

Querying Recombination Junctions of Replication-Competent Adeno-Associated Viruses in Gene Therapy Vector Preparations with Single Molecule, Real-Time Sequencing

Abstract
:1. Introduction
2. Materials and Methods
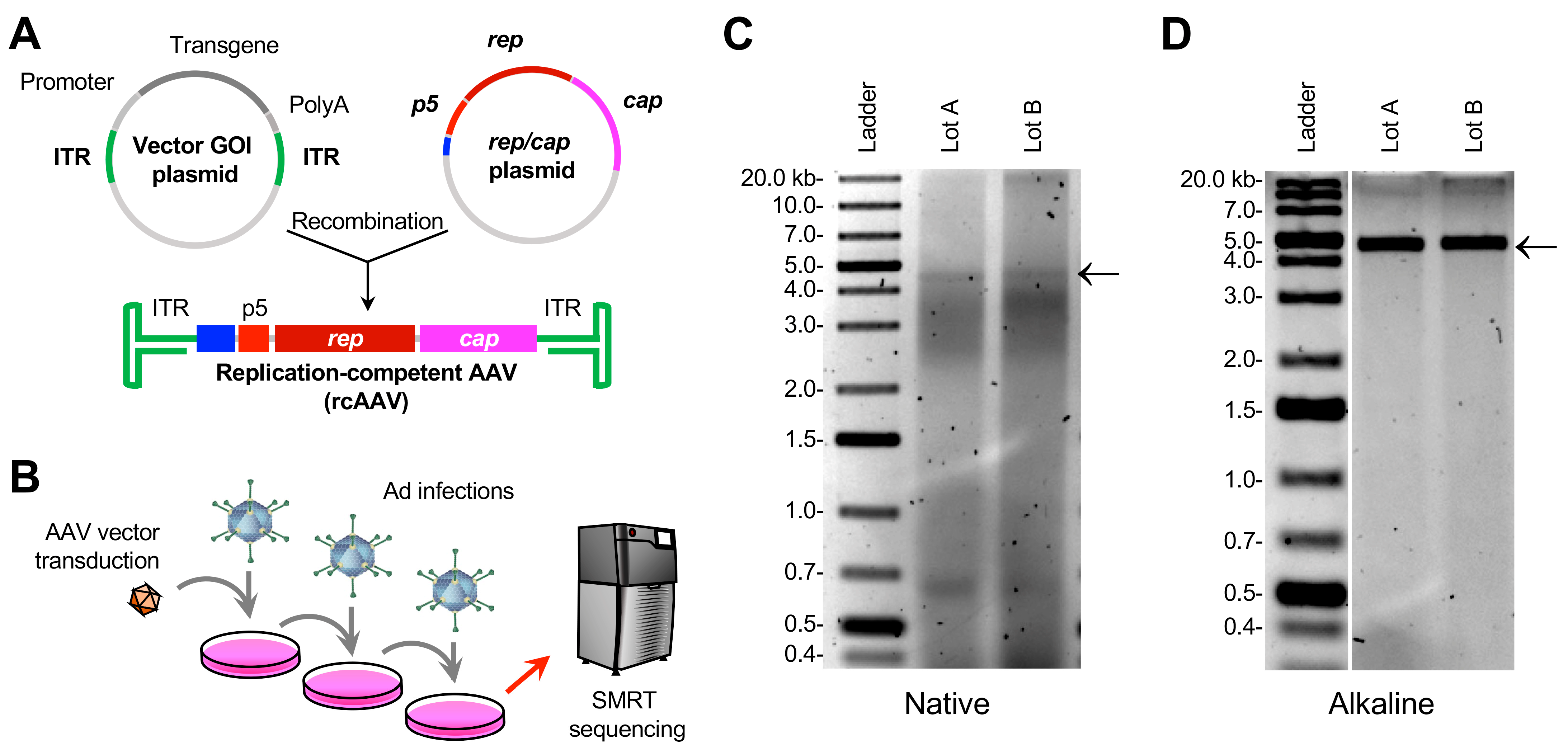
2.1. Plasmid and Vector Production
2.2. Propagation of rcAAV
2.3. Vector DNA Extraction and Agarose Gel Electrophoresis
2.4. SMRT Sequencing and AAV-GPseq
2.5. Bioinformatics
3. Results
3.1. Amplification of rcAAVs
3.2. Isolation of DNA from Crude Lysates of Cells Transduced with Expanded rcAAVs Yields Heterogeneous ~5-kb Genomes
3.3. Transgene-Related Sequences Were Rare in rcAAV-Enriched Lysates
3.4. Mapping Reads to a Reconstructed rcAAV Reference Revealed ITRs That Were Contiguous with rep and cap Sequences
3.5. Formation of rcAAVs Occur via Non-Specific Recombination Events at the 5′ and 3′ Ends of the rep/cap Genes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conroy, G. How gene therapy is emerging from its ‘dark age’. Nature 2022, 612, S24–S26. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Liu, S.; Ou, L. rAAV immunogenicity, toxicity, and durability in 255 clinical trials: A meta-analysis. Front. Immunol. 2022, 13, 1001263. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. FDA approves first haemophilia B gene therapy. Nat. Rev. Drug Discov. 2023, 22, 7. [Google Scholar] [CrossRef]
- Penaud-Budloo, M.; Francois, A.; Clement, N.; Ayuso, E. Pharmacology of Recombinant Adeno-associated Virus Production. Mol. Ther. Methods Clin. Dev. 2018, 8, 166–180. [Google Scholar] [CrossRef]
- Wright, J.F. Product-Related Impurities in Clinical-Grade Recombinant AAV Vectors: Characterization and Risk Assessment. Biomedicines 2014, 2, 80–97. [Google Scholar] [CrossRef]
- Gombold, J.; Karakasidis, S.; Niksa, P.; Podczasy, J.; Neumann, K.; Richardson, J.; Sane, N.; Johnson-Leva, R.; Randolph, V.; Sadoff, J.; et al. Systematic evaluation of in vitro and in vivo adventitious virus assays for the detection of viral contamination of cell banks and biological products. Vaccine 2014, 32, 2916–2926. [Google Scholar] [CrossRef] [PubMed]
- Samulski, R.J.; Chang, L.S.; Shenk, T. Helper-free stocks of recombinant adeno-associated viruses: Normal integration does not require viral gene expression. J. Virol. 1989, 63, 3822–3828. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.S.; Khuntirat, B.; Qing, K.; Ponnazhagan, S.; Kube, D.M.; Zhou, S.; Dwarki, V.J.; Srivastava, A. Characterization of wild-type adeno-associated virus type 2-like particles generated during recombinant viral vector production and strategies for their elimination. J. Virol. 1998, 72, 5472–5480. [Google Scholar] [CrossRef]
- Allen, J.M.; Debelak, D.J.; Reynolds, T.C.; Miller, A.D. Identification and elimination of replication-competent adeno-associated virus (AAV) that can arise by nonhomologous recombination during AAV vector production. J. Virol. 1997, 71, 6816–6822. [Google Scholar] [CrossRef]
- Meier, A.F.; Fraefel, C.; Seyffert, M. The Interplay between Adeno-Associated Virus and its Helper Viruses. Viruses 2020, 12, 662. [Google Scholar] [CrossRef]
- Spengler, U.; Fischer, H.-P.; Caselmann, W.H. Liver Disease Associated with Viral Infections. In Zakim and Boyer’s Hepatology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 629–643. [Google Scholar] [CrossRef]
- Lalazar, G.; Ilan, Y. Viral Diseases of the Liver. In Liver Immunology: Principles and Practice; Springer: Berlin/Heidelberg, Germany, 2013; pp. 159–171. [Google Scholar] [CrossRef]
- Gao, G.; Qu, G.; Burnham, M.S.; Huang, J.; Chirmule, N.; Joshi, B.; Yu, Q.C.; Marsh, J.A.; Conceicao, C.M.; Wilson, J.M. Purification of recombinant adeno-associated virus vectors by column chromatography and its performance in vivo. Hum. Gene Ther. 2000, 11, 2079–2091. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Lerch, T.F.; Hare, J.T.; Chapman, M.S. A pseudo-plaque method for infectious particle assay and clonal isolation of adeno-associated virus. J. Virol. Methods 2010, 170, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Tai, P.W.L.; Xie, J.; Fong, K.; Seetin, M.; Heiner, C.; Su, Q.; Weiand, M.; Wilmot, D.; Zapp, M.L.; Gao, G. Adeno-associated Virus Genome Population Sequencing Achieves Full Vector Genome Resolution and Reveals Human-Vector Chimeras. Mol. Ther. Methods Clin. Dev. 2018, 9, 130–141. [Google Scholar] [CrossRef]
- Tran, N.T.; Heiner, C.; Weber, K.; Weiand, M.; Wilmot, D.; Xie, J.; Wang, D.; Brown, A.; Manokaran, S.; Su, Q.; et al. AAV-Genome Population Sequencing of Vectors Packaging CRISPR Components Reveals Design-Influenced Heterogeneity. Mol. Ther. Methods Clin. Dev. 2020, 18, 639–651. [Google Scholar] [CrossRef]
- Tran, N.T.; Lecomte, E.; Saleun, S.; Namkung, S.; Robin, C.; Weber, K.; Devine, E.; Blouin, V.; Adjali, O.; Ayuso, E.; et al. Human and insect cell-produced rAAVs show differences in genome heterogeneity. Hum. Gene Ther. 2022, 33, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, M.; Terova, O. Downstream purification of adeno-associated virus for large-scale manufacturing of gene therapies. Cell Gene Ther. Insights 2020, 6, 955–963. [Google Scholar] [CrossRef]
- Joseph, S.; Russell, D. Molecular Cloning: A Laboratory Manual. In Alkaline Agarose Gel Electrohoresis; Green, M.R., Sambrook, J., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012; Volume 1, pp. 636–666. [Google Scholar]
- Afgan, E.; Sloggett, C.; Goonasekera, N.; Makunin, I.; Benson, D.; Crowe, M.; Gladman, S.; Kowsar, Y.; Pheasant, M.; Horst, R.; et al. Genomics Virtual Laboratory: A Practical Bioinformatics Workbench for the Cloud. PLoS ONE 2015, 10, e0140829. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar] [CrossRef]
- Yan, Z.; Sun, X.; Feng, Z.; Li, G.; Fisher, J.T.; Stewart, Z.A.; Engelhardt, J.F. Optimization of Recombinant Adeno-Associated Virus-Mediated Expression for Large Transgenes, Using a Synthetic Promoter and Tandem Array Enhancers. Hum. Gene Ther. 2015, 26, 334–346. [Google Scholar] [CrossRef]
- Excoffon, K.J.; Koerber, J.T.; Dickey, D.D.; Murtha, M.; Keshavjee, S.; Kaspar, B.K.; Zabner, J.; Schaffer, D.V. Directed evolution of adeno-associated virus to an infectious respiratory virus. Proc. Natl. Acad. Sci. USA 2009, 106, 3865–3870. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Tattersall, P. The autonomously replicating parvoviruses of vertebrates. Adv. Virus Res. 1987, 33, 91–174. [Google Scholar] [PubMed]
- Nguyen, G.N.; Everett, J.K.; Kafle, S.; Roche, A.M.; Raymond, H.E.; Leiby, J.; Wood, C.; Assenmacher, C.A.; Merricks, E.P.; Long, C.T.; et al. A long-term study of AAV gene therapy in dogs with hemophilia A identifies clonal expansions of transduced liver cells. Nat. Biotechnol. 2021, 39, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Dalwadi, D.A.; Calabria, A.; Tiyaboonchai, A.; Posey, J.; Naugler, W.E.; Montini, E.; Grompe, M. AAV integration in human hepatocytes. Mol. Ther. 2021, 29, 2898–2909. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, D.E.; Bushman, F.D.; Chandler, R.J.; Crystal, R.G.; Davidson, B.L.; Dolmetsch, R.; Eggan, K.C.; Gao, G.; Gil-Farina, I.; Kay, M.A.; et al. Evaluating the state of the science for adeno-associated virus integration: An integrated perspective. Mol. Ther. 2022, 30, 2646–2663. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, P.; Yu, X.; Frabutt, D.A.; Lam, A.K.; Mulcrone, P.L.; Chrzanowski, M.; Firrman, J.; Pouchnik, D.; Sang, N.; et al. Subgenomic particles in rAAV vectors result from DNA lesion/break and non-homologous end joining of vector genomes. Mol. Ther. Nucleic Acids 2022, 29, 852–861. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Lot A | Lot B |
|---|---|---|
| Total reads | 1,839,036 | 6,499,874 |
| hg38 (human genome) | 1,415,770 (76.98%) | 5,393,393 (82.98%) |
| rep/cap plasmid | 230,787 (12.55%) | 217,501 (3.35%) |
| AAV-GOI | 103,613 (5.63%) | 31,127 (0.49%) |
| Lot A Forms | Sequence | Read Count (%) Total 3147 |
|---|---|---|
| 5′-end rep/cap |  | |
| A-5.1 |  | 3073 (97.65%) |
| A-5.2 |  | 23 (0.73%) |
| A-5.3 |  | 20 (0.64%) |
| A-5.4 |  | 11 (0.35%) |
| A-5.5 |  | 6 (0.19%) |
| A-5.6 |  | 6 (0.19%) |
| A-5.7 |  | 5 (0.16%) |
| Lot B forms | Read count (%) Total 1019 | |
| B-5.1 |  | 487 (47.79%) |
| B-5.2 |  | 395 (38.76%) |
| B-5.3 |  | 44 (4.32%) |
| B-5.4 |  | 21 (2.06%) |
| B-5.5 |  | 11 (1.08%) |
| B-5.6 |  | 9 (0.88%) |
| B-5.7 |  | 8 (0.79%) |
| B-5.8 |  | 7 (0.69%) |
| B-5.9 |  | 7 (0.69%) |
| B-5.10 |  | 3 (0.29%) |
| B-5.11 |  | 2 (0.20%) |
| 5′-ITR (green) Partial Ad ITR (blue) AAV2 p5 promoter (red) | ||
| Lot A Form | Sequence | Read Count (%) Total 3159 |
|---|---|---|
| 3′-end rep/cap |  | |
| A-3.1 |  | 3,136 (99.27%) |
| Lot B forms | Read count (%) Total 1058 | |
| B-3.1 |  | 947 (89.51%) |
| B-3.2 |  | 86 (8.13%) |
| B-3.3 |  | 8 (0.76%) |
| AAV2 3′ end (magenta) AAV2 p5 partial insert (red) 3’-ITR (green) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yip, M.; Chen, J.; Zhi, Y.; Tran, N.T.; Namkung, S.; Pastor, E.; Gao, G.; Tai, P.W.L. Querying Recombination Junctions of Replication-Competent Adeno-Associated Viruses in Gene Therapy Vector Preparations with Single Molecule, Real-Time Sequencing. Viruses 2023, 15, 1228. https://0-doi-org.brum.beds.ac.uk/10.3390/v15061228
Yip M, Chen J, Zhi Y, Tran NT, Namkung S, Pastor E, Gao G, Tai PWL. Querying Recombination Junctions of Replication-Competent Adeno-Associated Viruses in Gene Therapy Vector Preparations with Single Molecule, Real-Time Sequencing. Viruses. 2023; 15(6):1228. https://0-doi-org.brum.beds.ac.uk/10.3390/v15061228
Chicago/Turabian StyleYip, Mitchell, Jing Chen, Yan Zhi, Ngoc Tam Tran, Suk Namkung, Eric Pastor, Guangping Gao, and Phillip W. L. Tai. 2023. "Querying Recombination Junctions of Replication-Competent Adeno-Associated Viruses in Gene Therapy Vector Preparations with Single Molecule, Real-Time Sequencing" Viruses 15, no. 6: 1228. https://0-doi-org.brum.beds.ac.uk/10.3390/v15061228