
Sulfated Glycans Inhibit the Interaction of MERS-CoV Receptor Binding Domain with Heparin
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
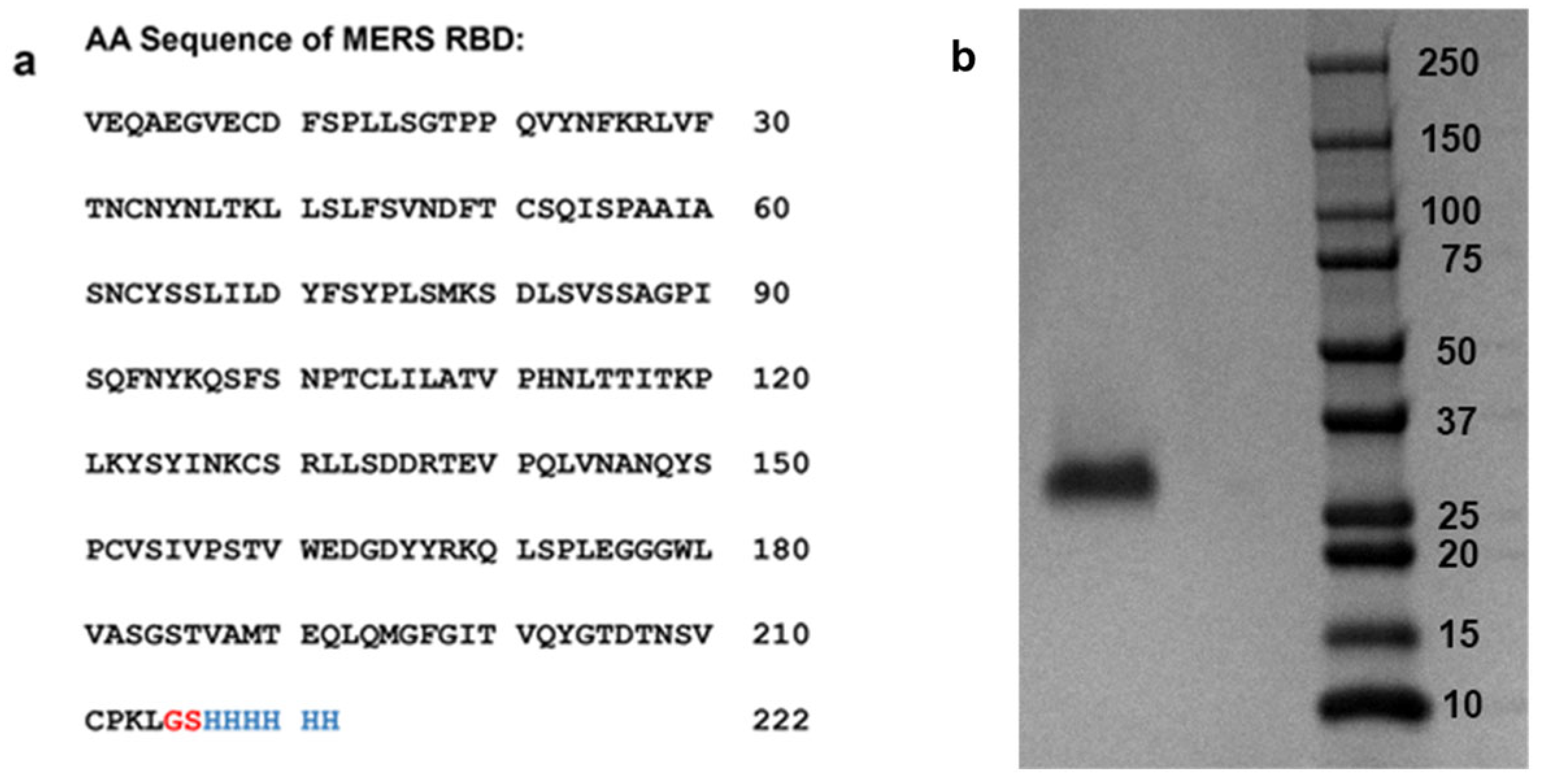
2.2. Expression and Purification of MERS-CoV RBD Protein
2.3. Preparation of Heparin Biochip
2.4. Binding Kinetics and Affinity Studies of the Interaction between Heparin and the MERS RBD Protein
2.5. Inhibition Activity of the Sulfated Glycans and Marine Sulfated Glycans on Heparin–MERS RBD Protein Interactions
2.6. Statistical Analysis
3. Results and Discussion
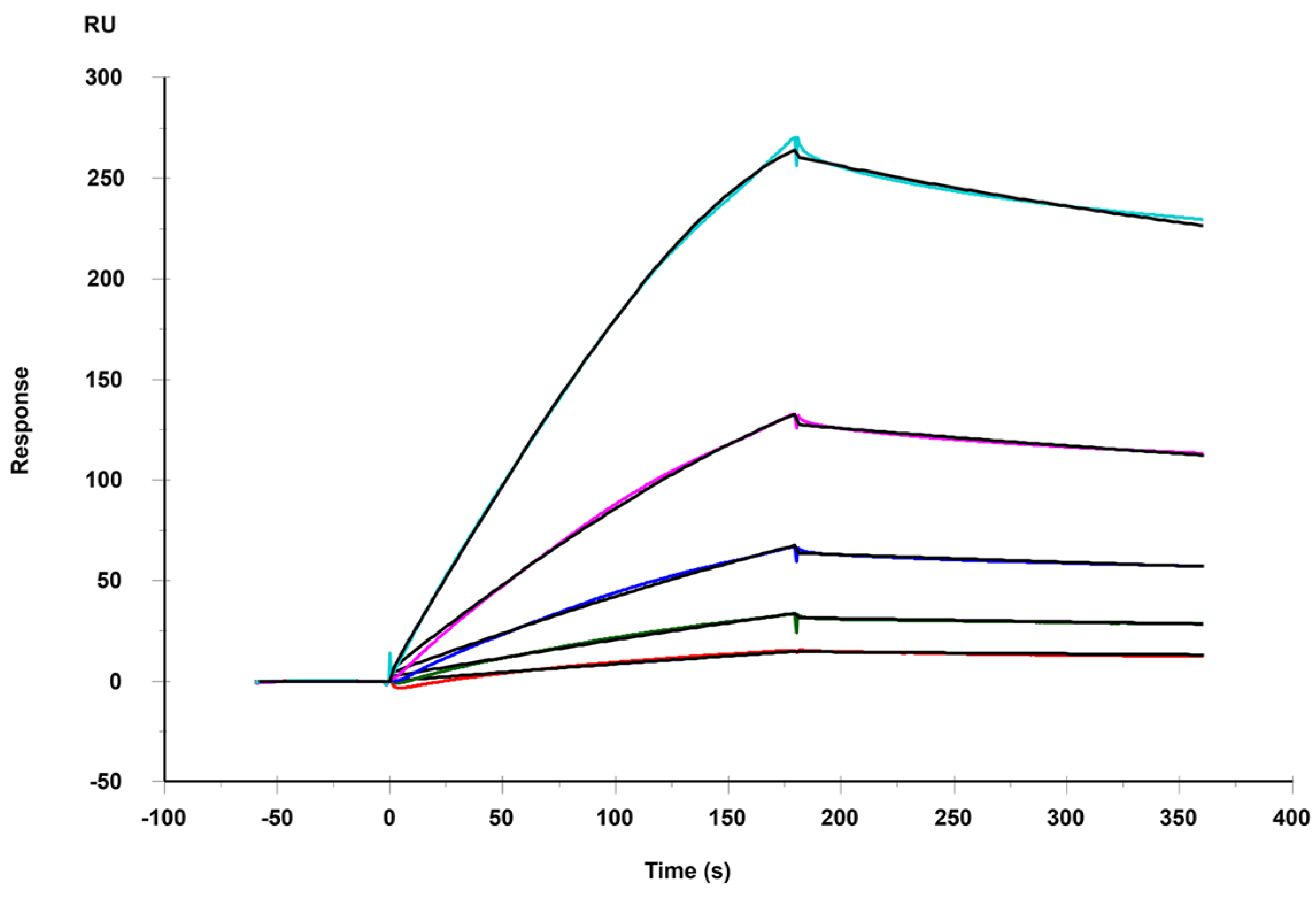
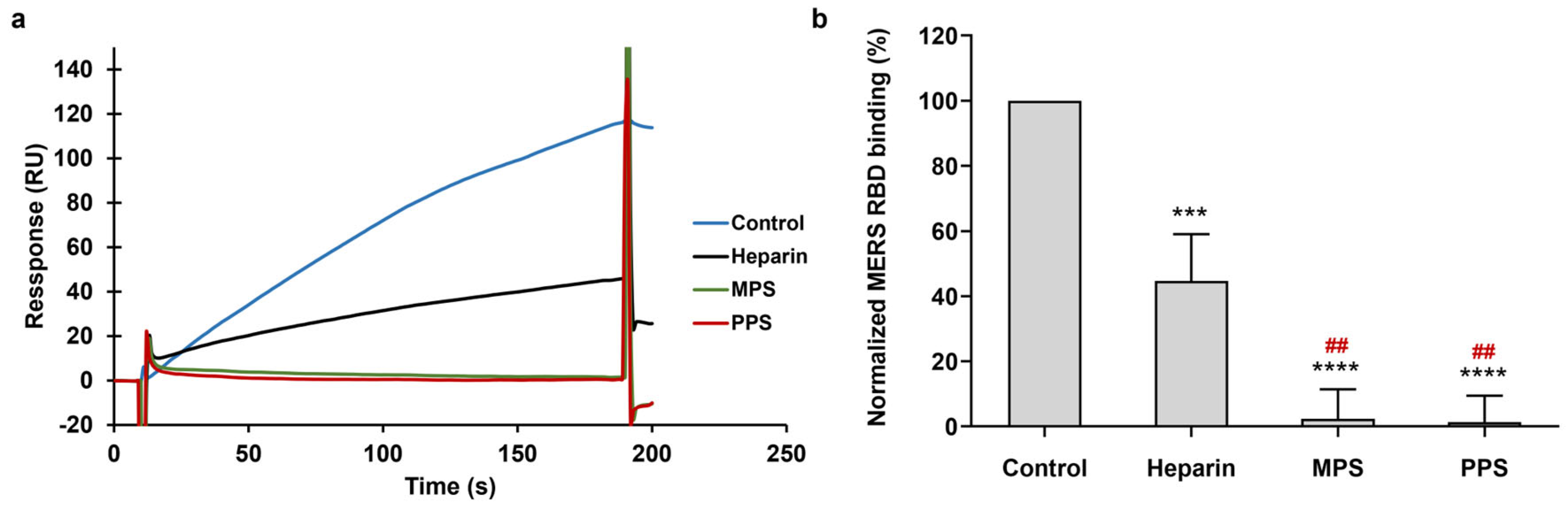
3.1. Binding Kinetics and Affinity of the Interaction between Heparin and MERS RBD Protein
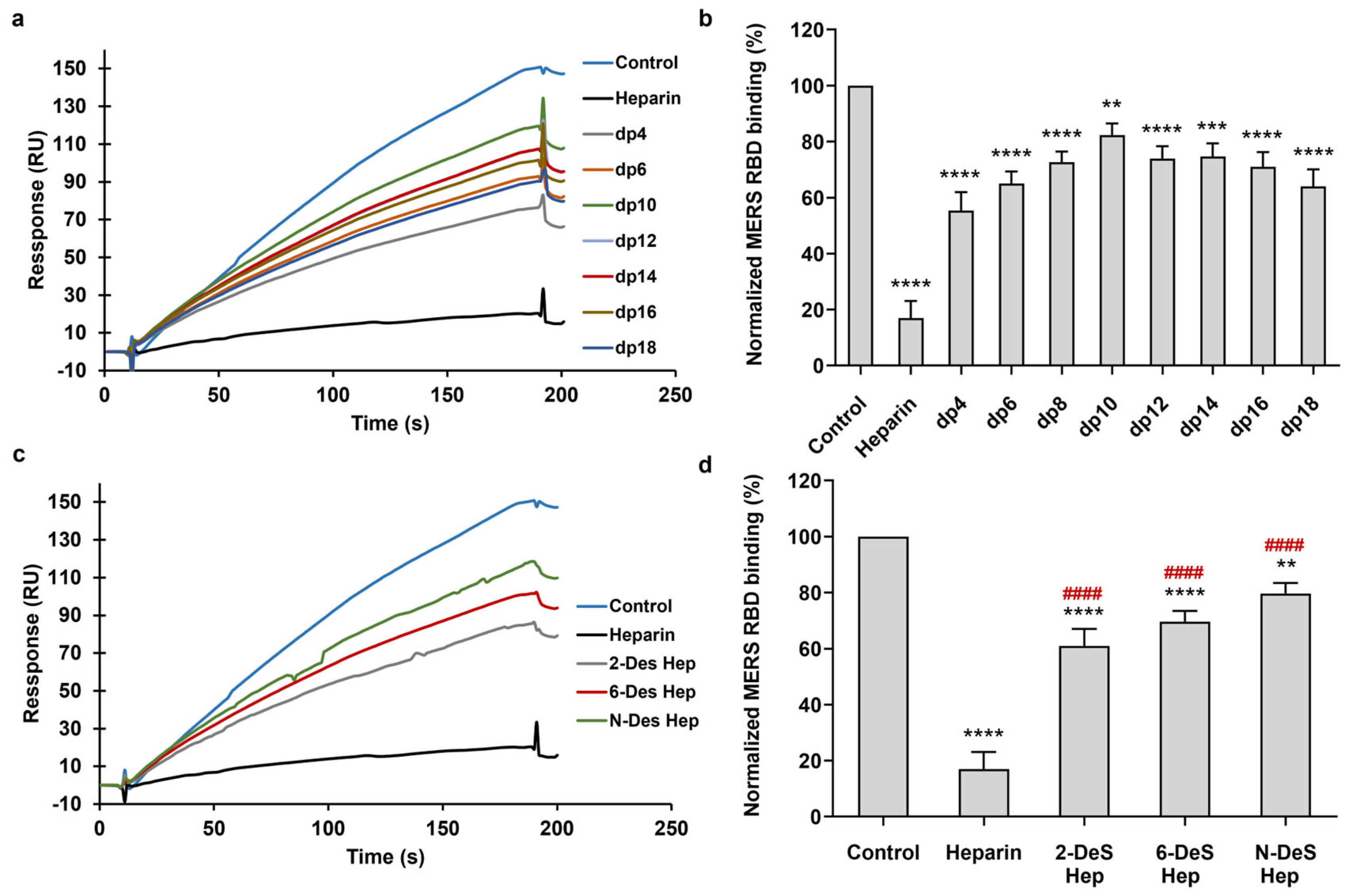
3.2. SPR Solution Competition between Surface-Immobilized Heparin and Heparin Oligosaccharides and Desulfated Heparins
3.3. Inhibition of PPS and MPS on the Interaction between Heparin and MERS RBD Protein
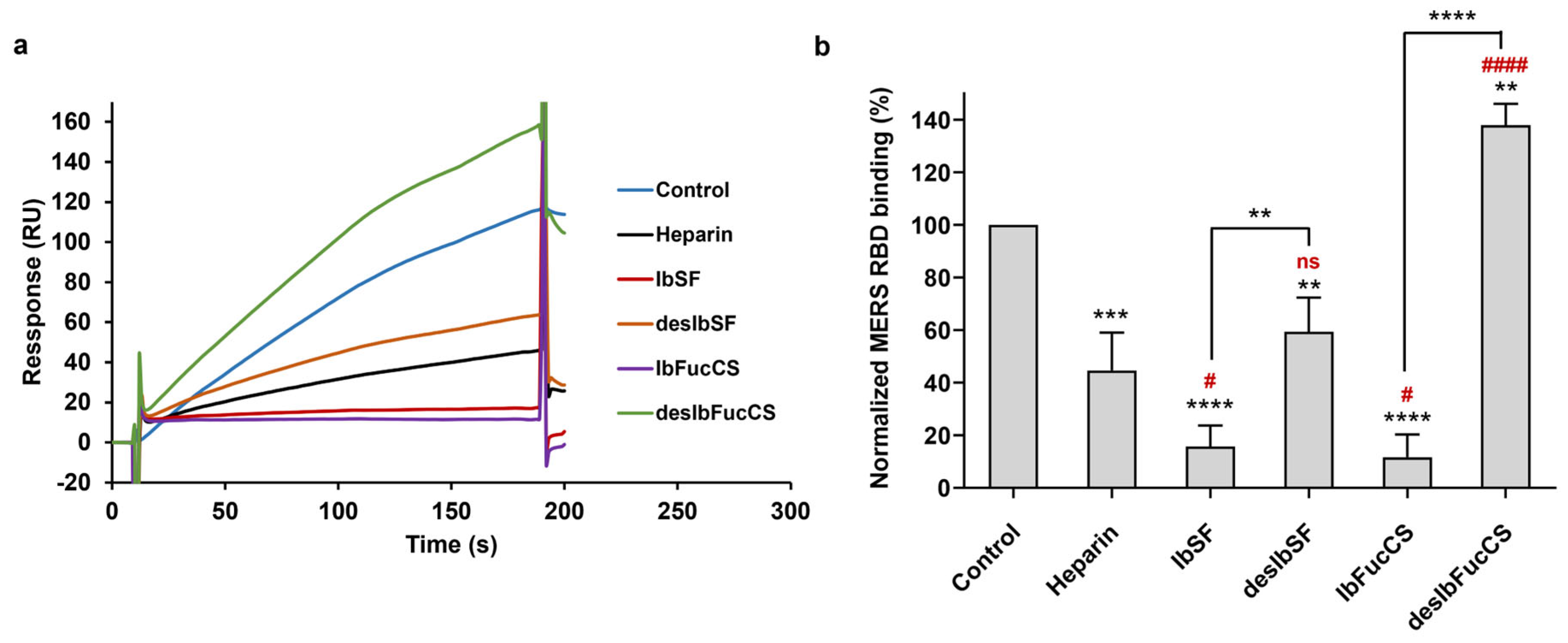
3.4. Inhibition of IbSF/IbFucCS on the Interaction between Heparin and MERS RBD Protein
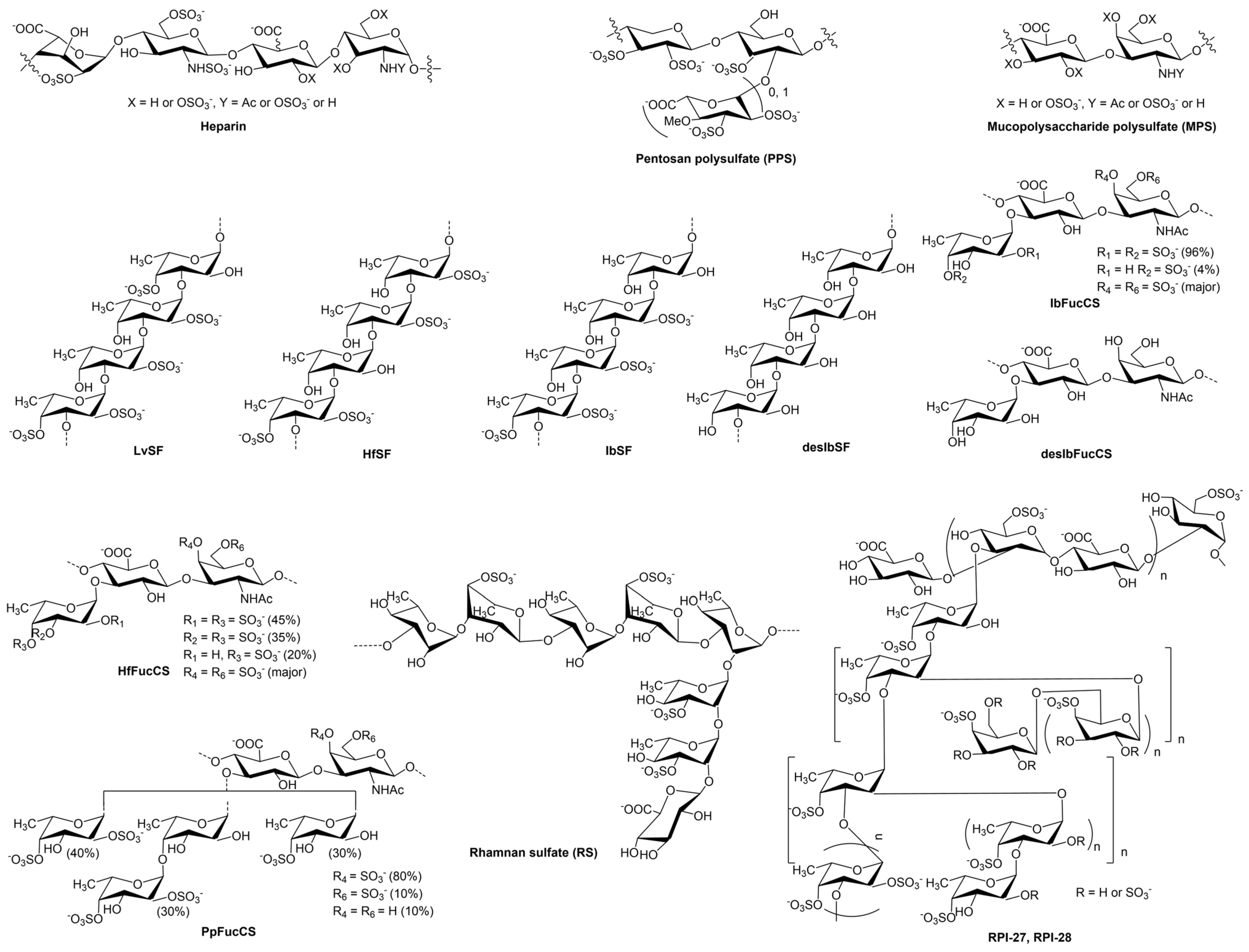
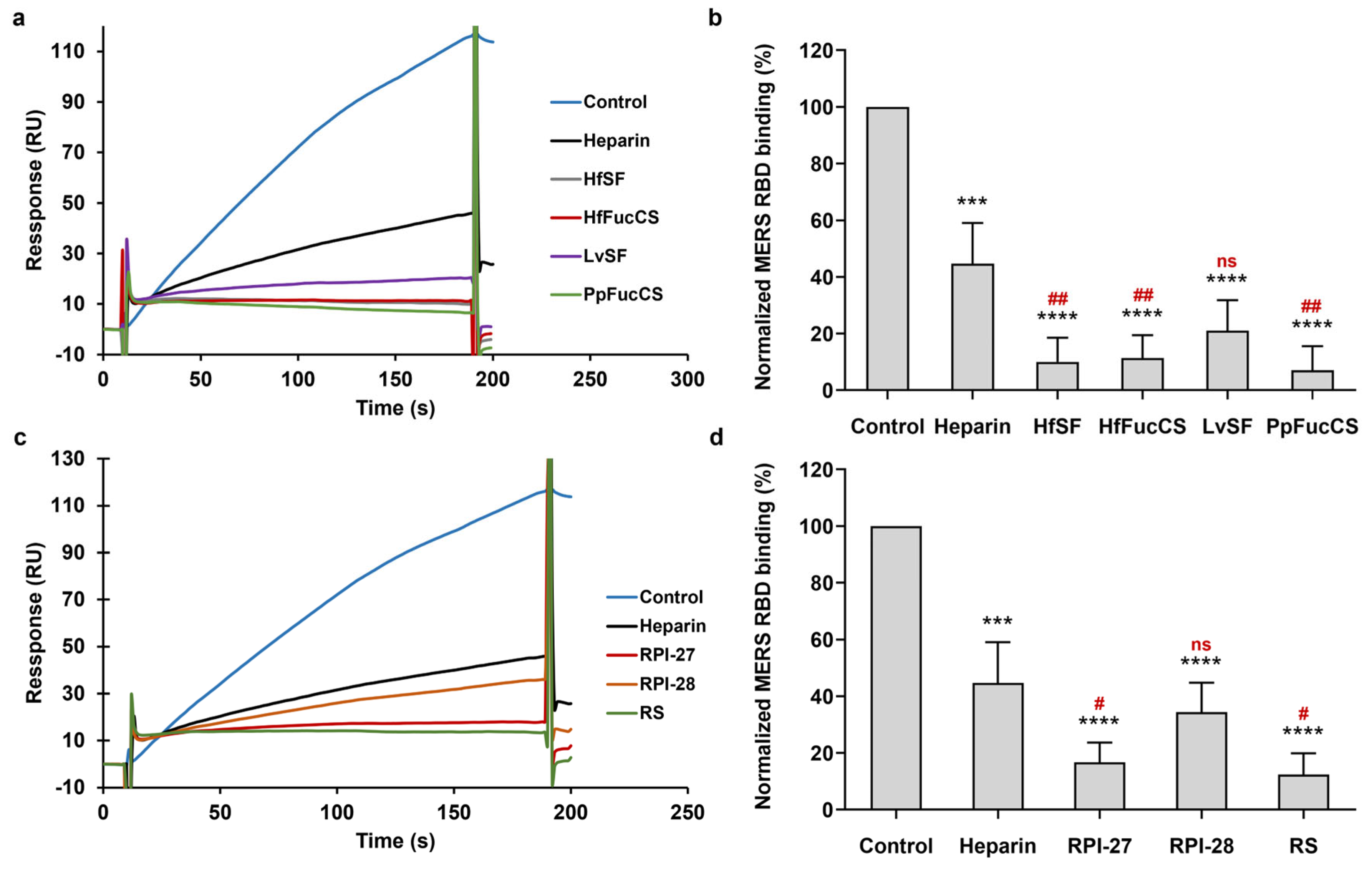
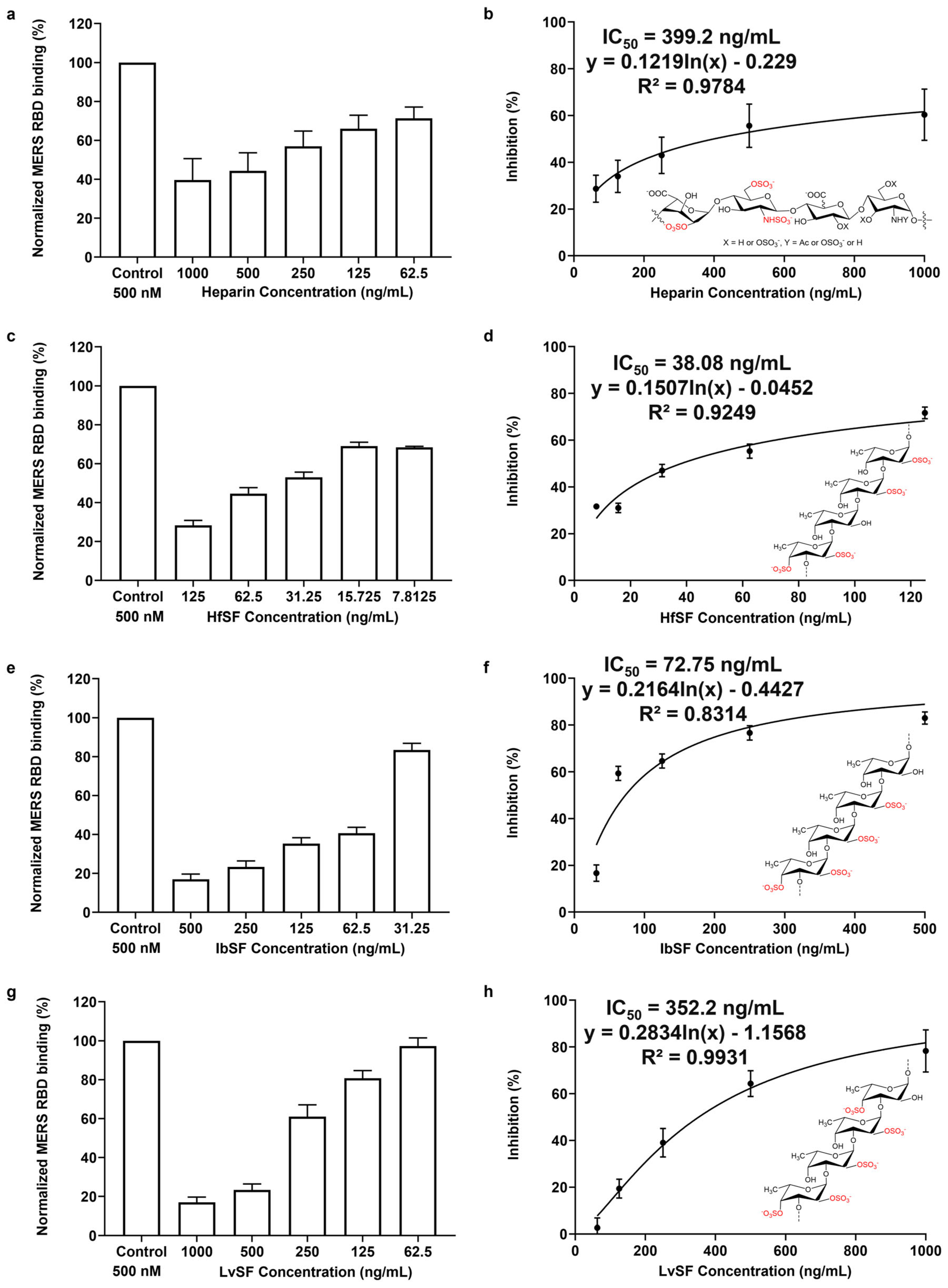
3.5. Inhibition of HfSF/HfFucCS; LvSF, PpFucCS, and Seaweed Sourced Glycans (RPI-27, RPI-28, and RS) on the Interaction between Heparin and MERS RBD Protein
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zaki, A.M.; Van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- De Groot, R.J.; Baker, S.C.; Baric, R.S.; Brown, C.S.; Drosten, C.; Enjuanes, L.; Fouchier, R.A.M.; Galiano, M.; Gorbalenya, A.E.; Memish, Z.A.; et al. Commentary: Middle East Respiratory Syndrome Coronavirus (MERS-CoV): Announcement of the Coronavirus Study Group. J. Virol. 2013, 87, 7790–7792. [Google Scholar] [CrossRef] [PubMed]
- MERS Situation Update. Available online: https://www.emro.who.int/health-topics/mers-cov/mers-outbreaks.html (accessed on 30 October 2023).
- Yang, Y.; Du, L.; Liu, C.; Wang, L.; Ma, C.; Tang, J.; Baric, R.S.; Jiang, S.; Li, F. Receptor Usage and Cell Entry of Bat Coronavirus HKU4 Provide Insight into Bat-to-Human Transmission of MERS Coronavirus. Proc. Natl. Acad. Sci. USA 2014, 111, 12516–12521. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Lu, G.; Qi, J.; Li, Y.; Wu, Y.; Deng, Y.; Geng, H.; Li, H.; Wang, Q.; Xiao, H.; et al. Structure of the Fusion Core and Inhibition of Fusion by a Heptad Repeat Peptide Derived from the S Protein of Middle East Respiratory Syndrome Coronavirus. J. Virol. 2013, 87, 13134–13140. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Filippi, G.; Cagliani, R.; De Gioia, L.; Pozzoli, U.; Al-Daghri, N.; Clerici, M.; Sironi, M. The Heptad Repeat Region Is a Major Selection Target in MERS-CoV and Related Coronaviruses. Sci. Rep. 2015, 5, 14480. [Google Scholar] [CrossRef] [PubMed]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.W.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.A.; Zaki, A.; Fouchier, R.A.M.; et al. Dipeptidyl Peptidase 4 Is a Functional Receptor for the Emerging Human Coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Millet, J.K.; Whittaker, G.R. Host Cell Entry of Middle East Respiratory Syndrome Coronavirus after Two-Step, Furin-Mediated Activation of the Spike Protein. Proc. Natl. Acad. Sci. USA 2014, 111, 15214–15219. [Google Scholar] [CrossRef]
- Xia, S.; Liu, Q.; Wang, Q.; Sun, Z.; Su, S.; Du, L.; Ying, T.; Lu, L.; Jiang, S. Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Entry Inhibitors Targeting Spike Protein. Virus Res. 2014, 194, 200–210. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Q.; Zhu, Y.; Chan, K.-H.; Qin, L.; Li, Y.; Wang, Q.; Chan, J.F.-W.; Du, L.; Yu, F.; et al. Structure-Based Discovery of Middle East Respiratory Syndrome Coronavirus Fusion Inhibitor. Nat. Commun. 2014, 5, 3067. [Google Scholar] [CrossRef]
- Xu, Y.; Lou, Z.; Liu, Y.; Pang, H.; Tien, P.; Gao, G.F.; Rao, Z. Crystal Structure of Severe Acute Respiratory Syndrome Coronavirus Spike Protein Fusion Core. J. Biol. Chem. 2004, 279, 49414–49419. [Google Scholar] [CrossRef]
- Liang, R.; Wang, L.; Zhang, N.; Deng, X.; Su, M.; Su, Y.; Hu, L.; He, C.; Ying, T.; Jiang, S.; et al. Development of Small-Molecule MERS-CoV Inhibitors. Viruses 2018, 10, 721. [Google Scholar] [CrossRef]
- Du, L.; Zhao, G.; Kou, Z.; Ma, C.; Sun, S.; Poon, V.K.M.; Lu, L.; Wang, L.; Debnath, A.K.; Zheng, B.-J.; et al. Identification of a Receptor-Binding Domain in the S Protein of the Novel Human Coronavirus Middle East Respiratory Syndrome Coronavirus as an Essential Target for Vaccine Development. J. Virol. 2013, 87, 9939–9942. [Google Scholar] [CrossRef]
- Parag, S.; Carnevale, K. COVID-19 Pathogen Viral Evolution Leading to Increased Infectivity. Cureus 2022, 14, e26660. [Google Scholar] [CrossRef] [PubMed]
- De Pasquale, V.; Quiccione, M.S.; Tafuri, S.; Avallone, L.; Pavone, L.M. Heparan Sulfate Proteoglycans in Viral Infection and Treatment: A Special Focus on SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 6574. [Google Scholar] [CrossRef]
- Ling, J.; Li, J.; Khan, A.; Lundkvist, Å.; Li, J.-P. Is Heparan Sulfate a Target for Inhibition of RNA Virus Infection? Am. J. Physiol.-Cell Physiol. 2022, 322, C605–C613. [Google Scholar] [CrossRef]
- Hao, W.; Ma, B.; Li, Z.; Wang, X.; Gao, X.; Li, Y.; Qin, B.; Shang, S.; Cui, S.; Tan, Z. Binding of the SARS-CoV-2 Spike Protein to Glycans. Sci. Bull. 2021, 66, 1205–1214. [Google Scholar] [CrossRef]
- Song, Y.; He, P.; Rodrigues, A.L.; Datta, P.; Tandon, R.; Bates, J.T.; Bierdeman, M.A.; Chen, C.; Dordick, J.; Zhang, F.; et al. Anti-SARS-CoV-2 Activity of Rhamnan Sulfate from Monostroma nitidum. Mar. Drugs 2021, 19, 685. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Shi, D.; Li, Y.; Xia, K.; Kim, S.B.; Dwivedi, R.; Farrag, M.; Pomin, V.H.; Linhardt, R.J.; Dordick, J.S.; et al. SPR Sensor-Based Analysis of the Inhibition of Marine Sulfated Glycans on Interactions between Monkeypox Virus Proteins and Glycosaminoglycans. Mar. Drugs 2023, 21, 264. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, S.; Swain, B.; Verma, R.S.; Gunthe, S.S. SARS-CoV, MERS-CoV, and 2019-nCoV Viruses: An Overview of Origin, Evolution, and Genetic Variations. VirusDisease 2020, 31, 411–423. [Google Scholar] [CrossRef]
- Oh, J.; Park, U.; Kim, J.; Jeon, K.; Kim, C.; Cho, N.-H.; Choi, Y.S. Enhancing Immune Protection against MERS-CoV: The Synergistic Effect of Proteolytic Cleavage Sites and the Fusion Peptide and RBD Domain Targeting VLP Immunization. Front. Immunol. 2023, 14, 1201136. [Google Scholar] [CrossRef]
- Shi, D.; Bu, C.; He, P.; Song, Y.; Dordick, J.S.; Linhardt, R.J.; Chi, L.; Zhang, F. Structural Characteristics of Heparin Binding to SARS-CoV-2 Spike Protein RBD of Omicron Sub-Lineages BA.2.12.1, BA.4 and BA.5. Viruses 2022, 14, 2696. [Google Scholar] [CrossRef] [PubMed]
- Bertini, S.; Alekseeva, A.; Elli, S.; Pagani, I.; Zanzoni, S.; Eisele, G.; Krishnan, R.; Maag, K.P.; Reiter, C.; Lenhart, D.; et al. Pentosan Polysulfate Inhibits Attachment and Infection by SARS-CoV-2 In Vitro: Insights into Structural Requirements for Binding. Thromb. Haemost. 2022, 122, 984–997. [Google Scholar] [CrossRef] [PubMed]
- Hoppensteadt, D.A.; Neville, B.; Schultz, C.; Jeske, W.; Raake, W.; Fareed, J. Comparative Studies on the Topical Administration of Mucopolysaccharide and Heparin Ointments in Nonhuman Primates. Clin. Appl. Thromb. Hemost. 2010, 16, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xue, C.; Yin, L.; Tang, Q.; Yu, G.; Chai, W. Comparison of Structures and Anticoagulant Activities of Fucosylated Chondroitin Sulfates from Different Sea Cucumbers. Carbohydr. Polym. 2011, 83, 688–696. [Google Scholar] [CrossRef]
- Chen, S.; Hu, Y.; Ye, X.; Li, G.; Yu, G.; Xue, C.; Chai, W. Sequence Determination and Anticoagulant and Antithrombotic Activities of a Novel Sulfated Fucan Isolated from the Sea Cucumber Isostichopus Badionotus. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 989–1000. [Google Scholar] [CrossRef] [PubMed]
- Abdulsalam, H.; Li, J.; Loka, R.S.; Sletten, E.T.; Nguyen, H.M. Heparan Sulfate-Mimicking Glycopolymers Bind SARS-CoV-2 Spike Protein in a Length- and Sulfation Pattern-Dependent Manner. ACS Med. Chem. Lett. 2023, 14, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Qi, J.; Zhang, H.; Yang, H.; Yang, Y.; Zhao, X. Comparison of Hydrothermal Depolymerization and Oligosaccharide Profile of Fucoidan and Fucosylated Chondroitin Sulfate from Holothuria Floridana. Int. J. Biol. Macromol. 2019, 132, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H.; Pereira, M.S.; Valente, A.-P.; Tollefsen, D.M.; Pavão, M.S.G.; Mourão, P.A.S. Selective Cleavage and Anticoagulant Activity of a Sulfated Fucan: Stereospecific Removal of a 2-Sulfate Ester from the Polysaccharide by Mild Acid Hydrolysis, Preparation of Oligosaccharides, and Heparin Cofactor II–Dependent Anticoagulant Activity. Glycobiology 2005, 15, 369–381. [Google Scholar] [CrossRef]
- Jin, W.; Wang, J.; Ren, S.; Song, N.; Zhang, Q. Structural Analysis of a Heteropolysaccharide from Saccharina japonica by Electrospray Mass Spectrometry in Tandem with Collision-Induced Dissociation Tandem Mass Spectrometry (ESI-CID-MS/MS). Mar. Drugs 2012, 10, 2138–2152. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Du, G.; Du, R.; Zhao, J.; Jin, Y.; Fu, S.; Gao, L.; Cheng, Z.; Lu, Q.; et al. Remdesivir in Adults with Severe COVID-19: A Randomised, Double-Blind, Placebo-Controlled, Multicentre Trial. Lancet 2020, 395, 1569–1578. [Google Scholar] [CrossRef]
- Yamashiro, Y. Anticoagulant Activity of Rhamnan Sulfate Isolated from Commercially Cultured Monostroma nitidum. Int. J. Biomed. Mater. Res. 2017, 5, 37–43. [Google Scholar] [CrossRef]
- Suzuki, K.; Terasawa, M. Biological Activities of Rhamnan Sulfate Extract from the Green Algae Monostroma nitidum (Hitoegusa). Mar. Drugs 2020, 18, 228. [Google Scholar] [CrossRef] [PubMed]
- Zoepfl, M.; Dwivedi, R.; Kim, S.B.; McVoy, M.A.; Pomin, V.H. Antiviral Activity of Marine Sulfated Glycans against Pathogenic Human Coronaviruses. Sci. Rep. 2023, 13, 4804. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ka (M−1 S−1) | kd (S−1) | KD (M) | |
|---|---|---|---|
| MERS RBD | 4.43 × 104 (±460) * | 1.45 × 10−3 (±5.8 × 10−6) * | 2.94 × 10−8 (±4.5 × 10−9) ** |
| Heparin | RPI-27 | RPI-28 | RS | HfSF | IbSF | Lvsf | |
|---|---|---|---|---|---|---|---|
| IC50 (ng/mL) | 399.2 | 21.1 | 269.4 | 17.2 | 38.1 | 72.8 | 352.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Song, Y.; Jin, W.; Xia, K.; Burnett, G.C.; Qiao, W.; Bates, J.T.; Pomin, V.H.; Wang, C.; Qiao, M.; et al. Sulfated Glycans Inhibit the Interaction of MERS-CoV Receptor Binding Domain with Heparin. Viruses 2024, 16, 237. https://0-doi-org.brum.beds.ac.uk/10.3390/v16020237
Yang J, Song Y, Jin W, Xia K, Burnett GC, Qiao W, Bates JT, Pomin VH, Wang C, Qiao M, et al. Sulfated Glycans Inhibit the Interaction of MERS-CoV Receptor Binding Domain with Heparin. Viruses. 2024; 16(2):237. https://0-doi-org.brum.beds.ac.uk/10.3390/v16020237
Chicago/Turabian StyleYang, Jiyuan, Yuefan Song, Weihua Jin, Ke Xia, Grace C. Burnett, Wanjin Qiao, John T. Bates, Vitor H. Pomin, Chunyu Wang, Mingqiang Qiao, and et al. 2024. "Sulfated Glycans Inhibit the Interaction of MERS-CoV Receptor Binding Domain with Heparin" Viruses 16, no. 2: 237. https://0-doi-org.brum.beds.ac.uk/10.3390/v16020237