
Evaluation of NS4A, NS4B, NS5 and 3′UTR Genetic Determinants of WNV Lineage 1 Virulence in Birds and Mammals
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Viruses
2.2. Generation of Chimeric Viruses
2.3. WNV Plaque Phenotype
2.4. Viral Growth Kinetics
2.5. Quantitative RT-PCR
2.6. Virus Titration by TCID50
2.7. Virulence in Mice
2.8. Virulence in Specific-Pathogen-Free (SPF) Chickens
2.9. Statistical Analysis
3. Results
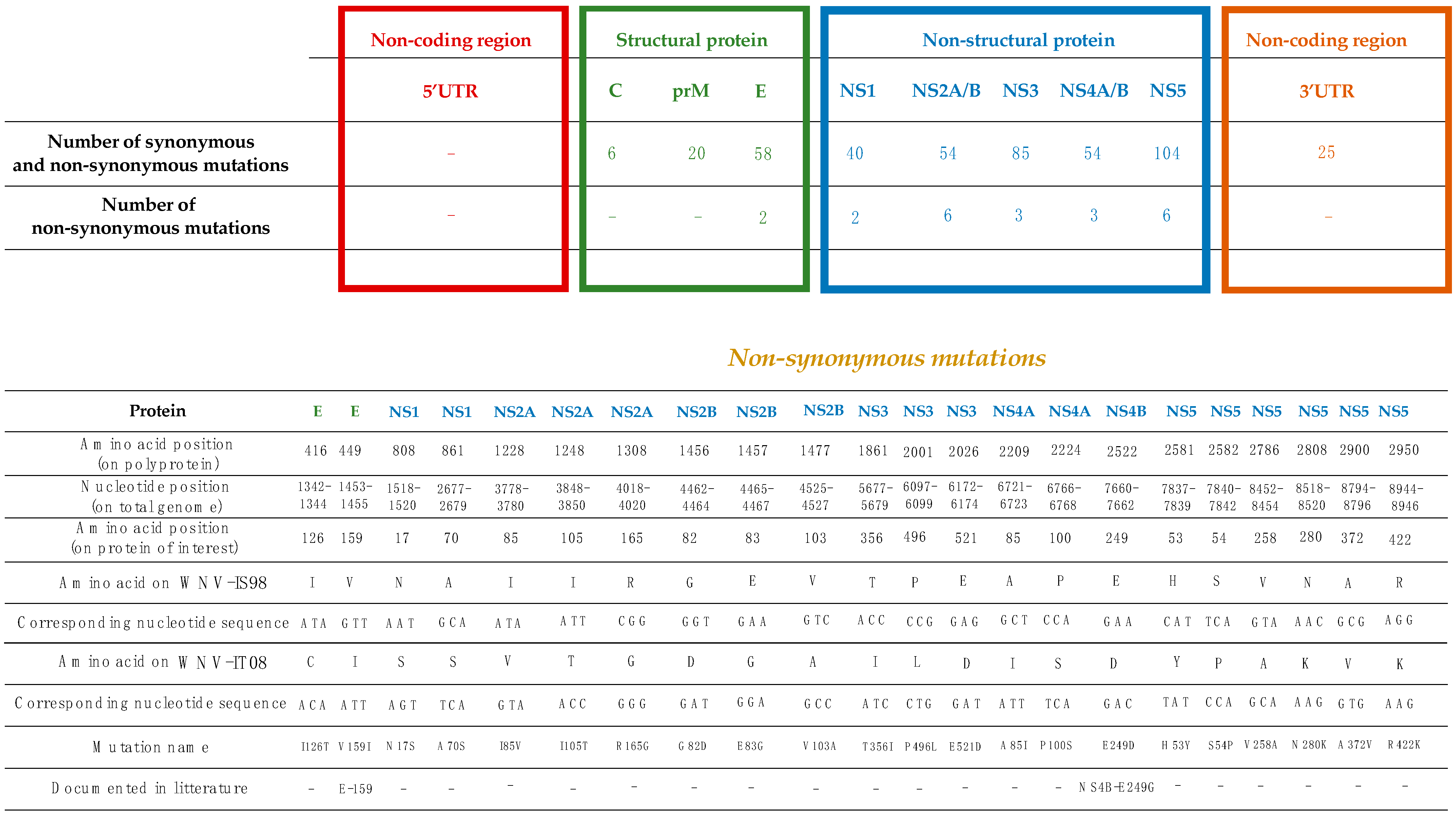
3.1. Comparison of Genome Sequences of Parental WNV-IS98 and IT08 Strains
3.2. Viral Replication in Vero Cells
3.3. Replication Kinetics of the Parental WNV-IS98 and IT08 and Chimeras thereof in Mammalian and Avian Cell Culture
3.4. Replication of Parental and Chimeric Viruses in BALB/cByJ Mice
3.5. Neuroinvasion and Neurovirulence of Parental Strains and Chimeras in BALB/cByJ Mice
3.6. Replication of Parental and Chimeric Viruses in SPF Chickens
3.7. Neurovirulence of Parental Strains and Chimeras in SPF Chickens
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Sequence | Literature Known Molecular Markers | Role | References | Comparison to Our Study | Conclusion for Our Study |
|---|---|---|---|---|---|
| NS4 | NS4A-F92L | Increase virulence in birds | [58] | NS4A-92 amino acid is the same between IS98 and IT08 so could not explain any difference in chicks | No role of NS4A-92 amino acid in modulating virulence in chicks |
| NS4 | NS4B-E249G | The substitution of the glutamic acid at position 249 (E249) was shown to be crucial for conferring an attenuated phenotype in mice and it was identified in an attenuated strain isolated from birds in 2003 in Texas | [60,61] | RNA quantity in oral swab if IT08 and IS98-NS4A/NS4B/5′NS5 are significantly lower than IS98 and IS98-NS5/3′UTR at 7 days p.i. → suggesting a role of NS4A/NS4B/5′NS5 in reduced virulence in birds IT08 possesses NS4B-D249 and IS98 possesses E249 | Possible role of NS4B-E249D mutation in IT08 or chimera IS98-NS4A/NS4B/5′NS5 in attenuated virulence in chicks |
| NS5 | K-D-K-E motif (NS5-K61-D146-K182-E218) | Candidate for DENV vaccine | [57,63] | No difference in K-D-K-E motif between IS98 and IT08 (Appendix A Figure A1) | Known molecular markers of virulence of DENV-2, a flavivirus closely related to WNV, cannot, by extension, explain differences observed between WNV IS98 and IT08 and corresponding chimeras |
| NS5 | NS5-R325A, NS5-R519A, NS5-R769A, NS5-K840A, NS5-R841A | High RNA synthesis activity in vitro but delayed or impaired replication in vivo | [64] | Sequence alignment between DENV-2 and our parental strain does not show any difference between IS98 and IT08 (Appendix A Figure A1) 100% identity between IS98/IT08 at these sites and DENV-2 virus, confirming their potential role in flavivirus replication | Known molecular markers of virulence of DENV-2, a flavivirus closely related to WNV, cannot, by extension, explain differences observed between WNV IS98 and IT08 and corresponding chimeras |
| NS5 | PBM motif (three amino acids, T-V-L), just upstream the codon stops | C-term PBM of WNV NS5 recognizes several human PDZ PBM is known to be implicated in virulence | [65] | Sequence alignment of IS98 and IT08 does not show any difference in PBM motif (Appendix A Figure A1) | Known molecular markers of virulence of DENV-2, a flavivirus closely related to WNV, cannot, by extension, explain differences observed between WNV IS98 and IT08 and corresponding chimeras |
| NS5 | Threonine phosphorylation (Thr449) | Regulatory roles | [66,67] | Sequence alignment of IS98 and IT08 does not show any differences in PBM motif (Appendix A Figure A2) | Known molecular markers of virulence of DENV-2, a flavivirus closely related to WNV, cannot, by extension, explain differences observed between WNV IS98 and IT08 and corresponding chimeras |
| NS5 | General modifications in NS5 | NS5 protein is known to frequently use multiple strategies to suppress the JAK-STAT signaling pathway and consequently type I IFN response to mammalian NS5. ZIKV inhibits the phosphorylation of STAT1 and leads to degradation of STAT2; JEV, DENV, and WNV antagonize this pathway too STAT2 is frequently targeted for NS5-mediated IFN suppression in flaviviruses, so modifications in NS5 could impact host immune response to viral infection | [69] | IS98-NS5/3′UTR and IT08 engender reduced mortality in mice at 12 days p.i. compared to IS98 It is possible that IT08 and IS98-NS5/3′UTR present molecular features that could not inhibit host innate response | Possible role of NS5 in decreased virulence of IT08 in a mammalian model, maybe by influencing host IFN response |
| 3′UTR | 3′-terminal stem-loop (3′SL) | Interaction between 5′UTR and 3′UTR is implied in viral RNA replication. This interaction is important for NS5 polymerase recruitment. Role of 3′SL in viral replication of DENV | [71] | None of the 25 3′UTR mutations found between IS98 and IT08 are in 3′SL, suggesting that the difference of viral replication between IS98 and IT08 could not be explained by a difference in cyclization depending on 3′SL sequence | No role of 3′SL sequence in attenuation of virulence of IT08 or IS98-NS5/3′UTR in a mammalian model |
| 3′UTR | Double deletion mutant nt 10,474–10,478/nt 10562–10566 in DENV | Viral translation rate was reduced by 60% in mammalian cells | [72] | Sequence alignment of 3′UTR region of DENV-2 virus and parental strain, IS98 and IT08 show 100% identity in the 10562–10566 sequence between the four viruses (Appendix A Figure A3) However, 10,474–10,478 differs in one nucleotide between IS98 and IT08. Nucleotide 10,478 of IT08 is the same as DENV-2 (nt T), whereas it is different in IS98 (nt C). That could impact viral translation | Possible role of nucleotide 10,478 in reduced virulence of IT08 and IS98-NS5/3′UTR in a mammalian model |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A



References
- Smithburn, K.; Hugues, T.; Burke, A.; Paul, J. A neurotropic virus isolated from the blood of a native of uganda. Am. J. Trop. Med. Hyg. 1940, s1-20, 471. [Google Scholar] [CrossRef]
- Suthar, M.S.; Diamond, M.S.; Gale, M. West Nile virus infection and immunity. Nat. Rev. Microbiol. 2013, 2, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Hubálek, Z.; Halouzka, J. West Nile Fever—A Reemerging Mosquito-Borne Viral Disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.J.; Haddad, M.B.; Tierney, B.C.; Campbell, G.L.; Marfin, A.A.; Van Gerpen, J.A.; Fleischauer, A.; Leis, A.A.; Stokic, D.S.; Petersen, L.R. Neurologic Manifestations and Outcome of West Nile Virus Infection. JAMA 2003, 290, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Li, J.; Shi, P.Y. West Nile virus. Lancet Neurol. 2007, 6, 171–181. [Google Scholar] [CrossRef]
- Pealer, L.N.; Marfin, A.A.; Petersen, L.R.; Lanciotti, R.S.; Page, P.L.; Stramer, S.L.; Chamberland, M.E. Transmission of West Nile virus through blood transfusion in the United States in 2002. N. Engl. J. Med. 2003, 349, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.R. Epidemiology of West Nile Virus in the United States: Implications for Arbovirology and Public Health. J. Med. Entomol. 2019, 56, 1456–1462. [Google Scholar] [CrossRef]
- Iwamoto, M.; Jernigan, D.B.; Guasch, A.; Trepka, M.J.; Blackmore, C.G.; Hellinger, W.C.; Petersen, L.R. Transmission of West Nile virus from an organ donor to four transplant recipients. N. Engl. J. Med. 2003, 22, 2196–2203. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Savic, V.; Petrovic, T.; Toplak, I.; Barbic, L.; Petric, D.; Savini, G. Emerging Trends in the Epidemiology of West Nile and Usutu Virus Infections in Southern Europe. Front. Vet. Sci. 2019, 6, 437. [Google Scholar] [CrossRef]
- Brustolin, M.; Talavera, S.; Santamaría, C.; Rivas, R.; Pujol, N.; Aranda, C.; Busquets, N. Culex pipiens and Stegomyia albopicta (=Aedes albopictus) populations as vectors for lineage 1 and 2 West Nile virus in Europe. Med. Vet. Entomol. 2016, 30, 166–173. [Google Scholar] [CrossRef]
- Komar, N.; Langevin, S.; Hinten, S. Experimental Infection of North American Birds with the New York 1999 Strain of West Nile Virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E.B.; Komar, N.; Nasci, R.S.; Montgomery, S.P.; O’Leary, D.R.; Campbell, G.L. Epidemiology and transmission dynamics of West Nile virus disease. Emerg. Infect. Dis. 2005, 11, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.J.; Leis, A.A.; Stokic, D.S.; Van Gerpen, J.A.; Marfin, A.A.; Webb, R.; Petersen, L.R. Acute Flaccid Paralysis and West Nile Virus Infection. Emerg. Infect. Dis. 2003, 9, 788–793. [Google Scholar] [CrossRef]
- Briese, T.; Jia, X.Y.; Huang, C.; Grady, L.J.; Lipkin, W.I. Identification of a Kunjin/West Nile-like flavivirus in brains of patients with New York encephalitis. Lancet 1999, 354, 1261–1262. [Google Scholar] [CrossRef]
- Lanciotti, R.; Roehrig, J.; Deubel, V.; Smith, J. Origin of the West Nile Virus Responsible for an Outbreak of Encephalitis in the Northeastern United States. Science 1999, 286, 2333–23337. [Google Scholar] [CrossRef]
- Jia, X.Y.; Briese, T.; Jordan, I.; Rambaut, A.; Chi, H.C.; Mackenzie, J.S.; Lipkin, W.I. Genetic analysis of West Nile New York 1999 encephalitis virus. Lancet 1999, 354, 1971–1972. [Google Scholar] [CrossRef]
- Ronca, S.E.; Murray, K.O.; Nolan, M.S. Cumulative Incidence of West Nile Virus Infection, Continental United States, 1999–2016. Emerg Infect Dis. 2019, 25, 325–327. [Google Scholar] [CrossRef]
- Barrett, A.D.T. West Nile in Europe: An increasing public health problem. J. Travel. Med. 2018, 25, tay096. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Montarsi, F.; Quaranta, E.; Monne, I.; Pacenti, M.; Michelutti, A.; Capelli, G. Early start of seasonal transmission and co-circulation of West Nile virus lineage 2 and a newly introduced lineage 1 strain, northern Italy, June 2022. Euro Surveill. 2022, 27, 2200548. [Google Scholar] [CrossRef] [PubMed]
- Shlomit, P.; Jan, C.S. Environmental drivers of West Nile fever epidemiology in Europe and Western Asia—A review. Int. J. Environ. Res. Public. Health 2013, 10, 3534–3562. [Google Scholar]
- Brown, H.E.; Childs, J.E.; Diuk-Wasser, M.A.; Fish, D. Ecologic Factors Associated with West Nile Virus Transmission, Northeastern United States. Emerg. Infect. Dis. 2008, 14, 1539–1545. [Google Scholar] [CrossRef] [PubMed]
- Marcantonio, M.; Rizzoli, A.; Metz, M.; Rosà, R.; Marini, G.; Chadwick, E.; Neteler, M. Identifying the environmental conditions favouring West Nile Virus outbreaks in Europe. PLoS ONE 2015, 10, e0121158. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, J.A.; Wang, T.; Barrett, A.D. Virulence determinants of West. Nile virus: How can these be used for vaccine design? Future Virol. 2017, 12, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.E.; Pierson, T.C.; Hubka, S.; Rucker, S.; Gordon, I.J.; Enama, M.E.; Vaccine Research Center 302 Study Team. A West Nile virus DNA vaccine induces neutralizing antibody in healthy adults during a phase 1 clinical trial. J. Infect. Dis. 2007, 196, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Durbin, A.P.; Wright, P.F.; Cox, A.; Kagucia, W.; Elwood, D.; Henderson, S.; Pletnev, A.G. The live attenuated chimeric vaccine rWN/DEN4Δ30 is well-tolerated and immunogenic in healthy flavivirus-naïve adult volunteers. Vaccine 2013, 31, 5772–5777. [Google Scholar] [CrossRef] [PubMed]
- Woods, C.W.; Sanchez, A.M.; Swamy, G.K.; McClain, M.T.; Harrington, L.; Freeman, D.; Walter, E.B. An observer blinded, randomized, placebo-controlled, phase I dose escalation trial to evaluate the safety and immunogenicity of an inactivated West Nile virus Vaccine, HydroVax-001, in healthy adults. Vaccine 2019, 37, 4222–4230. [Google Scholar] [CrossRef]
- Orlinger, K.K.; Holzer, G.W.; Schwaiger, J.; Mayrhofer, J.; Schmid, K.; Kistner, O.; Falkner, F.G. An inactivated West Nile Virus vaccine derived from a chemically synthesized cDNA system. Vaccine 2010, 28, 3318–3324. [Google Scholar] [CrossRef]
- Monath, T.P.; Liu, J.; Kanesa-Thasan, N.; Myers, G.A.; Nichols, R.; Deary, A.; Bedford, P. A live, attenuated recombinant West Nile virus vaccine. Proc. Natl. Acad. Sci. USA 2006, 103, 6694–6699. [Google Scholar] [CrossRef]
- Biedenbender, R.; Bevilacqua, J.; Gregg, A.M.; Watson, M.; Dayan, G. Phase II, Randomized, Double-Blind, Placebo-Controlled, Multicenter Study to Investigate the Immunogenicity and Safety of a West Nile Virus Vaccine in Healthy Adults. J. Infect. Dis. 2011, 203, 75–84. [Google Scholar] [CrossRef]
- Dayan, G.H.; Bevilacqua, J.; Coleman, D.; Buldo, A.; Risi, G. Phase II, dose ranging study of the safety and immunogenicity of single dose West Nile vaccine in healthy adults ≥ 50 years of age. Vaccine 2012, 30, 6656–6664. [Google Scholar] [CrossRef]
- Fiacre, L.; Pagès, N.; Albina, E.; Richardson, J.; Lecollinet, S.; Gonzalez, G. Molecular Determinants of West Nile Virus Virulence and Pathogenesis in Vertebrate and Invertebrate Hosts. Int. J. Mol. Sci. 2020, 21, 9117. [Google Scholar] [CrossRef]
- Audsley, M.; Edmonds, J.; Liu, W.; Mokhonov, V.; Mokhonova, E.; Melian, E.B.; Khromykh, A.A. Virulence determinants between New York 99 and Kunjin strains of West Nile virus. Virology 2011, 414, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Shirato, K.; Miyoshi, H.; Goto, A.; Ako, Y.; Ueki, T.; Kariwa, H.; Takashima, I. Viral envelope protein glycosylation is a molecular determinant of the neuroinvasiveness of the New York strain of West Nile virus. J. Gen. Virol. 2004, 85, 3637–3645. [Google Scholar] [CrossRef]
- Malkinson, M.; Banet, C.; Weisman, Y.; Pokamunski, S.; King, R.; Deubel, V. Introduction of West Nile virus in the Middle East by migrating white storks. Emerg. Infect. Dis. 2002, 8, 392–397. [Google Scholar] [CrossRef]
- Bin, H.; Grossman, Z.; Pokamunski, S.; Malkinson, M.; Weiss, L.; Duvdevani, P.; Mendelson, E. West Nile fever in Israel 1999–2000: From geese to humans. Ann. N. Y. Acad. Sci. 2001, 951, 127–142. [Google Scholar] [CrossRef]
- Dridi, M.; Rauw, F.; Muylkens, B.; Lecollinet, S.; van den Berg, T.; Lambrecht, B. Setting up a SPF chicken model for the pathotyping of West Nile virus (WNV) strains. Transbound. Emerg. Dis. 2013, 60, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Shiryaev, S.A.; Chernov, A.V.; Aleshin, A.E.; Shiryaeva, T.N.; Strongin, A.Y. NS4A regulates the ATPase activity of the NS3 helicase: A novel cofactor role of the non-structural protein NS4A from West Nile virus. J. Gen. Virol. 2009, 90, 2081–2085. [Google Scholar] [CrossRef]
- Ambrose, R.L.; Mackenzie, J.M. A Conserved Peptide in West Nile Virus NS4A Protein Contributes to Proteolytic Processing and Is Essential for Replication. J. Virol. 2011, 85, 11274–11282. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Seeger, C. Differential Effects of Mutations in NS4B on West Nile Virus Replication and Inhibition of Interferon Signaling. J. Virol. 2007, 81, 11809–11816. [Google Scholar] [CrossRef]
- Wicker, J.A.; Whiteman, M.C.; Beasley, D.W.; Davis, C.T.; McGee, C.E.; Lee, J.C.; Barrett, A.D. Mutational analysis of the West Nile virus NS4B protein. Virology 2012, 426, 22–33. [Google Scholar] [CrossRef]
- Wicker, J.A.; Whiteman, M.C.; Beasley, D.W.; Davis, C.T.; Zhang, S.; Schneider, B.S.; Barrett, A.D. A single amino acid substitution in the central portion of the West Nile virus NS4B protein confers a highly attenuated phenotype in mice. Virology 2006, 349, 245–253. [Google Scholar] [CrossRef]
- Andrade, C.C.; Maharaj, P.D.; Reisen, W.K.; Brault, A.C. North American West Nile virus genotype isolates demonstrate differential replicative capacities in response to temperature. J. Gen. Virol. 2011, 92, 2523–2533. [Google Scholar] [CrossRef] [PubMed]
- Van Slyke, G.A.; Jia, Y.; Whiteman, M.C.; Wicker, J.A.; Barrett, A.D.T.; Kramer, L.D. Vertebrate attenuated West Nile virus mutants have differing effects on vector competence in Culex tarsalis mosquitoes. J. Gen. Virol. 2013, 94, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.N.; Wu, R.H.; Chen, S.L.; Chen, C.T.; Yueh, A. Mutagenesis of the dengue virus NS4A protein reveals a novel cytosolic N-terminal domain responsible for virus-induced cytopathic effects and intramolecular interactions within the N-terminus of NS4A. J. Gen. Virol. 2019, 100, 457–470. [Google Scholar] [CrossRef]
- Li, X.D.; Li, X.F.; Ye, H.Q.; Deng, C.L.; Ye, Q.; Shan, C.; Zhang, B. Recovery of a chemically synthesized Japanese encephalitis virus reveals two critical adaptive mutations in NS2B and NS4A. J. Gen. Virol. 2014, 95, 806–815. [Google Scholar] [CrossRef]
- Collins, N.D.; Beck, A.S.; Widen, S.G.; Wood, T.G.; Higgs, S.; Barrett, A.D.T. Structural and Nonstructural Genes Contribute to the Genetic Diversity of RNA Viruses. MBio 2018, 9, e01871-18. [Google Scholar] [CrossRef]
- Kaiser, J.A.; Luo, H.; Widen, S.G.; Wood, T.G.; Huang CY, H.; Wang, T.; Barrett, A.D. Genotypic and phenotypic characterization of West Nile virus NS5 methyltransferase mutants. Vaccine 2019, 37, 7155–7164. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.T.; Galbraith, S.E.; Zhang, S.; Whiteman, M.C.; Li, L.; Kinney, R.M.; Barrett, A.D. A Combination of Naturally Occurring Mutations in North American West Nile Virus Nonstructural Protein Genes and in the 3′ Untranslated Region Alters Virus Phenotype. J. Virol. 2007, 81, 6111–6116. [Google Scholar] [CrossRef]
- Bahuon, C.; Despres, P.; Pardigon, N.; Panthier, J.J.; Cordonnier, N.; Lowenski, S.; Lecollinet, S. IS-98-ST1 West Nile virus derived from an infectious cDNA clone retains neuroinvasiveness and neurovirulence properties of the original virus. PLoS ONE 2012, 7, e47666. [Google Scholar] [CrossRef]
- Linke, S.; Ellerbrok, H.; Niedrig, M.; Nitsche, A.; Pauli, G. Detection of West Nile virus lineages 1 and 2 by real-time PCR. J. Virol. Methods 2007, 146, 355–358. [Google Scholar] [CrossRef]
- Toussaint, J.F.; Sailleau, C.; Breard, E.; Zientara, S.; De Clercq, K. Bluetongue virus detection by two real-time RT-qPCRs targeting two different genomic segments. J. Virol. Methods 2007, 140, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Jaimipak, T.; Yoksan, S.; Ubol, S.; Pulmanausahakul, R. Small plaque size variant of chikungunya primary isolate showed reduced virulence in mice. Asian Pac. J. Allergy Immunol. 2018, 36, 201–205. [Google Scholar] [PubMed]
- Dalrymple, N.A.; Cimica, V.; Mackow, E.R. Dengue Virus NS Proteins Inhibit RIG-I/MAVS Signaling by Blocking TBK1/IRF3 Phosphorylation: Dengue Virus Serotype 1 NS4A Is a Unique Interferon-Regulating Virulence Determinant. MBio 2015, 6, e00553-15. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Jordán, J.L.; Laurent-Rolle, M.; Ashour, J.; Martínez-Sobrido, L.; Ashok, M.; Lipkin, W.I.; García-Sastre, A. Inhibition of Alpha/Beta Interferon Signaling by the NS4B Protein of Flaviviruses. J. Virol. 2005, 79, 8004–8013. [Google Scholar] [CrossRef] [PubMed]
- Knyazhanskaya, E.; Morais, M.C.; Choi, K.H. Flavivirus enzymes and their inhibitors. Enzymes 2021, 49, 265–303. [Google Scholar] [PubMed]
- Züst, R.; Li, S.H.; Xie, X.; Velumani, S.; Chng, M.; Toh, Y.X.; Fink, K. Characterization of a candidate tetravalent vaccine based on 2’-O-methyltransferase mutants. PLoS ONE 2018, 13, e0189262. [Google Scholar] [CrossRef]
- Kinney, R.M.; Huang, C.Y.H.; Whiteman, M.C. Avian virulence and thermostable replication of the North American strain of West Nile virus. J. Gen. Virol. 2006, 87, 3611–3622. [Google Scholar] [CrossRef]
- Lim, S.P.; Noble, C.G.; Shi, P.Y. The dengue virus NS5 protein as a target for drug discovery. Antivir. Res. 2015, 119, 57–67. [Google Scholar] [CrossRef]
- Davis, C.T.; Beasley, D.W.; Guzman, H.; Siirin, M.; Parsons, R.E.; Tesh, R.B.; Barrett, A.D. Emergence of attenuated West Nile virus variants in Texas, 2003. Virology 2004, 330, 342–350. [Google Scholar] [CrossRef]
- Puig-Basagoiti, F.; Tilgner, M.; Bennett, C.J.; Zhou, Y.; Muñoz-Jordán, J.L.; García-Sastre, A.; Shi, P.Y. A mouse cell-adapted NS4B mutation attenuates West Nile virus RNA synthesis. Virology 2007, 361, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Klema, V.J.; Padmanabhan, R.; Choi, K.H. Flaviviral Replication Complex: Coordination between RNA Synthesis and 5’-RNA Capping. Viruses 2015, 7, 4640–4656. [Google Scholar] [CrossRef]
- Zhou, Y.; Ray, D.; Zhao, Y.; Dong, H.; Ren, S.; Li, Z.; Li, H. Structure and function of flavivirus NS5 methyltransferase. J. Virol. 2007, 81, 3891–3903. [Google Scholar] [CrossRef]
- Iglesias, N.G.; Filomatori, C.V.; Gamarnik, A.V. The F1 Motif of Dengue Virus Polymerase NS5 Is Involved in Promoter-Dependent RNA Synthesis. J. Virol. 2011, 85, 5745–5756. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Del Val, C.O.; Caillet-Saguy, C.; Zehrouni, N.; Khou, C.; Caillet, J.; Wolff, N. Role of PDZ-binding motif from West Nile virus NS5 protein on viral replication. Sci. Rep. 2021, 11, 3266. [Google Scholar] [CrossRef]
- Morozova, O.V.; Tsekhanovskaya, N.A.; Maksimova, T.G.; Bachvalova, V.N.; Matveeva, V.A.; Kit, Y.; Ya, N. Phosphorylation of tick-borne encephalitis virus NS5 protein. Virus Res. 1997, 49, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.E.; Gorbalenya, A.E.; Rice, C.M. The NS5A/NS5 proteins of viruses from three genera of the family flaviviridae are phosphorylated by associated serine/threonine kinases. J. Virol. 1998, 72, 6199–6206. [Google Scholar] [CrossRef]
- Best, S.M. The Many Faces of the Flavivirus NS5 Protein in Antagonism of Type I Interferon Signaling. J. Virol. 2017, 91, e01970-16. [Google Scholar] [CrossRef]
- Thurmond, S.; Wang, B.; Song, J.; Hai, R. Suppression of Type I Interferon Signaling by Flavivirus NS5. Viruses 2018, 10, E712. [Google Scholar] [CrossRef]
- Yong, X.E.; Raghuvamsi, P.V.; Anand, G.S.; Wohland, T.; Sharma, K.K. Dengue virus strain 2 capsid protein switches the annealing pathway and reduces intrinsic dynamics of the conserved 5’ untranslated region. RNA Biol. 2021, 18, 718–731. [Google Scholar] [CrossRef]
- Meyer, A.; Freier, M.; Schmidt, T.; Rostowski, K.; Zwoch, J.; Lilie, H.; Friedrich, S. An RNA Thermometer Activity of the West Nile Virus Genomic 3’-Terminal Stem-Loop Element Modulates Viral Replication Efficiency during Host Switching. Viruses 2020, 12, E104. [Google Scholar] [CrossRef] [PubMed]
- Manzano, M.; Reichert, E.D.; Polo, S.; Falgout, B.; Kasprzak, W.; Shapiro, B.A.; Padmanabhan, R. Identification of cis-acting elements in the 3’-untranslated region of the dengue virus type 2 RNA that modulate translation and replication. J. Biol. Chem. 2011, 286, 22521–22534. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S. Virus and host determinants of West Nile virus pathogenesis. PLoS Pathog. 2009, 5, e1000452. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Takeda, K.; Markoff, L. Protein-protein interactions among West Nile non-structural proteins and transmembrane complex formation in mammalian cells. Virology 2013, 446, 365–377. [Google Scholar] [CrossRef]
- Tay, M.Y.F.; Vasudevan, S.G. The Transactions of NS3 and NS5 in Flaviviral RNA Replication. Adv. Exp. Med. Biol. 2018, 1062, 147–163. [Google Scholar]
- Phong, W.Y.; Moreland, N.J.; Lim, S.P.; Wen, D.; Paradkar, P.N.; Vasudevan, S.G. Dengue protease activity: The structural integrity and interaction of NS2B with NS3 protease and its potential as a drug target. Biosci. Rep. 2011, 31, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Xie, X.; Wang, Q.Y.; Dong, H.; Lee, M.Y.; Kang, C.; Shi, P.Y. Characterization of dengue virus NS4A and NS4B protein interaction. J. Virol. 2015, 89, 3455–3470. [Google Scholar] [CrossRef]
- Zou, J.; Xie, X.; Lee, L.T.; Chandrasekaran, R.; Reynaud, A.; Yap, L.; Shi, P.Y. Dimerization of flavivirus NS4B protein. J. Virol. 2014, 88, 3379–3391. [Google Scholar] [CrossRef]
- Umareddy, I.; Chao, A.; Sampath, A.; Gu, F.; Vasudevan, S.G. Dengue virus NS4B interacts with NS3 and dissociates it from single-stranded RNA. J. Gen. Virol. 2006, 87, 2605–2614. [Google Scholar] [CrossRef]
- Kapoor, M.; Zhang, L.; Ramachandra, M.; Kusukawa, J.; Ebner, K.E.; Padmanabhan, R. Association between NS3 and NS5 proteins of dengue virus type 2 in the putative RNA replicase is linked to differential phosphorylation of NS5. J. Biol. Chem. 1995, 270, 19100–19106. [Google Scholar] [CrossRef]
- Alsaleh, K.; Khou, C.; Frenkiel, M.P.; Lecollinet, S.; Vazquez, A.; de Arellano, E.R.; Pardigon, N. The E glycoprotein plays an essential role in the high pathogenicity of European-Mediterranean IS98 strain of West Nile virus. Virology 2016, 492, 53–65. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiacre, L.; Lowenski, S.; Bahuon, C.; Dumarest, M.; Lambrecht, B.; Dridi, M.; Albina, E.; Richardson, J.; Zientara, S.; Jiménez-Clavero, M.-Á.; et al. Evaluation of NS4A, NS4B, NS5 and 3′UTR Genetic Determinants of WNV Lineage 1 Virulence in Birds and Mammals. Viruses 2023, 15, 1094. https://0-doi-org.brum.beds.ac.uk/10.3390/v15051094
Fiacre L, Lowenski S, Bahuon C, Dumarest M, Lambrecht B, Dridi M, Albina E, Richardson J, Zientara S, Jiménez-Clavero M-Á, et al. Evaluation of NS4A, NS4B, NS5 and 3′UTR Genetic Determinants of WNV Lineage 1 Virulence in Birds and Mammals. Viruses. 2023; 15(5):1094. https://0-doi-org.brum.beds.ac.uk/10.3390/v15051094
Chicago/Turabian StyleFiacre, Lise, Steeve Lowenski, Céline Bahuon, Marine Dumarest, Bénédicte Lambrecht, Maha Dridi, Emmanuel Albina, Jennifer Richardson, Stéphan Zientara, Miguel-Ángel Jiménez-Clavero, and et al. 2023. "Evaluation of NS4A, NS4B, NS5 and 3′UTR Genetic Determinants of WNV Lineage 1 Virulence in Birds and Mammals" Viruses 15, no. 5: 1094. https://0-doi-org.brum.beds.ac.uk/10.3390/v15051094