12.15 (11.38, 16.00) | 0.005 | <0.001 | <0.001 |
| % CTLA-4+ in CD4 T-cells | 2.96 (2.30, 3.94) | 3.16 (2.16, 4.29) | 2.73 (2.15, 3.79) | 2.70 (2.15, 3.56) | 0.2 | >0.9 | 0.7 | 2.44 (1.90, 3.15) | 2.84 (1.87, 3.89) | 0.023 | 0.056 | 0.8 |
| % Senescent (CD28-CD57+) in CD4 T-cells | 4.16 (0.98, 9.00) | 2.50 (0.70, 6.60) | 2.15 (0.71, 4.10) | 1.61 (0.85, 4.82) | <0.001 | <0.001 | <0.001 | 1.18 (0.37, 3.22) | 0.32 (0.13, 3.24) | 0.009 | 0.030 | 0.025 |
| % CD39+ in CD4 T-cells | 4.92 (3.66, 7.37) | 4.92 (3.73, 6.81) | 5.34 (4.00, 7.69) | 5.07 (3.65, 6.98) | 0.8 | 0.6 | 0.3 | 2.39 (1.90, 3.60) | 2.89 (2.29, 4.36) | 0.037 | <0.001 | 0.006 |
| % CD73+ in CD4 T-cells | 8.59 (5.22, 12.35) | 8.45 (5.77, 12.40) | 9.46 (6.10, 13.03) | 9.12 (5.98, 13.55) | 0.9 | 0.048 | 0.005 | 11.05 (9.23, 14.12) | 12.45 (8.56, 15.95) | 0.079 | 0.062 | 0.036 |
| Chemokine receptors | ||||||||||||
| % CCR4+ in CD4 T-cells | 17.05 (12.20, 22.48) | 16.95 (14.90, 23.58) | 19.15 (15.85, 22.48) | 18.75 (16.72, 22.87) | 0.5 | 0.006 | 0.073 | 16.35 (6.43, 23.18) | 16.25 (10.28, 24.33) | 0.2 | 0.4 | 0.5 |
| % CCR6+ in CD4 T-cells | 38.90 (36.12, 41.20) | 38.60 (35.75, 40.25) | 39.50 (36.97, 41.25) | 39.45 (37.45, 41.25) | 0.4 | 0.7 | 0.7 | 29.35 (19.60, 32.05) | 31.30 (22.10, 34.67) | 0.064 | <0.001 | <0.001 |
| % CXCR3+ in CD4 T-cells | 19.45 (13.15, 26.20) | 17.60 (13.48, 21.32) | 15.70 (13.62, 18.17) | 16.90 (12.60, 18.77) | 0.13 | 0.004 | 0.018 | 35.40 (25.35, 44.88) | 19.40 (12.20, 26.30) | <0.001 | <0.001 | 0.3 |
| Th subsets | ||||||||||||
| % Th17 in CD4 T-cells (CD45RA-CCR4+CCR6+ CXCR3-) | 4.46 (3.26, 6.66) | 4.99 (3.09, 7.38) | 6.10 (5.26, 8.09) | 6.11 (4.54, 7.16) | 0.3 | <0.001 | 0.010 | 1.45 (1.07, 2.12) | 3.19 (2.19, 4.48) | <0.001 | <0.001 | <0.001 |
| % Th1-Th17 in CD4 T-cells (CD45RA-CCR4-CCR6+CXCR3+) | 5.06 (2.71, 6.87) | 3.85 (2.49, 5.55) | 3.97 (2.91, 5.03) | 4.12 (2.78, 5.41) | 0.030 | 0.007 | 0.059 | 4.28 (2.42, 6.52) | 2.68 (2.04, 4.70) | 0.010 | 0.6 | 0.040 |
| % Th2 in CD4 T-cells (CD45RA-CCR4+CCR6-CXCR3-) | 6.36 (4.86, 8.41) | 7.88 (5.95, 10.06) | 8.66 (6.64, 10.31) | 9.11 (6.32, 10.40) | 0.053 | <0.001 | 0.014 | 4.97 (2.86, 9.53) | 8.57 (4.80, 10.48) | 0.004 | 0.2 | 0.8 |
| % Th1 in CD4 T-cells (CD45RA-CCR4-CCR6-CXCR3+) | 9.97 (5.84, 12.55) | 9.00 (5.82, 10.52) | 7.26 (5.65, 9.19) | 8.14 (5.84, 8.92) | 0.2 | 0.002 | 0.013 | 23.90 (15.40, 27.60) | 11.15 (6.63, 16.00) | <0.001 | <0.001 | 0.037 |
| Regulatory T-cells | ||||||||||||
| % Treg (CD25hi CD127lo FoxP3+) in CD4 T-cells | 0.82 (0.52, 1.23) | 1.03 (0.76, 1.95) | 1.44 (1.15, 2.17) | 1.56 (1.11, 1.97) | 0.003 | <0.001 | <0.001 | 0.64 (0.38, 1.05) | 1.21 (0.74, 2.24) | <0.001 | 0.14 | 0.5 |
| % CD39+ Treg in CD4 T-cells | 0.47 (0.28, 0.65) | 0.49 (0.30, 0.72) | 0.58 (0.43, 0.85) | 0.64 (0.34, 0.86) | 0.4 | 0.002 | 0.003 | 0.29 (0.13, 0.49) | 0.35 (0.22, 0.88) | 0.004 | 0.032 | 0.3 |
| % CD73+ Treg in CD4 T-cells | 0.04 (0.02, 0.07) | 0.05 (0.03, 0.11) | 0.09 (0.05, 0.15) | 0.08 (0.05, 0.14) | 0.022 | <0.001 | <0.001 | 0.05 (0.01, 0.10) | 0.13 (0.02, 0.18) | <0.001 | 0.8 | 0.7 |
| % CD39+CD73+ Treg in CD4 T-cells | 0.01 (0.01, 0.03) | 0.01 (0.01, 0.04) | 0.02 (0.02, 0.05) | 0.03 (0.01, 0.04) | 0.4 | <0.001 | 0.002 | 0.02 (0.00, 0.04) | 0.03 (0.01, 0.06) | 0.071 | 0.8 | 0.6 |
| Characteristics | PLWH Longitudinal | Uninfected Longitudinal | PLWH vs. Uninfected Cross-Sectional | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V1 N = 38 1 | V2 N = 38 1 | V3 N = 38 1 | B1 N = 38 1 | V1 vs. V2 2 | V1 vs. V3 2 | V1 vs. B1 2 | V1 N = 24 1 | B1 N = 24 1 | V1 vs. B1 2 | V1 3 | B1 3 | |
| Memory Subsets | ||||||||||||
| Naive (CD45RA+CD28+CCR7+) CD8 T-cells | 8.33 (5.07, 15.63) | 12.95 (8.91, 20.60) | 15.40 (8.79, 23.02) | 15.75 (7.96, 26.08) | <0.001 | <0.001 | <0.001 | 8.25 (4.69, 15.15) | 20.90 (8.97, 39.67) | <0.001 | 0.8 | 0.2 |
| Central Memory (CD45RA-CD28+CCR7+) CD8 T-cells | 7.70 (4.30, 13.47) | 8.49 (5.41, 16.05) | 10.90 (6.95, 16.50) | 10.90 (6.40, 16.65) | 0.027 | <0.001 | 0.003 | 8.01 (5.77, 10.50) | 12.45 (8.38, 14.82) | <0.001 | 0.9 | 0.6 |
| Transitional Memory (CD45RA-CD28+CCR7-) CD8 T-cells | 15.35 (10.60, 17.50) | 17.05 (11.28, 21.02) | 17.15 (13.12, 20.73) | 16.40 (12.22, 21.25) | 0.040 | 0.002 | 0.009 | 12.40 (9.21, 14.72) | 13.95 (11.88, 20.95) | 0.007 | 0.2 | 0.5 |
| Effector Memory (CD45RA-CD28-CCR7-) CD8 T-cells | 24.40 (17.35, 34.98) | 20.95 (14.72, 27.80) | 15.25 (11.77, 26.12) | 17.80 (12.00, 24.70) | 0.009 | <0.001 | <0.001 | 30.25 (21.68, 35.25) | 14.10 (9.51, 24.65) | <0.001 | 0.4 | 0.5 |
| Terminally Differentiated (CD45RA+CD28-CCR7-) CD8 T-cells | 13.90 (8.24, 19.17) | 11.90 (7.04, 15.90) | 9.93 (6.80, 16.45) | 9.82 (7.18, 12.60) | 0.001 | <0.001 | 0.021 | 8.25 (5.56, 11.25) | 5.74 (4.45, 8.03) | 0.010 | 0.009 | <0.001 |
| T-cell function | ||||||||||||
| % HLA-DR+CD38+ in CD8 T-cells | 3.55 (2.18, 7.49) | 3.91 (2.42, 6.18) | 3.72 (2.47, 5.25) | 3.97 (2.65, 7.11) | 0.5 | 0.7 | 0.4 | 3.02 (2.67, 4.22) | 3.15 (2.77, 3.88) | 0.9 | 0.4 | 0.12 |
| % Ki-67+ in CD8 T-cells | 1.90 (1.63, 2.55) | 2.13 (1.57, 2.66) | 1.83 (1.48, 2.24) | 1.84 (1.47, 2.46) | 0.3 | 0.045 | 0.023 | 2.20 (1.95, 2.51) | 1.95 (1.47, 2.23) | 0.042 | 0.2 | 0.9 |
| % PD-1+ in CD8 T-cells | 22.20 (17.20, 27.95) | 22.90 (15.57, 28.25) | 25.90 (18.68, 31.77) | 24.75 (17.45, 30.62) | 0.7 | 0.011 | 0.045 | 16.95 (11.62, 23.92) | 17.85 (15.23, 26.67) | 0.037 | 0.078 | 0.12 |
| % CTLA-4+ in CD8 T-cells | 1.16 (0.87, 1.52) | 1.13 (0.91, 1.41) | 1.15 (0.87, 1.40) | 1.11 (0.88, 1.57) | 0.9 | 0.4 | 0.6 | 1.23 (1.06, 1.59) | 1.11 (0.93, 1.39) | 0.4 | 0.4 | 0.7 |
| % Senescent (CD28-CD57+) in CD8 T-cells | 36.90 (25.90, 51.78) | 31.20 (23.35, 44.67) | 27.60 (17.48, 43.40) | 28.50 (18.40, 44.52) | <0.001 | <0.001 | <0.001 | 30.00 (15.23, 36.53) | 13.45 (9.50, 31.17) | 0.001 | 0.022 | 0.007 |
| % CD73+ in CD8 T-cells | 17.75 (11.53, 31.97) | 23.40 (14.95, 32.12) | 24.45 (18.42, 39.15) | 27.05 (19.02, 38.48) | 0.005 | <0.001 | <0.001 | 28.80 (22.80, 45.13) | 45.10 (35.62, 57.58) | <0.001 | 0.007 | <0.001 |
| % CD39+ in CD8 T-cells | 0.93 (0.70, 1.33) | 1.00 (0.66, 1.44) | 0.95 (0.61, 1.27) | 0.88 (0.47, 1.63) | 0.5 | 0.13 | 0.5 | 1.41 (0.96, 1.94) | 1.97 (1.21, 2.71) | 0.002 | 0.023 | <0.001 |
| Chemokine receptors | ||||||||||||
| % CCR4+ in CD8 T-cells | 35.45 (28.53, 46.92) | 33.00 (24.93, 43.13) | 22.55 (16.70, 28.25) | 23.40 (17.42, 35.88) | 0.4 | <0.001 | <0.001 | 39.65 (28.15, 52.00) | 35.25 (21.50, 46.45) | 0.005 | 0.4 | 0.059 |
| % CCR6+ in CD8 T-cells | 28.25 (26.80, 29.35) | 28.05 (25.83, 29.73) | 27.80 (25.15, 29.58) | 28.00 (25.85, 30.05) | >0.9 | 0.3 | >0.9 | 12.98 (5.14, 23.40) | 14.91 (4.63, 25.52) | 0.2 | <0.001 | <0.001 |
| % CXCR3+ in CD8 T-cells | 21.20 (12.85, 36.65) | 18.60 (12.85, 26.40) | 15.05 (12.05, 19.82) | 17.80 (11.47, 22.77) | 0.2 | <0.001 | 0.004 | 36.55 (30.50, 46.33) | 19.55 (13.40, 24.70) | <0.001 | <0.001 | 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandrova, Y.; Yero, A.; Mboumba Bouassa, R.-S.; Comeau, E.; Samarani, S.; Brumme, Z.L.; Hull, M.; Crawley, A.M.; Langlois, M.-A.; Angel, J.B.; et al. SARS-CoV-2 Vaccine-Induced T-Cell Response after Three Doses in People Living with HIV on Antiretroviral Therapy Compared to Seronegative Controls (CTN 328 COVAXHIV Study). Viruses 2023, 15, 575. https://0-doi-org.brum.beds.ac.uk/10.3390/v15020575
Alexandrova Y, Yero A, Mboumba Bouassa R-S, Comeau E, Samarani S, Brumme ZL, Hull M, Crawley AM, Langlois M-A, Angel JB, et al. SARS-CoV-2 Vaccine-Induced T-Cell Response after Three Doses in People Living with HIV on Antiretroviral Therapy Compared to Seronegative Controls (CTN 328 COVAXHIV Study). Viruses. 2023; 15(2):575. https://0-doi-org.brum.beds.ac.uk/10.3390/v15020575
Chicago/Turabian StyleAlexandrova, Yulia, Alexis Yero, Ralph-Sydney Mboumba Bouassa, Eve Comeau, Suzanne Samarani, Zabrina L. Brumme, Mark Hull, Angela M. Crawley, Marc-André Langlois, Jonathan B. Angel, and et al. 2023. "SARS-CoV-2 Vaccine-Induced T-Cell Response after Three Doses in People Living with HIV on Antiretroviral Therapy Compared to Seronegative Controls (CTN 328 COVAXHIV Study)" Viruses 15, no. 2: 575. https://0-doi-org.brum.beds.ac.uk/10.3390/v15020575